Вам также может понравиться
- Appareil génital: Les Grands Articles d'UniversalisОт EverandAppareil génital: Les Grands Articles d'UniversalisОценок пока нет
- HTTPS://FR - Scribd.com/doc/86326849/tableau Myologie Du Membre InfДокумент21 страницаHTTPS://FR - Scribd.com/doc/86326849/tableau Myologie Du Membre Infsorades68Оценок пока нет
- EMBRYOLOGIE NOTES Final PDFДокумент32 страницыEMBRYOLOGIE NOTES Final PDFmadauleОценок пока нет
- Sexualisation en biologie: Les Grands Articles d'UniversalisОт EverandSexualisation en biologie: Les Grands Articles d'UniversalisОценок пока нет
- Meiose Et GametogeneseДокумент4 страницыMeiose Et GametogeneseFaculté De Médecine Béchar100% (1)
- 02 - Gamétogenèse GДокумент50 страниц02 - Gamétogenèse GIslęm OuaribОценок пока нет
- Résumé - GamétogénèseДокумент3 страницыRésumé - GamétogénèseRafaОценок пока нет
- Le Cycle CellulaireДокумент16 страницLe Cycle CellulaireyoussefОценок пока нет
- LA ReproductionДокумент10 страницLA ReproductionRim ElmoutaoukkilОценок пока нет
- PR Yardin - GamétogenèseДокумент21 страницаPR Yardin - GamétogenèsehauswirthОценок пока нет
- COURS D'embryologie-ConvertiДокумент24 страницыCOURS D'embryologie-ConvertiAnas IsmailОценок пока нет
- 2Документ62 страницы2Sofiane BousifeОценок пока нет
- Chapitre 03 Le Cycle CellulaireДокумент10 страницChapitre 03 Le Cycle CellulaireĐî ŅæОценок пока нет
- Détermination Du SexeДокумент9 страницDétermination Du SexeTaniaОценок пока нет
- TD ThallophytesДокумент5 страницTD ThallophytesKaoutar ChaoukiОценок пока нет
- UNITÉ 2 Corrigé PDFДокумент10 страницUNITÉ 2 Corrigé PDFIrene PGОценок пока нет
- Biologie Animale GametogeneseДокумент23 страницыBiologie Animale GametogeneseRodo KoulОценок пока нет
- Chapitre I Les Premieres Cellules Du DeveloppementДокумент13 страницChapitre I Les Premieres Cellules Du DeveloppementŽdn ÂśšiaОценок пока нет
- Chapitre Ii Cycles de VieДокумент5 страницChapitre Ii Cycles de Vieevident 2014Оценок пока нет
- Chapitre II SpermatogenèseДокумент6 страницChapitre II SpermatogenèseDel HadОценок пока нет
- TD 4 Physiologie de La ReproductionДокумент5 страницTD 4 Physiologie de La ReproductionMorgane FaconОценок пока нет
- Embryologie A1Chapitre1Gamétogénèse PDFДокумент24 страницыEmbryologie A1Chapitre1Gamétogénèse PDFVéto SamОценок пока нет
- UE2 Embryologie - Gamétogenèse (Final)Документ18 страницUE2 Embryologie - Gamétogenèse (Final)sun-nee-chan9Оценок пока нет
- Développement Embryonaire HumaineДокумент11 страницDéveloppement Embryonaire HumaineWijdene SassiОценок пока нет
- Gamatogenese Et FecondationДокумент47 страницGamatogenese Et FecondationAmïn Fïlalï100% (1)
- Cours de Biologie Générale Chapitres 3 À 5 Pour 2022Документ93 страницыCours de Biologie Générale Chapitres 3 À 5 Pour 2022jeffkakwanda10Оценок пока нет
- GENETIQUE Chapitre 2Документ114 страницGENETIQUE Chapitre 2LectorОценок пока нет
- La Division Cellulaire Mitose Et Méiose: Institut National de Formation Paramédicale BatnaДокумент11 страницLa Division Cellulaire Mitose Et Méiose: Institut National de Formation Paramédicale BatnaDelenda ChamchoumaОценок пока нет
