Вам также может понравиться
- Manual de ParasitologiaДокумент30 страницManual de Parasitologiaanalistaensistemas0% (1)
- Disciplina PositivaДокумент2 страницыDisciplina PositivaLisbeth Ponce CallenovaОценок пока нет
- Violencia de Género en Las EscuelasДокумент6 страницViolencia de Género en Las Escuelasprueba365Оценок пока нет
- Proyecto Espacio Aventura DiminutaДокумент28 страницProyecto Espacio Aventura DiminutaEsther VazquezОценок пока нет
- Creatividad - Libro Pienso 3Документ4 страницыCreatividad - Libro Pienso 3marco avendaño guzmanОценок пока нет
- Puerto PlataДокумент96 страницPuerto Platapedro perezОценок пока нет
- La Pedagogía Del Oprimido y La Pedagogía LiberadoraДокумент5 страницLa Pedagogía Del Oprimido y La Pedagogía LiberadoraDanielaОценок пока нет
- Proyecto de Desarrollo Estratégico de Los Recursos NaturalesДокумент20 страницProyecto de Desarrollo Estratégico de Los Recursos NaturalesRoger Hernan Rojas AmaroОценок пока нет
- Una Estrategia de Asesoramiento para Promover La Disciplina PositivaДокумент1 страницаUna Estrategia de Asesoramiento para Promover La Disciplina PositivaCarmen MelladoОценок пока нет
- Moleculas de Adhesion Celular 1223605666724403 9Документ68 страницMoleculas de Adhesion Celular 1223605666724403 9JoseioОценок пока нет
- Presentación de La FamiliaДокумент6 страницPresentación de La FamiliaNicole100% (1)
- Las Vitaminas y CoenzimasДокумент13 страницLas Vitaminas y Coenzimasanacarolinne2014Оценок пока нет
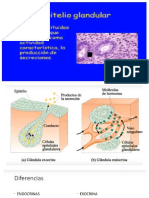
- Glándulas ExocrinasДокумент2 страницыGlándulas ExocrinasDulce MoralesОценок пока нет
- Ser Padres Hoy Un RetoДокумент10 страницSer Padres Hoy Un RetopatriciaОценок пока нет
- Principios de La Igualdad NaturalДокумент6 страницPrincipios de La Igualdad NaturalLia CamachoОценок пока нет
- El Bullying y Sus Efectos en El Exito EscolarДокумент10 страницEl Bullying y Sus Efectos en El Exito EscolargersonОценок пока нет
- Elementos Necesarios para Plantear Un Problema de Investigación CientíficaДокумент27 страницElementos Necesarios para Plantear Un Problema de Investigación Científicaa_osoriod@yahoo.esОценок пока нет
- Normas Del Sistema Educativo Dominicano para La Convivencia y La Disciplina Escolar en Los Centros Educativos Públicos y PrivadosДокумент1 страницаNormas Del Sistema Educativo Dominicano para La Convivencia y La Disciplina Escolar en Los Centros Educativos Públicos y PrivadosJhojanny RoblesОценок пока нет
- Lista de Cotejo para Supervisión UGELДокумент4 страницыLista de Cotejo para Supervisión UGELKevinAlvarezОценок пока нет
- Rafael Barret TPДокумент12 страницRafael Barret TPpablorlandog2788Оценок пока нет
- Todos Somos IgualesДокумент2 страницыTodos Somos IgualesKaren VascoОценок пока нет
- Condicionamiento ClásicoДокумент7 страницCondicionamiento ClásicoBryanCarbonelTejadaОценок пока нет
- Enfermedades GranulomatosasДокумент13 страницEnfermedades GranulomatosasEstela Chumbiray0% (1)
- Presentación Taller La Familia PDFДокумент24 страницыPresentación Taller La Familia PDFRosmar Lopez100% (2)
- Fibras ElásticasДокумент2 страницыFibras ElásticasShany Barrios100% (1)
- Its, Vih-Sida en La EscuelaДокумент2 страницыIts, Vih-Sida en La EscuelaLalaFriasОценок пока нет
- Amor, Matrimonio y FamiliaДокумент24 страницыAmor, Matrimonio y FamiliafabisoleОценок пока нет
- Ficha Del Alumno Primaria 2011 Drec Ysela LazoДокумент1 страницаFicha Del Alumno Primaria 2011 Drec Ysela LazoCesar JuarezОценок пока нет
- Disciplina Positiva DiapositivasДокумент6 страницDisciplina Positiva DiapositivasOndinaMaldonado100% (1)
- Control de EstímulosДокумент18 страницControl de EstímulosalbertoОценок пока нет
- Clima EscolarДокумент1 страницаClima EscolarmemmomiriamОценок пока нет
- Conjugación BacterianaДокумент6 страницConjugación BacterianaMicaela PerezОценок пока нет
