Вам также может понравиться
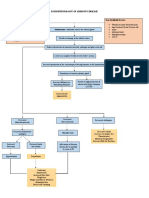
- Introduction To Human Body Anatomy PhysiologyДокумент85 страницIntroduction To Human Body Anatomy PhysiologyTalha AbbasiОценок пока нет
- Support in Plants and AnimalsДокумент2 страницыSupport in Plants and Animalslastjoe71100% (1)
- Anatomy and Physiology Terms: Brief Definitions, Roots & Morphology; An Abecedary; Vol 10 - Digestive System TermsОт EverandAnatomy and Physiology Terms: Brief Definitions, Roots & Morphology; An Abecedary; Vol 10 - Digestive System TermsОценок пока нет
- Chapter 1 A & P Check PointsДокумент11 страницChapter 1 A & P Check PointsRobin Wilfong89% (9)
- Regulation of Blood GlucoseДокумент21 страницаRegulation of Blood GlucoseDimple PatelОценок пока нет
- Uman Hysiology: Integrative Physiology III: ExerciseДокумент23 страницыUman Hysiology: Integrative Physiology III: ExercisepaskalinaОценок пока нет
- Addisons Disease PathophysiologyДокумент2 страницыAddisons Disease PathophysiologyHanna NocumОценок пока нет
- (Chapter 1 and 2) : Anaphy Quiz Reviewer (Chapters 1-3)Документ20 страниц(Chapter 1 and 2) : Anaphy Quiz Reviewer (Chapters 1-3)Mariam GamosОценок пока нет
- Copybook Physiology EnglishДокумент143 страницыCopybook Physiology EnglishamrendraОценок пока нет
- Human Anatomy and Physiology II: Biology 1414 Unit 8 Metabolism and NutritionДокумент103 страницыHuman Anatomy and Physiology II: Biology 1414 Unit 8 Metabolism and Nutritionciroj25Оценок пока нет
- Parts of The Human Eye With DefinitionДокумент4 страницыParts of The Human Eye With DefinitionStarsky Allence Puyoc0% (1)
- BIO 3200 - Human Physiology LCT 1 - Ch1-5Документ9 страницBIO 3200 - Human Physiology LCT 1 - Ch1-5HoreaОценок пока нет
- Metabolism NotesДокумент208 страницMetabolism Notesbairdman89Оценок пока нет
- Anat 5.2 Pelvis and Perineum - ElevazoДокумент14 страницAnat 5.2 Pelvis and Perineum - Elevazolovelots1234Оценок пока нет
- Introduction To Physiology: General Physiology The Cell HomeostasisДокумент38 страницIntroduction To Physiology: General Physiology The Cell HomeostasisAnanta Subedi100% (1)
- Anatomy & Physiology: Exam Study GuideДокумент4 страницыAnatomy & Physiology: Exam Study GuideJessan Ybañez JoreОценок пока нет
- Medicine The American Journal of SportsДокумент5 страницMedicine The American Journal of SportsTaregh KaramiОценок пока нет
- Nervous SystemДокумент71 страницаNervous SystemSyra May PadlanОценок пока нет
- Muscles: Dr. Maung MyintДокумент54 страницыMuscles: Dr. Maung MyintIntan AinОценок пока нет
- The Fate of Metabolism Andmabolic PathwaysДокумент25 страницThe Fate of Metabolism Andmabolic PathwaysXuân Vi100% (1)
- Volkmann's Ischemic ContractureДокумент41 страницаVolkmann's Ischemic ContractureKrishna Madhukar91% (11)
- Muscle Physiology Formal ReportДокумент10 страницMuscle Physiology Formal ReportHermann ManaОценок пока нет
- Lymphatic SystemДокумент70 страницLymphatic SystemNang Maizana Megat Yahya100% (1)
- Biological Systems and Processes BookletДокумент31 страницаBiological Systems and Processes Bookletapi-422428700100% (1)
- CyclemenuДокумент1 страницаCyclemenuapi-308855010100% (1)
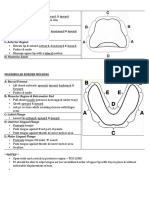
- Border Molding PDFДокумент1 страницаBorder Molding PDFNajeeb UllahОценок пока нет
- Baxarias-Herrerin Atlas of PaleopathologyДокумент300 страницBaxarias-Herrerin Atlas of PaleopathologyNardy100% (1)
- Experiment 7 - Speed of Mechanical WavesДокумент27 страницExperiment 7 - Speed of Mechanical WavesdaОценок пока нет
- Lab 6 ReportДокумент19 страницLab 6 Reportapi-311215267100% (2)
- Cardiovascular Response To ExerciseДокумент18 страницCardiovascular Response To ExerciseChristian JaraОценок пока нет
- Clinton Hackney Anatomy & Physiology II BctcsДокумент65 страницClinton Hackney Anatomy & Physiology II BctcsJack HolthouseОценок пока нет
