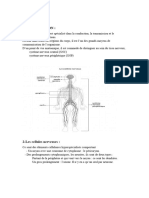
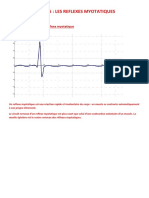
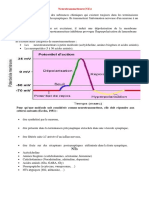
Вам также может понравиться
- Physiologie LA JONCTION NEUROMUSCULAIREДокумент9 страницPhysiologie LA JONCTION NEUROMUSCULAIREtoni.slmОценок пока нет
- Chapitre 4 - Les Messagers Au Niveau NeuronalДокумент68 страницChapitre 4 - Les Messagers Au Niveau NeuronalHadjer BoukhatmiОценок пока нет
- 6 - Transmission SynaptiqueДокумент7 страниц6 - Transmission Synaptiquemintchimechak2Оценок пока нет
- EchangesДокумент12 страницEchangesAmina assala Hadj MohamedОценок пока нет
- Synapse Et Transmission SynaptiqueДокумент10 страницSynapse Et Transmission SynaptiqueFaculté De Médecine BécharОценок пока нет
- CSC 1Документ4 страницыCSC 1muratОценок пока нет
- 14 Histophysiologie Du Tissu NerveuxДокумент7 страниц14 Histophysiologie Du Tissu NerveuxGeorge BorsОценок пока нет
- La Transmission SynaptiqueДокумент39 страницLa Transmission SynaptiquesmalldoctorivansОценок пока нет
- MOSTA Physiologie de La Jonction NeuromusculaireДокумент27 страницMOSTA Physiologie de La Jonction NeuromusculaireSamia BelabbasОценок пока нет
- Chapitre 3 Le Système NerveuxДокумент26 страницChapitre 3 Le Système NerveuxdfhohfeicxhОценок пока нет
- UE 2-1 Cellules Excitables Nerveuses Et Musculaires Partie IДокумент40 страницUE 2-1 Cellules Excitables Nerveuses Et Musculaires Partie IMoh LyaliОценок пока нет
- Mécanisme de La Transmission SynaptiqueДокумент11 страницMécanisme de La Transmission SynaptiqueAbubakr SalhiОценок пока нет
- 4-La Transmission Synaptique PDFДокумент14 страниц4-La Transmission Synaptique PDFNara ĪntiSsãrОценок пока нет
- Corrigé Muscles Métropole Sujet 1 2021Документ8 страницCorrigé Muscles Métropole Sujet 1 2021bellayerdianeОценок пока нет
- SN Chap3 Transmission SynaptiqueДокумент7 страницSN Chap3 Transmission SynaptiqueKevin HeimanuОценок пока нет
- Résumé QE Physio-1Документ20 страницRésumé QE Physio-1Elidlbi IbrahimОценок пока нет
- 08-Physiologie de La Jonction NeuromusculaireДокумент26 страниц08-Physiologie de La Jonction NeuromusculaireBook HunterОценок пока нет
- Révision Bac 2022 Sujet 1 CorrigéДокумент4 страницыRévision Bac 2022 Sujet 1 CorrigéClément CourbonОценок пока нет
- 4Документ1 страница4Erwann Ordinateur PhilippeОценок пока нет
- Dia sn6 SynapsesДокумент40 страницDia sn6 Synapsesكوردورلي الحاج100% (2)
- 1.physiologie Cardiaque Tanger Cours PR El Hangouche Part 1Документ104 страницы1.physiologie Cardiaque Tanger Cours PR El Hangouche Part 1elbakkalilibrairieОценок пока нет
- Tpsynpase CorrigeДокумент2 страницыTpsynpase CorrigeMankeur GueyeОценок пока нет
- Af Neuro PDFДокумент18 страницAf Neuro PDFMarco RodriguezОценок пока нет
- Physique Et TéléphoniqueДокумент4 страницыPhysique Et TéléphoniquemeyemobiangjОценок пока нет
- Resume de NeurophysiologieДокумент5 страницResume de NeurophysiologieMoussa DIAFOОценок пока нет
- Série Neurophysiologie CorrectionsДокумент14 страницSérie Neurophysiologie CorrectionsDavid King100% (2)
- C6.La Transmission SynaptiqueДокумент19 страницC6.La Transmission SynaptiqueAbdelOuahidSenhadjiОценок пока нет
