Вам также может понравиться
- Entomologia PDFДокумент326 страницEntomologia PDFJose Diaz100% (1)
- Libro Entomologia AplicadaДокумент326 страницLibro Entomologia AplicadaSonyernesto100% (2)
- Cuestionario de PlatelmitosДокумент65 страницCuestionario de PlatelmitosJUAN PABLO TORRES NUÑEZОценок пока нет
- Informe AnelidosДокумент5 страницInforme Anelidosliz dayanna0% (1)
- Libro Entomologia Aplicada CompressДокумент326 страницLibro Entomologia Aplicada CompressRoberth MendezОценок пока нет
- 9 Filos BiologiaДокумент8 страниц9 Filos BiologiaJaime GutierrezОценок пока нет
- Glosario Cordados PDFДокумент8 страницGlosario Cordados PDFJuliana ArangurenОценок пока нет
- TAREA 3 - ZOO OriginalДокумент114 страницTAREA 3 - ZOO OriginalJohanna Vanessa Ramirez DoñoОценок пока нет
- Guía 7 Reinos de Los Seres Vivos-AnimalДокумент6 страницGuía 7 Reinos de Los Seres Vivos-AnimalARLENIS PAMELA ARIZA PEREIRAОценок пока нет
- Moluscos Anelidos y NematodosДокумент17 страницMoluscos Anelidos y NematodosMariajose MariajoseОценок пока нет
- BIOLOGÍAДокумент2 страницыBIOLOGÍAgleidys colunche livaqueОценок пока нет
- Arthropoda - Wikipedia, La Enciclopedia LibreДокумент80 страницArthropoda - Wikipedia, La Enciclopedia LibreAndrea CuelloОценок пока нет
- Entomologia AplicadaДокумент326 страницEntomologia AplicadaKarina De Avila AguirreОценок пока нет
- Tema 1. IntroducciónДокумент10 страницTema 1. IntroducciónLucero TorradoОценок пока нет
- Practica de Tegumento 2555Документ12 страницPractica de Tegumento 2555anselmo VivasОценок пока нет
- 8° - Taller Invertebrados ResumidoДокумент11 страниц8° - Taller Invertebrados ResumidoKeisy TatianaОценок пока нет
- Actividad CnidariosДокумент8 страницActividad CnidariosGisela PoloОценок пока нет
- Teoria Resumida ReptilesДокумент6 страницTeoria Resumida Reptilesoscar gonzalesОценок пока нет
- Informe 5 ZooДокумент13 страницInforme 5 ZooDiego NapánОценок пока нет
- Otros Unikontes y Animales sin simetría bilateralДокумент5 страницOtros Unikontes y Animales sin simetría bilateralRocioGHОценок пока нет
- Separata Proteccion For I - Entomologia General 2014Документ76 страницSeparata Proteccion For I - Entomologia General 2014Carol Vasquez HuamanОценок пока нет
- ArtrópodosДокумент2 страницыArtrópodosElmerHernanCuroCuroОценок пока нет
- CnidariosДокумент2 страницыCnidariosSamuel Samuel100% (1)
- Informe N°3 PlatelmintosДокумент32 страницыInforme N°3 PlatelmintosLizeth100% (1)
- TaxonomiaДокумент54 страницыTaxonomiawilber rojasОценок пока нет
- Eumetazoos BilateralesДокумент18 страницEumetazoos BilateralesSol DeacónОценок пока нет
- Invertebrados: esponjas y artrópodosДокумент62 страницыInvertebrados: esponjas y artrópodosdjmauro25Оценок пока нет
- MkoinokДокумент8 страницMkoinokJimena OrtizОценок пока нет
- Vertebrados e InvertebradosДокумент19 страницVertebrados e InvertebradosAndres BolivarОценок пока нет
- Lab 12 - LofotrocozoaДокумент14 страницLab 12 - Lofotrocozoajuan aceroОценок пока нет
- TRABAJO PRÁCTICO #6 - Animal y HongosДокумент10 страницTRABAJO PRÁCTICO #6 - Animal y Hongospilar nieva mendozaОценок пока нет
- Narváez A, Ruiz M, 3733. Laboratorio Artropodos CrustáceosДокумент11 страницNarváez A, Ruiz M, 3733. Laboratorio Artropodos CrustáceosMarilyn RuizОценок пока нет
