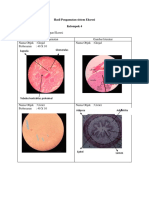
Вам также может понравиться
- 130822423-Tubulasi (1) EmbrioДокумент33 страницы130822423-Tubulasi (1) EmbrioRohmatul AjjaОценок пока нет
- OBAT ALERGIДокумент71 страницаOBAT ALERGINurhidayahОценок пока нет
- Fisiologi Selera MakanДокумент6 страницFisiologi Selera MakanSyukran Ab0% (1)
- BAB II EstrogenДокумент28 страницBAB II EstrogenSam DayОценок пока нет
- MAKALAH BAHASA INDONESIA (II) Siti FaridhaДокумент39 страницMAKALAH BAHASA INDONESIA (II) Siti FaridhaSitiNoerFaridhaОценок пока нет
- Makalah BiokimiaДокумент21 страницаMakalah BiokimiaHenerasia Enhys AnnisaОценок пока нет
- HCG FUNGSI BIOLOGIS DAN APLIKASI KLINISДокумент12 страницHCG FUNGSI BIOLOGIS DAN APLIKASI KLINISelОценок пока нет
- Makalah Antibodi Monoklonal Dan PoliklonalДокумент13 страницMakalah Antibodi Monoklonal Dan PoliklonalSumarlina SoamoleОценок пока нет
- Uji Kandungan UrinДокумент24 страницыUji Kandungan UrinAzhaОценок пока нет
- Respirasi TumbuhanДокумент31 страницаRespirasi TumbuhanHanifОценок пока нет
- Ketikan Regulasi Hormon Adrenal NdutДокумент3 страницыKetikan Regulasi Hormon Adrenal NdutYanie RuhuputtyОценок пока нет
- Review Jurnal Biokimia-Resistensi LeptinДокумент6 страницReview Jurnal Biokimia-Resistensi LeptinSadar Ginting MuntheОценок пока нет
- Makalah HormonДокумент13 страницMakalah Hormonega welliyaniОценок пока нет
- Cascade Hormon System - Rahman KBKДокумент12 страницCascade Hormon System - Rahman KBKRommy Kurniawan DeskyОценок пока нет
- SISTEM ENDOKRINДокумент22 страницыSISTEM ENDOKRINVinka Meilina PutriiОценок пока нет
- OSTEOGENESISДокумент18 страницOSTEOGENESISOlivia NurudhiyaОценок пока нет
- Efek Penuaan pada Sistem Reproduksi PriaДокумент13 страницEfek Penuaan pada Sistem Reproduksi PriaELVIRA RAHMA KARMEILIAОценок пока нет
- Alat Organ Pernapasan Manusia Dan FungsinyaДокумент4 страницыAlat Organ Pernapasan Manusia Dan FungsinyalilisОценок пока нет
- 5 Sistem Tubuh Utama Yang Menyumbang HomeostasisДокумент2 страницы5 Sistem Tubuh Utama Yang Menyumbang HomeostasisTEUKUOKAОценок пока нет
- Kelompok 2 - Hormon Poros Usus-Otak, Mekanisme Lapar KenyangДокумент9 страницKelompok 2 - Hormon Poros Usus-Otak, Mekanisme Lapar KenyangPengisi DompetОценок пока нет
- Fisiologi HewanДокумент26 страницFisiologi HewanFitri AzhariОценок пока нет
- RETIKULARДокумент11 страницRETIKULARNafla Rintisa DaselvrОценок пока нет
- Defisiensi Vitamin A, D, e & K - Yolla F & Zahrotul UДокумент34 страницыDefisiensi Vitamin A, D, e & K - Yolla F & Zahrotul Uzahrotul ulaОценок пока нет
- Laporan Praktikum EndokrinДокумент9 страницLaporan Praktikum EndokrinMuhammad Andhika NurОценок пока нет
- Embriologi Manusia (Kelompok 4)Документ32 страницыEmbriologi Manusia (Kelompok 4)EviОценок пока нет
- Hipofisis AnteriorДокумент11 страницHipofisis AnteriorHarpah NovianiОценок пока нет
- Kel.4 PBR 2010 Protozoa Dan AlgaДокумент70 страницKel.4 PBR 2010 Protozoa Dan AlgaAnni Kholilah100% (1)
- Monohibrid PersilanganДокумент48 страницMonohibrid Persilangannelmian100% (1)