- CytogénétiqueДокумент33 страницыCytogénétiqueFaiza HОценок пока нет
- Embryologie Médicale Par Roumaissa HRCДокумент57 страницEmbryologie Médicale Par Roumaissa HRCsevda mouvieОценок пока нет
- MI5 Genetique Reproduction Developpement Histologie de L Appareil Genital Pellestor2Документ10 страницMI5 Genetique Reproduction Developpement Histologie de L Appareil Genital Pellestor2Djallal HassaniОценок пока нет
- Fiche 3 TD 1Документ16 страницFiche 3 TD 1SalifОценок пока нет
- Biologie Complet S2Документ104 страницыBiologie Complet S2K Walid100% (1)
- SpermatozoïdeДокумент23 страницыSpermatozoïdeElja MohcineОценок пока нет
- Chap 5 FécondationДокумент11 страницChap 5 Fécondationabdoul djalil soumanaОценок пока нет
- Partie 1 Chap 1 Spé SVT Division CellulaireДокумент8 страницPartie 1 Chap 1 Spé SVT Division CellulaireNagwa Elimam100% (2)
- Histologie - Tubes Séminifère Et SpermatogéneseДокумент7 страницHistologie - Tubes Séminifère Et Spermatogéneseaacer3976Оценок пока нет
- Cycle CellulaireДокумент37 страницCycle CellulairegigiОценок пока нет
- Cours Etudiants V2 2016Документ44 страницыCours Etudiants V2 2016Zaidou RoambaОценок пока нет
- Revisions Du Cours SVI M2 S1 Avec QCEMДокумент52 страницыRevisions Du Cours SVI M2 S1 Avec QCEMazzamОценок пока нет
- Embryologie 1 - Les GametesДокумент5 страницEmbryologie 1 - Les GametesAmirou Baby MixicoОценок пока нет
- La GamétogenèseДокумент56 страницLa GamétogenèseFORN01 Dams01Оценок пока нет
- Développement Embryonnaire de La DrosophileДокумент25 страницDéveloppement Embryonnaire de La DrosophileSarah OlivierОценок пока нет
- Division Cellulaire 1Документ7 страницDivision Cellulaire 1islam04.maОценок пока нет
- 4 5 Exp Eco VegДокумент72 страницы4 5 Exp Eco VegMohammed EcharradiОценок пока нет
- Chapitre 6 La MitoseДокумент5 страницChapitre 6 La MitoseMeghane TagliaroОценок пока нет
- Cours ReproductionДокумент7 страницCours ReproductionBen MabОценок пока нет
- Chap 1 - Doc 4Документ21 страницаChap 1 - Doc 4LB3ALBAKIОценок пока нет
- Mon Cours - OdtДокумент4 страницыMon Cours - OdtAlea JajomaОценок пока нет
- Cours Sur La MeioseДокумент4 страницыCours Sur La MeioseYassine RajiОценок пока нет
- Cycle CellulaireДокумент52 страницыCycle CellulaireosefresistanceОценок пока нет
- Cours Etudiants pdf160215 PDFДокумент38 страницCours Etudiants pdf160215 PDFHichem HammadiОценок пока нет
- Chapitre 14Документ28 страницChapitre 14TML7Оценок пока нет
- Cours 3 Le Cycle Cellulaire PDFДокумент17 страницCours 3 Le Cycle Cellulaire PDFmalak kerakbiОценок пока нет
- Biologie Cellulaire IIIДокумент12 страницBiologie Cellulaire IIIInas HassanatОценок пока нет
- Cours Entier BDRДокумент45 страницCours Entier BDRrobin_faure5146Оценок пока нет
- Physiologie Sexuelle MasculinefДокумент30 страницPhysiologie Sexuelle Masculinefsteel gamОценок пока нет
- Chap2 - Cours Physiologie Cellulaire Ufr Stapv - Ussein 2Документ60 страницChap2 - Cours Physiologie Cellulaire Ufr Stapv - Ussein 2sowcapo2004Оценок пока нет
- MitoseДокумент10 страницMitoseOumarou KontaОценок пока нет