- Communication NerveuseДокумент5 страницCommunication NerveusemerzougОценок пока нет
- La Cellule Nerveuse W.PDF KKДокумент6 страницLa Cellule Nerveuse W.PDF KKas.cherfaouiОценок пока нет
- Les Neurotransmetteurs PacesДокумент30 страницLes Neurotransmetteurs PacesValentin GeorgetОценок пока нет
- Exo Propriete Tissu Nerveux 7dДокумент25 страницExo Propriete Tissu Nerveux 7dNiangОценок пока нет
- La Transmission Synaptique 1Документ6 страницLa Transmission Synaptique 1safemindОценок пока нет
- Communication Cellulaire Transmission Nerveuse PDFДокумент19 страницCommunication Cellulaire Transmission Nerveuse PDFmuratОценок пока нет
- Les ReflexesДокумент2 страницыLes ReflexeslozayОценок пока нет
- Physio1an31-7jonction NeuromusculaireДокумент15 страницPhysio1an31-7jonction Neuromusculaireelidrissimaryam733Оценок пока нет
- Neuroanatomie fonctionnelle cours 2Документ43 страницыNeuroanatomie fonctionnelle cours 2ezitounifatimaezzahraОценок пока нет
- CM 8 Remodelage MVДокумент5 страницCM 8 Remodelage MVouariah.aboubakrОценок пока нет
- Chapitre 2-système nerveux-saussez- 19 03 2021_706b84660320fb875ec3ce27d2e5609cДокумент81 страницаChapitre 2-système nerveux-saussez- 19 03 2021_706b84660320fb875ec3ce27d2e5609catoiniyassineОценок пока нет
- Résumé PsychophysioДокумент34 страницыRésumé PsychophysioJoël OdiОценок пока нет
- Question Type Examens Neurophysiologie PDFДокумент10 страницQuestion Type Examens Neurophysiologie PDFStephaneОценок пока нет
- Le Cerveau HumainДокумент65 страницLe Cerveau HumainhatimmarsouliОценок пока нет
- 08.10.2015 Organisation Du Trafic IntracellulaireДокумент17 страниц08.10.2015 Organisation Du Trafic IntracellulaireEaОценок пока нет
- Notion de SynapseДокумент9 страницNotion de SynapseYumiОценок пока нет
- Le Reflexe Myotatique Resume de Cours 1Документ5 страницLe Reflexe Myotatique Resume de Cours 1Ali SafiaОценок пока нет
- SEQ 4-COURS Redige-Comportement-ReflexeДокумент6 страницSEQ 4-COURS Redige-Comportement-ReflexeLadetОценок пока нет
- Electrophysiologie CardiaqueT PDFДокумент3 страницыElectrophysiologie CardiaqueT PDFnadaОценок пока нет
- RegulationДокумент30 страницRegulationYannick DsprbsОценок пока нет
- Transmission Synaptique PDFДокумент8 страницTransmission Synaptique PDFJean PatateОценок пока нет
- 19a Transmis SynaptДокумент15 страниц19a Transmis SynaptTalel TouaitiОценок пока нет
- s-svt-obligatoire-2013-metropole-sujet-officielДокумент9 страницs-svt-obligatoire-2013-metropole-sujet-officieldememamebousso09Оценок пока нет
- Physiologie Cours COMPLETДокумент492 страницыPhysiologie Cours COMPLETProjojo99Оценок пока нет
- Trafic Intracellulaire ModifiéeДокумент33 страницыTrafic Intracellulaire ModifiéeMzn JannahОценок пока нет
- Neuro 2Документ12 страницNeuro 2tinaОценок пока нет
- CorrigeДокумент2 страницыCorrigethiescoyabaОценок пока нет
- SVT 2nd A - L3 - La Transmission Du Message NerveuxДокумент14 страницSVT 2nd A - L3 - La Transmission Du Message NerveuxMayssaeОценок пока нет
- Chapitre 1Документ16 страницChapitre 1David FMОценок пока нет