- Introduction in Human Anatomy and PhysiologyДокумент35 страницIntroduction in Human Anatomy and Physiologyraul nino MoranОценок пока нет
- Exercise LabДокумент3 страницыExercise LabJonatan LeflerОценок пока нет
- Biochemistry: DR - Radhwan M. Asal Bsc. Pharmacy MSC, PHD Clinical BiochemistryДокумент13 страницBiochemistry: DR - Radhwan M. Asal Bsc. Pharmacy MSC, PHD Clinical BiochemistryAnas SeghayerОценок пока нет
- Muscle Tissue (Part 2) : Rate of ContractionДокумент5 страницMuscle Tissue (Part 2) : Rate of ContractionAimi Zafirah Saiful BahriОценок пока нет
- Skeletal Muscle Contraction QuestionsДокумент2 страницыSkeletal Muscle Contraction Questionsgoofy_jess25% (4)
- Short Note Biology Form 5-Chapter 3 Coordination and ResponseДокумент6 страницShort Note Biology Form 5-Chapter 3 Coordination and Responsesalamah_sabri75% (4)
- Sheep Brain Observation LAB 2015Документ4 страницыSheep Brain Observation LAB 2015Leo MatsuokaОценок пока нет
- The Muscular System Learning Enhancement Activity LectureДокумент6 страницThe Muscular System Learning Enhancement Activity LectureNathaniel CornistaОценок пока нет
- Experiment On Cellular Respiration (Using BTB Solution)Документ4 страницыExperiment On Cellular Respiration (Using BTB Solution)Lorraine M. Del RosarioОценок пока нет
- Biochemistry Practice QuestionsДокумент6 страницBiochemistry Practice QuestionsBen BarnesОценок пока нет
- You Are What You EatДокумент9 страницYou Are What You EatRemedios BandongОценок пока нет
- Analysis of An Antacid Lab ReportДокумент5 страницAnalysis of An Antacid Lab ReportClandy CoОценок пока нет
- Handouts in Skeletal SystemДокумент6 страницHandouts in Skeletal SystemShiela Mae EslumОценок пока нет
- Exercise Physiology Lab ReportДокумент11 страницExercise Physiology Lab Reportapi-253201402Оценок пока нет
- Body Fluids Fluid Physiology NotesДокумент19 страницBody Fluids Fluid Physiology NotesBrianОценок пока нет
- Pres1 Intro To KinesiologyДокумент31 страницаPres1 Intro To KinesiologyShahina Sherani100% (2)
- Neuromuscular and Nervous System 2010 With AnswersДокумент13 страницNeuromuscular and Nervous System 2010 With AnswersWenzy CruzОценок пока нет
- Anatomy August 2017Документ356 страницAnatomy August 2017tylermedОценок пока нет
- Chapter 3 Basic Anatomy and PhysiologyДокумент6 страницChapter 3 Basic Anatomy and PhysiologyAini LutfiОценок пока нет
- Exercise PhysiologyДокумент8 страницExercise PhysiologyY Z0% (1)
- Biology EssaysДокумент35 страницBiology Essayst rex422Оценок пока нет
- Lab Report 2 AutosavedДокумент18 страницLab Report 2 Autosavedapi-249482502100% (2)
- The Formula For Aerobic FitnessДокумент1 страницаThe Formula For Aerobic FitnessMelanie Saldivar CapalunganОценок пока нет
- PhysiologyДокумент31 страницаPhysiologyraza20100% (1)
- HomeostasisДокумент5 страницHomeostasisMaria Jayiera Alkiela Pe�alesОценок пока нет
- Chapter 14 - An Introduction To Organic ChemistryДокумент29 страницChapter 14 - An Introduction To Organic ChemistryNabindra RuwaliОценок пока нет
- PL1 3Документ2 страницыPL1 3Dennis Valdez80% (5)
- Bio 142 Exercise Physiology Lab ReportДокумент11 страницBio 142 Exercise Physiology Lab Reportapi-34160259975% (4)
- Freezing Point DepressionДокумент4 страницыFreezing Point DepressionJuan Agustin Garcia TancoОценок пока нет
- 2 MODULE CHAPTER 3&4 AnaPhy 2020-2021 PDFДокумент51 страница2 MODULE CHAPTER 3&4 AnaPhy 2020-2021 PDFJosh MagatОценок пока нет
- Animal Tissues: Epithelial TissueДокумент19 страницAnimal Tissues: Epithelial TissueJnana YumnaОценок пока нет
- BPH First Year Curriculum of Purbanchal UniversityДокумент48 страницBPH First Year Curriculum of Purbanchal Universityseo167Оценок пока нет
- Activity and ExerciseДокумент45 страницActivity and ExerciseAmiel Francisco ReyesОценок пока нет
- The HeartДокумент9 страницThe HeartShiela Belandres MendozaОценок пока нет