- Bactériologie (Version 1)Документ32 страницыBactériologie (Version 1)coursenligne.lille183% (6)
- Trafic Intracellulaire Et Compartiment CellulaireДокумент24 страницыTrafic Intracellulaire Et Compartiment Cellulairecoursenligne.lille186% (7)
- Eucaryotes Et Virologie (Version 1)Документ46 страницEucaryotes Et Virologie (Version 1)coursenligne.lille188% (17)
- Eucaryotes Et Virologie (Version 2)Документ24 страницыEucaryotes Et Virologie (Version 2)coursenligne.lille1100% (3)
- Bactériologie (Version 2)Документ70 страницBactériologie (Version 2)coursenligne.lille1100% (4)
- ExocytoseДокумент18 страницExocytosecoursenligne.lille1100% (1)
- Hérédité Cytoplasmique PDFДокумент22 страницыHérédité Cytoplasmique PDFcoursenligne.lille1100% (4)
- TD Mutants ThermosensiblesДокумент5 страницTD Mutants Thermosensiblescoursenligne.lille1100% (1)
- Optique GéométriqueДокумент51 страницаOptique Géométriquecoursenligne.lille1100% (1)
- TD1 Croissance BactérienneДокумент9 страницTD1 Croissance Bactériennecoursenligne.lille1100% (9)
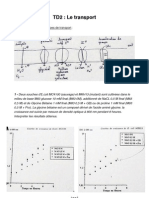
- TD Le TransportДокумент6 страницTD Le Transportcoursenligne.lille1Оценок пока нет
- Le Système Nerveux VégétatifДокумент15 страницLe Système Nerveux Végétatifcoursenligne.lille1Оценок пока нет
- Mécanique Des FluidesДокумент42 страницыMécanique Des Fluidescoursenligne.lille1100% (14)
- Bactériologie FondamentaleДокумент47 страницBactériologie Fondamentalecoursenligne.lille1100% (6)
- TD Milieu de CultureДокумент8 страницTD Milieu de Culturecoursenligne.lille180% (10)
- Les ChampignonsДокумент8 страницLes Champignonscoursenligne.lille1100% (6)
- AlgologieДокумент17 страницAlgologiecoursenligne.lille194% (17)
- Transgenèse Végétale Et ApplicationsДокумент16 страницTransgenèse Végétale Et Applicationscoursenligne.lille1100% (13)
- Les GlucidesДокумент21 страницаLes Glucidescoursenligne.lille1100% (3)
- Les ProtidesДокумент17 страницLes Protidescoursenligne.lille1100% (1)
- Biotechnologies VégétalesДокумент27 страницBiotechnologies Végétalescoursenligne.lille190% (30)
- Physiologie RespiratoireДокумент35 страницPhysiologie Respiratoirecoursenligne.lille1100% (16)
- TD Les Applications de La Fusion de ProtoplasteДокумент12 страницTD Les Applications de La Fusion de Protoplastecoursenligne.lille1100% (6)
- Fécondation Et GamétogènèseДокумент17 страницFécondation Et Gamétogènèsecoursenligne.lille1Оценок пока нет
- L'excrétion RénaleДокумент27 страницL'excrétion Rénalecoursenligne.lille1100% (1)
- TD Développement Des Ailes de DrosophileДокумент3 страницыTD Développement Des Ailes de Drosophilecoursenligne.lille1100% (1)
- TD DiapauseДокумент9 страницTD Diapausecoursenligne.lille1100% (2)
- Métamorphose Des AmphibiensДокумент13 страницMétamorphose Des Amphibienscoursenligne.lille1100% (1)
- Mue Et Métamorphose Des InsectesДокумент16 страницMue Et Métamorphose Des Insectescoursenligne.lille1100% (7)
- La ReproductionДокумент22 страницыLa Reproductionbiguilou100% (1)
- Chapitre VReproduction 114Документ83 страницыChapitre VReproduction 114mouhamadoulaminembengue64Оценок пока нет