- SVT Obli G1Документ8 страницSVT Obli G1LETUDIANTОценок пока нет
- Réponses Exams BlancДокумент5 страницRéponses Exams BlancMourad Oujda100% (1)
- TD L2 - Etu - QCMДокумент7 страницTD L2 - Etu - QCMTienéОценок пока нет
- Eucaryotes Et Virologie (Version 1)Документ46 страницEucaryotes Et Virologie (Version 1)coursenligne.lille188% (17)
- Trafic Intracellulaire Et Compartiment CellulaireДокумент24 страницыTrafic Intracellulaire Et Compartiment Cellulairecoursenligne.lille186% (7)
- Eucaryotes Et Virologie (Version 2)Документ24 страницыEucaryotes Et Virologie (Version 2)coursenligne.lille1100% (3)
- TD Mutants ThermosensiblesДокумент5 страницTD Mutants Thermosensiblescoursenligne.lille1100% (1)
- Bactériologie (Version 1)Документ32 страницыBactériologie (Version 1)coursenligne.lille183% (6)
- Bactériologie FondamentaleДокумент47 страницBactériologie Fondamentalecoursenligne.lille1100% (6)
- Mécanique Des FluidesДокумент42 страницыMécanique Des Fluidescoursenligne.lille1100% (14)
- Bactériologie (Version 2)Документ70 страницBactériologie (Version 2)coursenligne.lille1100% (4)
- Hérédité Cytoplasmique PDFДокумент22 страницыHérédité Cytoplasmique PDFcoursenligne.lille1100% (4)
- Optique GéométriqueДокумент51 страницаOptique Géométriquecoursenligne.lille1100% (1)
- AlgologieДокумент17 страницAlgologiecoursenligne.lille194% (17)
- Biotechnologies VégétalesДокумент27 страницBiotechnologies Végétalescoursenligne.lille190% (30)
- Les ChampignonsДокумент8 страницLes Champignonscoursenligne.lille1100% (6)
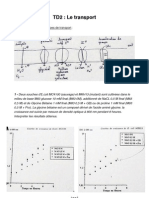
- TD Le TransportДокумент6 страницTD Le Transportcoursenligne.lille1Оценок пока нет
- TD1 Croissance BactérienneДокумент9 страницTD1 Croissance Bactériennecoursenligne.lille1100% (9)
- Fécondation Et GamétogènèseДокумент17 страницFécondation Et Gamétogènèsecoursenligne.lille1Оценок пока нет
- Fécondation Et GamétogènèseДокумент17 страницFécondation Et Gamétogènèsecoursenligne.lille1Оценок пока нет
- TD Milieu de CultureДокумент8 страницTD Milieu de Culturecoursenligne.lille180% (10)
- Transgenèse Végétale Et ApplicationsДокумент16 страницTransgenèse Végétale Et Applicationscoursenligne.lille1100% (13)
- Les GlucidesДокумент21 страницаLes Glucidescoursenligne.lille1100% (3)
- TD Développement Des Ailes de DrosophileДокумент3 страницыTD Développement Des Ailes de Drosophilecoursenligne.lille1100% (1)
- Physiologie RespiratoireДокумент35 страницPhysiologie Respiratoirecoursenligne.lille1100% (16)
- TD Les Applications de La Fusion de ProtoplasteДокумент12 страницTD Les Applications de La Fusion de Protoplastecoursenligne.lille1100% (6)
- Les ProtidesДокумент17 страницLes Protidescoursenligne.lille1100% (1)
- Le Système Nerveux VégétatifДокумент15 страницLe Système Nerveux Végétatifcoursenligne.lille1Оценок пока нет
- TD DiapauseДокумент9 страницTD Diapausecoursenligne.lille1100% (2)
- Mue Et Métamorphose Des InsectesДокумент16 страницMue Et Métamorphose Des Insectescoursenligne.lille1100% (7)
- Métamorphose Des AmphibiensДокумент13 страницMétamorphose Des Amphibienscoursenligne.lille1100% (1)
