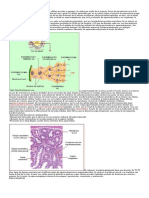
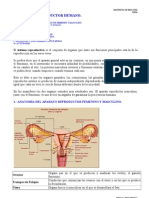
Вам также может понравиться
- Espermatogenesis y OvogenesisДокумент5 страницEspermatogenesis y OvogenesisJorge Navarro100% (1)
- Ovogénesis (WIkipediaДокумент4 страницыOvogénesis (WIkipediaFernandoОценок пока нет
- Informe GametogenesisДокумент16 страницInforme GametogenesisJonathan MejiaОценок пока нет
- Ciclo EstralДокумент9 страницCiclo EstralSary RuizОценок пока нет
- GametogénesisДокумент7 страницGametogénesisMonics GonzalezОценок пока нет
- BH - Guía de Estudio #1Документ13 страницBH - Guía de Estudio #1mczanaboni100% (1)
- EmbriologiaДокумент15 страницEmbriologiaMarijoОценок пока нет
- ESPERMATOGENESISДокумент15 страницESPERMATOGENESISMiriam E. MairimОценок пока нет
- 3 Citogenetica ReproductivaДокумент13 страниц3 Citogenetica Reproductivajustina galeanoОценок пока нет
- GAMETOGENESIS ExposicionДокумент5 страницGAMETOGENESIS ExposicionKEITLYN BENTHAMS SILVA100% (1)
- Reproducción AsexualДокумент7 страницReproducción Asexualaxel avendañoОценок пока нет
- Reproducción Sexual-1Документ25 страницReproducción Sexual-1Yelena ReverolОценок пока нет
- Universidad de Las Fuerzas Armadas Esp1Документ4 страницыUniversidad de Las Fuerzas Armadas Esp1kateryneОценок пока нет
- Desarrollo embrionario desde la fecundaciónДокумент21 страницаDesarrollo embrionario desde la fecundaciónJose Ignacio Vera GutierrezОценок пока нет
- Ramirez Carbajal123Документ21 страницаRamirez Carbajal123Cristhofer lamdon Ramirez carbajalОценок пока нет
- 2.4 Gametogenesis Animal y VegetalДокумент28 страниц2.4 Gametogenesis Animal y VegetalJosselyn Jeannette Lopez88% (24)
- Historia de la embriologíaДокумент15 страницHistoria de la embriologíaRoselys HiguereyОценок пока нет
- OntogeniaДокумент18 страницOntogeniaHéctor Alejandro Grau Díaz de BedoyaОценок пока нет
- BiologiaДокумент5 страницBiologiafabianaguilar1219Оценок пока нет
- OvogénesisДокумент4 страницыOvogénesisHevel Hard M VОценок пока нет
- GametogénesisДокумент3 страницыGametogénesisAlessia BernaОценок пока нет
- Espermatogénesis Material de ApoyoДокумент9 страницEspermatogénesis Material de ApoyoEdgar Leonardo CaprielОценок пока нет
- OvogenesisyespermatogenesisДокумент9 страницOvogenesisyespermatogenesisMirella LuyoОценок пока нет
- Game To GenesisДокумент5 страницGame To GenesisAna Marilyn Javier CastellanosОценок пока нет
- Sexo Pudor y Poses SexualesДокумент7 страницSexo Pudor y Poses SexualesLeonardo HinojosaОценок пока нет
- EmbriologíaДокумент23 страницыEmbriologíaMARIANAОценок пока нет
- Tarea03 - Proceso de La Ovogénesis y Fecundación - Huaranga AlvarezДокумент9 страницTarea03 - Proceso de La Ovogénesis y Fecundación - Huaranga AlvarezMilagros H. AlvarezОценок пока нет
- Gametogénesis, Ovulación e ImplantaciónДокумент31 страницаGametogénesis, Ovulación e ImplantaciónKeren Aquino BrunoОценок пока нет
- ESPERMATOGÉNESISДокумент4 страницыESPERMATOGÉNESISJacqueline La Rosa LeónОценок пока нет
- Gametogénesis: Ovogénesis y espermatogénesisДокумент12 страницGametogénesis: Ovogénesis y espermatogénesisTannia diazОценок пока нет
- EspermatogenesisДокумент7 страницEspermatogenesismariannyr13.02Оценок пока нет
- Ovogenesis, Espermatogenesis y PubertadДокумент12 страницOvogenesis, Espermatogenesis y PubertadLluvia CardenasОценок пока нет
- EmbriologiaДокумент14 страницEmbriologiaDario Grisales BОценок пока нет
- Biologia de La ConcepcionДокумент14 страницBiologia de La ConcepcionLiss Espana100% (2)
- NOveno 01 Clase EstudianteДокумент3 страницыNOveno 01 Clase EstudianteVentassam SamОценок пока нет
- Plan de Mejoramiento Biologia SeptimoДокумент4 страницыPlan de Mejoramiento Biologia SeptimoJosé Nicolás Díaz VelásquezОценок пока нет
- Reproduccion HumanaДокумент14 страницReproduccion HumanaArbelio Vásquez JíguanОценок пока нет
- Gametogénesis, ovogénesis y meiosisДокумент12 страницGametogénesis, ovogénesis y meiosisjhonnyromeroОценок пока нет
- La Gametogénesis Es La Formación de Gametos Por Medio de La MeiosisДокумент2 страницыLa Gametogénesis Es La Formación de Gametos Por Medio de La MeiosisNanosinx Kress UnityОценок пока нет
- EspermatogenesisДокумент5 страницEspermatogenesisJuan Jose Garcia MoralesОценок пока нет
- Cuestionario EmbriologiaДокумент12 страницCuestionario EmbriologiaClariandna PeraltaОценок пока нет
- Reproduccion y DesarrolloДокумент10 страницReproduccion y DesarrollonatОценок пока нет