- Technical DefinitionДокумент3 страницыTechnical DefinitionErin BarnesОценок пока нет
- Stop-Anemia-In-Migrants WongДокумент8 страницStop-Anemia-In-Migrants Wongapi-308855010Оценок пока нет
- Uc Davis Dining Services Standardized RecipeДокумент2 страницыUc Davis Dining Services Standardized Recipeapi-308855010Оценок пока нет
- Grant Proposal Letter of IntentДокумент2 страницыGrant Proposal Letter of Intentapi-308855010Оценок пока нет
- Nutrient AnalysisДокумент1 страницаNutrient Analysisapi-308855010Оценок пока нет
- Regular DietДокумент1 страницаRegular Dietapi-308855010Оценок пока нет
- HandoutДокумент1 страницаHandoutapi-308855010Оценок пока нет
- Summary SheetДокумент2 страницыSummary Sheetapi-308855010Оценок пока нет
- Group 1c - Pre - and Post-Consumer CompostingДокумент20 страницGroup 1c - Pre - and Post-Consumer Compostingapi-308855010Оценок пока нет
- Diagnosis Dan Tatalaksana Cedera Medula Spinalis Traumatik: Diagnosis and Treatment of Traumatic Spinal Cord InjuryДокумент11 страницDiagnosis Dan Tatalaksana Cedera Medula Spinalis Traumatik: Diagnosis and Treatment of Traumatic Spinal Cord InjuryAna Raniri UtariОценок пока нет
- AFO Orthometry Form OptДокумент1 страницаAFO Orthometry Form OptaryogaОценок пока нет
- Tissue LocationsДокумент4 страницыTissue Locationsrenee belle isturisОценок пока нет
- Byung Kang Pelvis 09.15.2014Документ121 страницаByung Kang Pelvis 09.15.2014Elena ConstantinОценок пока нет
- Nervous System Examination SynopsisДокумент26 страницNervous System Examination Synopsisaarti chandeОценок пока нет
- Introduction To Brain Anatomy: Wieslaw L. NowinskiДокумент37 страницIntroduction To Brain Anatomy: Wieslaw L. NowinskiAditya Rama DevaraОценок пока нет
- Carpal Tunnel Surgery, Hand Clinics, Volume 18, Issue 2, Pages 211-368 (May 2002)Документ151 страницаCarpal Tunnel Surgery, Hand Clinics, Volume 18, Issue 2, Pages 211-368 (May 2002)alinutza_childОценок пока нет
- Integumentary System NotesДокумент6 страницIntegumentary System NotesheeueuОценок пока нет
- CW El CuerpoДокумент2 страницыCW El CuerpovbllchckОценок пока нет
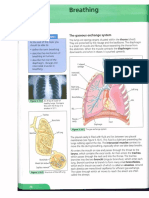
- Csec Study Guide BreathingДокумент2 страницыCsec Study Guide BreathingLynda BarrowОценок пока нет
- Age by Dental DataДокумент40 страницAge by Dental DataAnanda KumarОценок пока нет
- Low Back Pain-Orthoinfo - AaosДокумент13 страницLow Back Pain-Orthoinfo - AaosBang mantoОценок пока нет
- Sense of Hearing and EquilibriumДокумент21 страницаSense of Hearing and EquilibriumHamza KhanОценок пока нет
- Epithelial Tissue (Finale)Документ2 страницыEpithelial Tissue (Finale)Adz Na HarОценок пока нет
- Grade 9 Circulatory SystemДокумент5 страницGrade 9 Circulatory SystemCes GarceraОценок пока нет
- Final LMRДокумент118 страницFinal LMRPrabhat KcОценок пока нет
- Lecture Medskull - ModofiedДокумент88 страницLecture Medskull - ModofiedNikolai SamaniegoОценок пока нет
- Balancing The Sex Hormones Class 19Документ6 страницBalancing The Sex Hormones Class 19RukaphuongОценок пока нет
- Spinal Manual 20 August 2004Документ235 страницSpinal Manual 20 August 2004Forum PompieriiОценок пока нет
- Shivanshu Gangwar M. Pharma (Pharmaceutics)Документ16 страницShivanshu Gangwar M. Pharma (Pharmaceutics)Shibbu GangwarОценок пока нет
- Biological Basis of PersonalityДокумент58 страницBiological Basis of PersonalityAldrian Maulion EvangelistaОценок пока нет
- Dental and Oral Cavity: Andrea Van de WeteringДокумент11 страницDental and Oral Cavity: Andrea Van de WeteringfatimahОценок пока нет
- Beginner & Advanced Rotator Cuff Exercises For Shoulder Pain - MyodetoxДокумент18 страницBeginner & Advanced Rotator Cuff Exercises For Shoulder Pain - MyodetoxJustin Wigon100% (1)
- Developing ToothДокумент3 страницыDeveloping ToothZHIARA MAE FACUNОценок пока нет