- Mini Manuel Biologie VegetaleДокумент19 страницMini Manuel Biologie VegetalepletoyerОценок пока нет
- 6-La FécondationДокумент2 страницы6-La FécondationahlemmОценок пока нет
- Unite II - Chapitre1 - Reproduction Sexuee Chez Les Spermaphytes Lydex2022 - 2023 Copie Prof 1Документ27 страницUnite II - Chapitre1 - Reproduction Sexuee Chez Les Spermaphytes Lydex2022 - 2023 Copie Prof 1mohamedlaayoune252Оценок пока нет
- Cours 1 - Introduction À La Biologie AnimaleДокумент11 страницCours 1 - Introduction À La Biologie AnimaleNãřímäñ KŁayОценок пока нет
- Corrige Type SVT TD Dre-M 1er Sem 22-23Документ1 страницаCorrige Type SVT TD Dre-M 1er Sem 22-23Gnandi BilantéОценок пока нет
- Embryologie S1 - 2020-2021 PolyДокумент29 страницEmbryologie S1 - 2020-2021 PolyDec30mdi BtbОценок пока нет
- Systématique Évolutive - ReimersДокумент139 страницSystématique Évolutive - Reimersanonymedz100% (1)
- Diapo - Gamétog & Fécond DéfinitifДокумент14 страницDiapo - Gamétog & Fécond DéfinitifL kОценок пока нет
- Fee 406Документ2 страницыFee 406Taaleem SchoolОценок пока нет
- Cours Botanique 2020 Partie 2 Bryophytes Et Ptéridophytes PDFДокумент13 страницCours Botanique 2020 Partie 2 Bryophytes Et Ptéridophytes PDFÑõü Hâ100% (5)
- Unmode Figuration Des Cycles Biologiques Vegetaux Applique A Ginkyo, Araucaria, Taxus, Cephalotaxus e T EphedraДокумент4 страницыUnmode Figuration Des Cycles Biologiques Vegetaux Applique A Ginkyo, Araucaria, Taxus, Cephalotaxus e T EphedraferdinandmadjadoumngueneldibayОценок пока нет
- Infertilité 2023Документ436 страницInfertilité 2023Eliezer YoungОценок пока нет
- Exercices Corrigés de Di HybridismeДокумент4 страницыExercices Corrigés de Di HybridismeAdama Bagayoko100% (1)
- Base de ComparaisonДокумент2 страницыBase de Comparaisonarij alouiОценок пока нет
- Monde Vivant Chap 2 EumycètesДокумент2 страницыMonde Vivant Chap 2 EumycètesGrace MM.Оценок пока нет
- Museum - MNHN Fich Ponte CorauxДокумент24 страницыMuseum - MNHN Fich Ponte CorauxcyrilОценок пока нет
- H - FécondationДокумент25 страницH - FécondationFotso Nanfa0% (1)
- Print 1669642857495Документ1 страницаPrint 1669642857495amjad elbОценок пока нет
- Chapitre 2 Les Lois StatistiquesДокумент47 страницChapitre 2 Les Lois StatistiquesmouadboulhrouziОценок пока нет
- Controle 2 s2 2apic-1Документ1 страницаControle 2 s2 2apic-1Mohammed BrakineОценок пока нет
- CP 1 Dihybdrdisme 2324Документ3 страницыCP 1 Dihybdrdisme 2324Lena FettahiОценок пока нет
- Série Des Exos SpermaphyteДокумент3 страницыSérie Des Exos Spermaphytehassen22089782Оценок пока нет
- Fécondation Fusion Des NoyauxДокумент3 страницыFécondation Fusion Des NoyauxvivianogabrielpОценок пока нет
- Cycles de Développement Des ThallophytesДокумент6 страницCycles de Développement Des ThallophytesLSTL TACQОценок пока нет
- 7L2 Formation Du FruitДокумент12 страниц7L2 Formation Du FruitMelchi YadongaОценок пока нет
- Chapitre 3 +Документ13 страницChapitre 3 +محكمة TAKI DZОценок пока нет
- Chapitre 9 ST CorrДокумент2 страницыChapitre 9 ST Corr6233814Оценок пока нет
- Chap HaplodisationДокумент7 страницChap HaplodisationFatima Ezzahra LaftouhiОценок пока нет