- Diapositivas TEMA 3 en PDFДокумент25 страницDiapositivas TEMA 3 en PDFSilvia Manzanares PeñaОценок пока нет
- Animales 3 (Copia)Документ22 страницыAnimales 3 (Copia)Geovani RodriguezОценок пока нет
- Reconocimiento de Artrópodos en LaboratorioДокумент7 страницReconocimiento de Artrópodos en LaboratorioRafo CastroОценок пока нет
- ArtrópodosДокумент3 страницыArtrópodosRonald Miguel Zúñiga FuentesОценок пока нет
- Zoologia Nº04 - ARTROPODOSДокумент19 страницZoologia Nº04 - ARTROPODOSKarlita ParedesОценок пока нет
- Sinopsis Taxonómica de AnthropodaДокумент9 страницSinopsis Taxonómica de AnthropodaAngie LizethОценок пока нет
- PORIFEROSДокумент7 страницPORIFEROSedgar6433Оценок пока нет
- Características de los cnidariosДокумент19 страницCaracterísticas de los cnidariosIvan FrancoОценок пока нет
- Informe de Laboratorio de Zoologia Phylum CnidarioДокумент6 страницInforme de Laboratorio de Zoologia Phylum Cnidariocarolina palacioОценок пока нет
- Grupo 1 - Práctica 3Документ18 страницGrupo 1 - Práctica 3Cristian Espinoza QuiñonesОценок пока нет
- Reproducción y estructuras de los CnidariosДокумент2 страницыReproducción y estructuras de los CnidariosChismesito a la hora del téОценок пока нет
- ArtropodosДокумент21 страницаArtropodosOsminChvzОценок пока нет
- VDSGДокумент8 страницVDSGDiana PreciadoОценок пока нет
- Características de Los CnidariosДокумент15 страницCaracterísticas de Los CnidariosCelestinaОценок пока нет
- Zoología: Invertebrados y VertebradosДокумент131 страницаZoología: Invertebrados y Vertebradosgerly diazОценок пока нет
- Biologia Animal. Clase 3. Urquiza Micaela.Документ6 страницBiologia Animal. Clase 3. Urquiza Micaela.Micaela UrquizaОценок пока нет
- Evolucion ArtropodosДокумент21 страницаEvolucion ArtropodosCarlos MontoyaОценок пока нет
- Cnidarios y PoriferosДокумент22 страницыCnidarios y PoriferosJuan Diego DoradoОценок пока нет
- Diapositivas TEMA 2Документ15 страницDiapositivas TEMA 2Silvia Manzanares PeñaОценок пока нет
- Informe de InvertebradosДокумент7 страницInforme de InvertebradosWendyVanessaPulgarinOvalleОценок пока нет
- PRACTICA 5 AnelidosДокумент3 страницыPRACTICA 5 Anelidosjeffry2408Оценок пока нет
- QueliceradosДокумент20 страницQueliceradosWero BcОценок пока нет
- Modulo DosДокумент12 страницModulo DosFranyelis HidalgoОценок пока нет
- Taller de Zoología de Invertebrados AcuáticosДокумент7 страницTaller de Zoología de Invertebrados Acuáticosjuan pablo marinОценок пока нет
- BILATERIAДокумент28 страницBILATERIAAndrés SbОценок пока нет
- BiologiaДокумент5 страницBiologiaAna Martinez RodriguezОценок пока нет
- Arañas ArgentinasДокумент160 страницArañas ArgentinasJuanManuelRodríguezRegoОценок пока нет
- Características Generales Del Phylum MolluscaДокумент4 страницыCaracterísticas Generales Del Phylum MolluscaBernardo Alfonso Fajardo LabraОценок пока нет
- JurásicoДокумент13 страницJurásicoBernardo Alfonso Fajardo LabraОценок пока нет
- Orden PezizalesДокумент7 страницOrden PezizalesBernardo Alfonso Fajardo LabraОценок пока нет
- Islas Del Golfo de CaliforniaДокумент14 страницIslas Del Golfo de CaliforniaBernardo Alfonso Fajardo LabraОценок пока нет
- Art 08Документ8 страницArt 08Irene SantosОценок пока нет