- L'excrétion RénaleДокумент27 страницL'excrétion Rénalecoursenligne.lille1100% (1)
- Module Niebe DoriДокумент28 страницModule Niebe DoriDEMI MONDE INTERNATIONALОценок пока нет
- Formation de Couches de Protection Et PassivationДокумент7 страницFormation de Couches de Protection Et PassivationabdelbarrОценок пока нет
- La FertilisationДокумент6 страницLa FertilisationBio ecoОценок пока нет
- 51-52 Cem III B 425 N LH SR PMДокумент2 страницы51-52 Cem III B 425 N LH SR PMSakerAbdelazizОценок пока нет
- Thèse LMD 66 2016 PDFДокумент141 страницаThèse LMD 66 2016 PDFLaiadhi DjemouiОценок пока нет
- Appareil UrinaireДокумент3 страницыAppareil UrinaireNissa AlloucheОценок пока нет
- PC1 LDP Lelivrescolaire - FR Ch4 ReactionsOxydorectionДокумент21 страницаPC1 LDP Lelivrescolaire - FR Ch4 ReactionsOxydorectionkasalo6754Оценок пока нет
- Feuille 4 Exercices Atomes Molécules RV PDFДокумент2 страницыFeuille 4 Exercices Atomes Molécules RV PDFblachii valoОценок пока нет
- Devoir Maison-2Документ2 страницыDevoir Maison-2Mohamed MaimounyОценок пока нет
- Cours Thermodynamique PDFДокумент121 страницаCours Thermodynamique PDFConception & Fabrication MécaniqueОценок пока нет
- Synthèse Et Structure Électronique de Phases MAX Et MXènes PDFДокумент153 страницыSynthèse Et Structure Électronique de Phases MAX Et MXènes PDFb5h55b5nz5r5Оценок пока нет
- DNB Blanc - 1 - 2021 - 2022 - CorrigeДокумент1 страницаDNB Blanc - 1 - 2021 - 2022 - Corrigesbarel209Оценок пока нет
- 2 SanogoДокумент53 страницы2 SanogoIbrahima Sory Camara100% (1)
- Nguyen Thi Vi NaДокумент388 страницNguyen Thi Vi NaMatt SowlОценок пока нет
- Calcul de Force de Verrouillage FV (TonneДокумент6 страницCalcul de Force de Verrouillage FV (TonneAnonymous 9qKdViDP450% (4)
- Les Treillis PDFДокумент14 страницLes Treillis PDFMamoudou Sarre100% (1)
- Chapitre-4-Les-Transformations-A-Etat-Solide-Des-Alliages Fe-CДокумент5 страницChapitre-4-Les-Transformations-A-Etat-Solide-Des-Alliages Fe-CWalid Ben AmirОценок пока нет
- Applications Charbon ActifДокумент3 страницыApplications Charbon ActifKhaoula ZefaneОценок пока нет
- Iam Arv NutritionДокумент20 страницIam Arv NutritionsafemindОценок пока нет
- Cours Reacteur CH1Документ38 страницCours Reacteur CH1Mokhtaria ReguigОценок пока нет
- AM3039DДокумент2 страницыAM3039DZeinab Ben RomdhaneОценок пока нет
- RouibehДокумент20 страницRouibehInesОценок пока нет
- Glossairede La Biotechnologie Ang-FrДокумент133 страницыGlossairede La Biotechnologie Ang-Frjaizoz100% (2)
- Stage ScimatДокумент18 страницStage ScimatMouh ChawiОценок пока нет
- Les TP Du LaitДокумент63 страницыLes TP Du LaitZineb BelalaОценок пока нет
- Corrosion Par Les Gaz ChaudsДокумент10 страницCorrosion Par Les Gaz ChaudsFlorian HonfinОценок пока нет
- PrecipitationДокумент5 страницPrecipitationnadpharm13Оценок пока нет
- La Fabrication Du Pain Au LevainДокумент15 страницLa Fabrication Du Pain Au LevainAbde SamadОценок пока нет
- Cours PneuДокумент7 страницCours PneuKhalid TemsamaniОценок пока нет
- Initiation A La Toxicologie V2 PDFДокумент91 страницаInitiation A La Toxicologie V2 PDFFtОценок пока нет