- ProteoglicanosДокумент7 страницProteoglicanosLina Montaño100% (1)
- Cuento Estrategia Mejorar Lectoescritura AlumnosДокумент28 страницCuento Estrategia Mejorar Lectoescritura Alumnosdavidt505@hotmail.com0% (1)
- Tesis de Evaluacion de EdificacionesДокумент122 страницыTesis de Evaluacion de EdificacionesigaradiОценок пока нет
- Glándulas HISTOLOGIAДокумент33 страницыGlándulas HISTOLOGIALiseth HumalaОценок пока нет
- Todos Somos Iguales Con Nuestras DiferenciasДокумент2 страницыTodos Somos Iguales Con Nuestras DiferenciasSergio Miranda López100% (1)
- Sindrome de MarfanДокумент2 страницыSindrome de Marfandakito181292Оценок пока нет
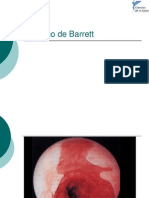
- Esofago de BarretДокумент10 страницEsofago de BarretThiago SantosОценок пока нет
- Sopa de LetrasДокумент2 страницыSopa de LetrasmisteriolkОценок пока нет
- Cadera y RodillaДокумент2 страницыCadera y RodillaJose CJОценок пока нет
- El Semáforo de ConductaДокумент3 страницыEl Semáforo de ConductaGuerrero Campos ClajuОценок пока нет
- Las Relaciones Interactivas en Clase Lunes 22 AbrilДокумент0 страницLas Relaciones Interactivas en Clase Lunes 22 AbrilEsteban BeltranОценок пока нет
- Niebla y NeblinaДокумент13 страницNiebla y NeblinaAlexander EspinozaОценок пока нет
- Pautas Elementales de Modificación de ConductaДокумент1 страницаPautas Elementales de Modificación de Conductamatildedelatorre_muriedasОценок пока нет
- Test Personalidad OjosДокумент3 страницыTest Personalidad OjosAnthony MacbrayanОценок пока нет
- Qué Es La ElastinaДокумент2 страницыQué Es La Elastinadesiree castilloОценок пока нет
- CorrespondenciaДокумент5 страницCorrespondenciaJESUSОценок пока нет
- Caso Clinico Terapia ConcdutualДокумент6 страницCaso Clinico Terapia ConcdutualBohane SantosОценок пока нет
- Guia Censo NeaeДокумент14 страницGuia Censo NeaejsabioteОценок пока нет
- 130 Propuestas para Trabajar La CoeducaciónДокумент32 страницы130 Propuestas para Trabajar La Coeducacióncarmenpalma100% (1)
- Tesis, Ansiedad FinalДокумент46 страницTesis, Ansiedad FinalAntonio RojasОценок пока нет
- PRESENTACION Promoción Salud MentalДокумент7 страницPRESENTACION Promoción Salud MentalLucía PuenteОценок пока нет
- Enfermedades GranulomatosasДокумент50 страницEnfermedades GranulomatosasOscar HurtadoОценок пока нет
- Intervencion Psicopedagogica y Farmacologica Ante Los Trastornos Del Comportamiento de Inicio en La Infancia y AdolescenciaДокумент13 страницIntervencion Psicopedagogica y Farmacologica Ante Los Trastornos Del Comportamiento de Inicio en La Infancia y AdolescenciaLiiZz Díaz DíazОценок пока нет
- Esófago de BarrettДокумент19 страницEsófago de BarrettRamon RvОценок пока нет
- Matriz ExtracelularДокумент36 страницMatriz ExtracelularArmando BravoОценок пока нет
- Moléculas de Adhesión CelularДокумент7 страницMoléculas de Adhesión CelularAbel Gallegos AguilarОценок пока нет
- Adherencia CelularДокумент14 страницAdherencia CelularAndy Paúl García OrbeОценок пока нет
- Tema 3Документ7 страницTema 3letiОценок пока нет
- Interacción Entre Las Células y Su EntornoДокумент8 страницInteracción Entre Las Células y Su EntornoAnita EspinosaОценок пока нет
- HongosДокумент2 страницыHongosKaren CabanaОценок пока нет
- 149lombriabono AmeДокумент37 страниц149lombriabono AmeKaren CabanaОценок пока нет
- Practicaibioest 2016Документ7 страницPracticaibioest 2016Karen CabanaОценок пока нет
- Para Metros 2Документ5 страницPara Metros 2Karen CabanaОценок пока нет
- Imarpe 6) Area de Proteccion de Chanque-Tacna 1007Документ22 страницыImarpe 6) Area de Proteccion de Chanque-Tacna 1007Karen CabanaОценок пока нет
- Practica 10 MitosisДокумент4 страницыPractica 10 MitosisKaren CabanaОценок пока нет
- Desechos de La Industria PesqueraДокумент3 страницыDesechos de La Industria PesqueraKaren Cabana0% (1)