- Mandala Movilidad, Músculos de Los Miembros Inferiores.Документ2 страницыMandala Movilidad, Músculos de Los Miembros Inferiores.Walter PiñaОценок пока нет
- Guia FosfolipidosДокумент17 страницGuia FosfolipidosJorgeОценок пока нет
- El Inconsciente SocialДокумент2 страницыEl Inconsciente SocialGore CabreraОценок пока нет
- Memoria Descriptiva - AucayacuДокумент56 страницMemoria Descriptiva - AucayacuTony Quevedo GuimareyОценок пока нет
- Esquistos de Moscovita-CloritoideДокумент12 страницEsquistos de Moscovita-CloritoideDiana Carolina Paz JaramilloОценок пока нет
- Bibliografía RevisadaДокумент6 страницBibliografía RevisadaLuis Pablo Acevedo CaballeroОценок пока нет
- Personalidad, Resiliencia y Calidad de Vida PDFДокумент19 страницPersonalidad, Resiliencia y Calidad de Vida PDFExequiel PorattiОценок пока нет
- Ada 4 PDFДокумент2 страницыAda 4 PDFMICHELLE URBINA ORDOÑEZОценок пока нет
- 10 posiciones sexuales para el orgasmo femeninoДокумент7 страниц10 posiciones sexuales para el orgasmo femeninoHeriberto Mercedes0% (1)
- ea8ACT2 2° 3Документ3 страницыea8ACT2 2° 3ssdОценок пока нет
- Inteligencia emocional en el trabajo: qué es y cómo practicarlaДокумент2 страницыInteligencia emocional en el trabajo: qué es y cómo practicarlaFelipe NahumОценок пока нет
- A Mi También Me Duele El Duelo Propio Del AcompañanteДокумент26 страницA Mi También Me Duele El Duelo Propio Del Acompañantesonia_mata_9Оценок пока нет
- UntitledДокумент20 страницUntitledмοοмƒlοωєяОценок пока нет
- Vih en EmbarazoДокумент18 страницVih en EmbarazoNallely LucioОценок пока нет
- Etapas de La Elaboracion Del PanДокумент11 страницEtapas de La Elaboracion Del PanMelissa Antón CadenillasОценок пока нет
- Dar agua al sedientoДокумент2 страницыDar agua al sedientoalain de lunaОценок пока нет
- Vivero Jaibana La Virginia 1233Документ13 страницVivero Jaibana La Virginia 1233tillo1995Оценок пока нет
- TESIS Jaén RevisadaДокумент63 страницыTESIS Jaén Revisadacecilia diaz tantaricoОценок пока нет
- Ref de CC - NN 8vo 7-Nov-22Документ6 страницRef de CC - NN 8vo 7-Nov-22Evelin RodriguezОценок пока нет
- CatalogoPublicaciones2016 PDFДокумент210 страницCatalogoPublicaciones2016 PDFPascual Bernal OrduñoОценок пока нет
- MBR - Rc-Isu-12 A Rc-Isu-13 - Unidad A.4 CostaДокумент2 страницыMBR - Rc-Isu-12 A Rc-Isu-13 - Unidad A.4 CostaJADER MARTINОценок пока нет
- Introducción A La FisioterapiaДокумент36 страницIntroducción A La FisioterapiaKaren SantanaОценок пока нет
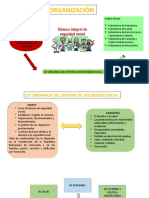
- Seguridad Social y Sistema EconomicoДокумент8 страницSeguridad Social y Sistema EconomicoyusmaryОценок пока нет
- 11-Tips para Ganar Mas DineroДокумент21 страница11-Tips para Ganar Mas DineroAlly Guadalupe Sánchez Alegría100% (1)
- HDS - Resifum U PDFДокумент8 страницHDS - Resifum U PDFedgarОценок пока нет
- Campus Virtual UPAO - Reporte de NotasДокумент2 страницыCampus Virtual UPAO - Reporte de NotasWarmy Plaza HotelОценок пока нет
- 1 3 1 Semana 3 y 4 Manejo Seguro y Manipulacion Del Sistema de Alta TensionДокумент42 страницы1 3 1 Semana 3 y 4 Manejo Seguro y Manipulacion Del Sistema de Alta TensionGonzalo ContrerasОценок пока нет
- Características Del Plano AstralДокумент2 страницыCaracterísticas Del Plano Astraljonh11Оценок пока нет
- Revista GEPU3Документ284 страницыRevista GEPU3Mairin AngelОценок пока нет