- Wacana TerbaruДокумент8 страницWacana TerbaruArief rahman HОценок пока нет
- Proses Sintesis ProteinДокумент16 страницProses Sintesis ProteinshintaОценок пока нет
- ORGANOGENESISДокумент30 страницORGANOGENESISJD SmithОценок пока нет
- EFEK KALORIGENIKДокумент12 страницEFEK KALORIGENIKpeenОценок пока нет
- HORMONДокумент13 страницHORMONHendrik_NurfitriantoОценок пока нет
- Mutasi Dan Rekombinasi Kel.3Документ15 страницMutasi Dan Rekombinasi Kel.3Rinna AmeliaОценок пока нет
- Hasil pengamatan sistem eksresi dan reproduksiДокумент5 страницHasil pengamatan sistem eksresi dan reproduksiMelianaОценок пока нет
- DIETDMДокумент14 страницDIETDMekahijriantiОценок пока нет
- Sistem Pencernaan: Usus Halus dan Usus BesarДокумент22 страницыSistem Pencernaan: Usus Halus dan Usus BesarJim KhulafaОценок пока нет
- Peranan Zat Besi Dalam Pembentukan Sel DarahДокумент12 страницPeranan Zat Besi Dalam Pembentukan Sel DarahAkuf Suradal Wibisono100% (1)
- SE Domba KambingДокумент14 страницSE Domba KambingRaihan NaufalОценок пока нет
- Suhu Tubuh Kel. 4BДокумент18 страницSuhu Tubuh Kel. 4B20Nurlena duheОценок пока нет
- GASTRULA, TUBULASI DAN BLASTOULAДокумент72 страницыGASTRULA, TUBULASI DAN BLASTOULAZulfa Nabilah JupretОценок пока нет
- Ringkasan Jurnal UtamaДокумент6 страницRingkasan Jurnal UtamaMawar Jelita HatiОценок пока нет
- 4 Metabolisme Lipida. HAL. 6Документ24 страницы4 Metabolisme Lipida. HAL. 6rara fransiskaОценок пока нет
- Pengertian Jaringan Epitelium: Ektoderma, Mesoderma, Atau Endoderma Endotelium Mesotelium Endotelium Mesotelium MesodermaДокумент34 страницыPengertian Jaringan Epitelium: Ektoderma, Mesoderma, Atau Endoderma Endotelium Mesotelium Endotelium Mesotelium MesodermaFahmi FadillahОценок пока нет
- Asam NukleatДокумент90 страницAsam NukleatUmi FazaОценок пока нет
- LKM Sistem Hormon FixДокумент16 страницLKM Sistem Hormon FixGarin Aini100% (1)
- Biosintesis Karbohidrat - RisyadДокумент29 страницBiosintesis Karbohidrat - RisyadRais Salsa MuhammadОценок пока нет
- Sintesis ProteinДокумент29 страницSintesis ProteinAtika RizkiОценок пока нет
- Pengertian EnzimДокумент6 страницPengertian EnzimHansen HartantoОценок пока нет
- Struktur Xylem Sekunder Dan Variasinya - Nabila Dwi Septiana MohuneДокумент15 страницStruktur Xylem Sekunder Dan Variasinya - Nabila Dwi Septiana MohuneAgril Pratama MohuneОценок пока нет
- HISTOLOGI SALURAN KEMIHДокумент4 страницыHISTOLOGI SALURAN KEMIHrizkaОценок пока нет
- Histologi Kelenjar TiroidДокумент1 страницаHistologi Kelenjar TiroidAulia Rahman AmbiyaОценок пока нет
- Kelenjar HipofisisДокумент1 страницаKelenjar HipofisisAjeng FikihОценок пока нет
- Gerak Refleks (KLMPK 7)Документ10 страницGerak Refleks (KLMPK 7)Fauzan SamsОценок пока нет
- Makalah Siklus Estrus - B3Документ10 страницMakalah Siklus Estrus - B3dede lusiОценок пока нет
- Kelompok 2Документ5 страницKelompok 2Sri WahyuningsihОценок пока нет
- Pengaruh Hormon Terhadap MetabolismeeДокумент31 страницаPengaruh Hormon Terhadap Metabolismeed100% (2)
- Diversitas, Konsep Sel Target, Dan ReseptorДокумент39 страницDiversitas, Konsep Sel Target, Dan ReseptornuruldrОценок пока нет
- Biosintesis Sekresi Dan Mekanisme Kerja HormonДокумент26 страницBiosintesis Sekresi Dan Mekanisme Kerja HormonHafsah RiyantiОценок пока нет
- Biosintesis Sekresi Dan Mekanisme Kerja Hormon-With-Cover-Page-V2Документ27 страницBiosintesis Sekresi Dan Mekanisme Kerja Hormon-With-Cover-Page-V2MUHAMMAD HAFIZH201 024Оценок пока нет
- untuk Dokumen Hormon Pelepasan dan SintesisДокумент76 страницuntuk Dokumen Hormon Pelepasan dan SintesisKangjul AripinОценок пока нет
- Referat PrintДокумент55 страницReferat PrintNurhidayahОценок пока нет
- Ira BawahДокумент47 страницIra BawahNurhidayahОценок пока нет
- Edit Kasus 4Документ37 страницEdit Kasus 4Diana Indah LestariОценок пока нет
- HemoptisisДокумент50 страницHemoptisisLp Askep Cuii EatyОценок пока нет
- Isi FixДокумент47 страницIsi FixNurhidayahОценок пока нет
- Kuliah Breast TumorДокумент45 страницKuliah Breast TumorEmallia Phypiet FitrianiОценок пока нет
- Selenium, Zink, Fe (Hapsoro)Документ44 страницыSelenium, Zink, Fe (Hapsoro)NurhidayahОценок пока нет
- Trauma Pada TelingaДокумент16 страницTrauma Pada TelingaNurhidayahОценок пока нет
- 3.diagnosis Dan Penatalaksanaan SCR Comp Medan 2010Документ47 страниц3.diagnosis Dan Penatalaksanaan SCR Comp Medan 2010NurhidayahОценок пока нет
- Pernafasan dan PengendaliannyaДокумент41 страницаPernafasan dan PengendaliannyaNurhidayahОценок пока нет
- Gangguan Sel Darah MerahДокумент50 страницGangguan Sel Darah MerahEmallia Phypiet FitrianiОценок пока нет
- Lembar Pengesahannya FIXДокумент1 страницаLembar Pengesahannya FIXNurhidayahОценок пока нет
- DIAGNOSALEUKEMIAДокумент46 страницDIAGNOSALEUKEMIAEmallia Phypiet FitrianiОценок пока нет
- LIPOPROTEINДокумент43 страницыLIPOPROTEINEmallia Phypiet FitrianiОценок пока нет
- Leptospirosis Kuliah M LuthfiДокумент28 страницLeptospirosis Kuliah M LuthfiNurhidayahОценок пока нет
- Lepto Spiros IsДокумент11 страницLepto Spiros IsNurhidayahОценок пока нет
- Referat PrintДокумент52 страницыReferat PrintNurhidayahОценок пока нет
- Pelayanan Neonatal EsensialДокумент37 страницPelayanan Neonatal EsensialNurhidayahОценок пока нет
- PP Sist Kardiovask, Mod 2.3Документ65 страницPP Sist Kardiovask, Mod 2.3NurhidayahОценок пока нет
- Laporan PBL Blok 235Документ8 страницLaporan PBL Blok 235NurhidayahОценок пока нет
- Infeksi BumilДокумент70 страницInfeksi BumilDeaОценок пока нет
- PP Bahan Diskusi Pakar Sistem Kardiorespiasi, April 12Документ47 страницPP Bahan Diskusi Pakar Sistem Kardiorespiasi, April 12NurhidayahОценок пока нет
- Pertumbuhan &pembelahanДокумент24 страницыPertumbuhan &pembelahanNurhidayahОценок пока нет
- Bio OptikДокумент49 страницBio OptikNurhidayahОценок пока нет
- 2.pengenalan EpilepsiДокумент27 страниц2.pengenalan EpilepsiNurhidayahОценок пока нет
- Epilepsi UmumДокумент20 страницEpilepsi UmumNurhidayahОценок пока нет
- Spiro Chae Tales Bu GersonДокумент28 страницSpiro Chae Tales Bu GersonDeaОценок пока нет
- Pengantar Virus FKДокумент30 страницPengantar Virus FKNurhidayahОценок пока нет