- gametogeenesisДокумент18 страницgametogeenesisrossipe2007Оценок пока нет
- GametogenesisДокумент28 страницGametogenesisReinaldo Chirinos ArredondoОценок пока нет
- Taller Reproducción en HumanosДокумент5 страницTaller Reproducción en HumanosLorenitaLäväöОценок пока нет
- Nociones de EmbriologiaДокумент11 страницNociones de EmbriologiaOriana Hnnd'z-m0% (1)
- Reproduccion HumanaДокумент6 страницReproduccion HumanaMichell ReyesОценок пока нет
- El Sistema Reproductor HumanoДокумент9 страницEl Sistema Reproductor HumanoMaryAngelОценок пока нет
- Reproduccion de Los Seres VivosДокумент16 страницReproduccion de Los Seres VivosNaldy MaritzaОценок пока нет
- Ovogenesis y maduración del ovocito en 40Документ3 страницыOvogenesis y maduración del ovocito en 40Leo ArellanoОценок пока нет
- Tema 1 - GametogénesisДокумент7 страницTema 1 - GametogénesisMarc Biosca CalabuigОценок пока нет
- GametogenesisДокумент38 страницGametogenesisEripatitreviОценок пока нет
- Actividad 1Документ10 страницActividad 1Alan Rdz SalasОценок пока нет
- Formación de gametos y reproducción sexualДокумент7 страницFormación de gametos y reproducción sexualponz0804100% (1)
- EmbriologíaДокумент44 страницыEmbriologíaLenis C Vasques100% (1)
- Biologia - Sem7 - Fecundación Humana y ConcepciónДокумент17 страницBiologia - Sem7 - Fecundación Humana y ConcepciónDomenica MatuteОценок пока нет
- Elementos OntogénicosДокумент10 страницElementos Ontogénicosjuancarlos307197750% (2)
- Practica 4 - Ind 3216Документ3 страницыPractica 4 - Ind 3216PedroEstebanQuisbertОценок пока нет
- Introducción Cuyes ..... CleoДокумент44 страницыIntroducción Cuyes ..... CleohenrryОценок пока нет
- Masticacion Clinica de NiñoДокумент18 страницMasticacion Clinica de NiñoBibi MoleОценок пока нет
- Ficha Técnica KIKKO 2021Документ2 страницыFicha Técnica KIKKO 2021jlogisticaОценок пока нет
- Estandarizacion EngranajesДокумент4 страницыEstandarizacion EngranajesArmandoОценок пока нет
- Orden de mérito estudiantes Ciencias de la ComunicaciónДокумент45 страницOrden de mérito estudiantes Ciencias de la Comunicaciónpamela0% (2)
- Edicon Fluidos PDFДокумент29 страницEdicon Fluidos PDFGino StalinОценок пока нет
- El Sistema SolarДокумент3 страницыEl Sistema SolarGrover Yamil Flores CórdovaОценок пока нет
- Cuadro Comparativo Ley Antievasion IIДокумент127 страницCuadro Comparativo Ley Antievasion IIJess TelloОценок пока нет
- Capitulo 4Документ52 страницыCapitulo 4Maria SarayОценок пока нет
- Functional Programming With Elixir EsДокумент29 страницFunctional Programming With Elixir Eselkinlon0930Оценок пока нет
- 4R70W Gran Marquis PDFДокумент285 страниц4R70W Gran Marquis PDFRichard Medrano Almaraz67% (9)
- Carros NuevosДокумент6 страницCarros NuevosAndres Moreno SandovalОценок пока нет
- Examen Final Dinamica 2021-2Документ7 страницExamen Final Dinamica 2021-2Sergio Fernandez quispeОценок пока нет
- ANÁLISIS ManovaДокумент4 страницыANÁLISIS ManovaKaren ContrerasОценок пока нет
- Ficha de Aplicacion - Julio 3 A SДокумент120 страницFicha de Aplicacion - Julio 3 A Sgerald augusto lopez bardales100% (2)
- Poemas CortosДокумент24 страницыPoemas CortosArmand MartinezОценок пока нет
- Naval de Nasa PDFДокумент133 страницыNaval de Nasa PDFdaniilОценок пока нет
- Taller Operación Pixel A PixelДокумент15 страницTaller Operación Pixel A PixelIfran SierraОценок пока нет
- Caso Clinico EscleritisДокумент5 страницCaso Clinico EscleritislauraОценок пока нет
- Tarea 1 Leyde CoulombДокумент2 страницыTarea 1 Leyde Coulombdaniel ortizОценок пока нет
- Sueños y amigos: cuento sobre la amistadДокумент4 страницыSueños y amigos: cuento sobre la amistaddeicy carvajalОценок пока нет
- Determinación de cenizas en alimentos: Instructivo I2Документ3 страницыDeterminación de cenizas en alimentos: Instructivo I2Hector LuceroОценок пока нет
- Lista de MaterialesДокумент8 страницLista de MaterialesMiguel A. Martinez PОценок пока нет
- Stanford InnovacionesДокумент30 страницStanford InnovacionesRené Pedroza0% (1)
- Huesos de la cara: Maxilar superiorДокумент18 страницHuesos de la cara: Maxilar superiorKarenОценок пока нет
- Formatos de Actividades de Aprendizajes MINEDUДокумент3 страницыFormatos de Actividades de Aprendizajes MINEDUESTEPHANY CRISTHEL NEGRON CHACONОценок пока нет
- Estrategia de Producto PDFДокумент41 страницаEstrategia de Producto PDFwarxela100% (3)
- Cristalografía y MineralogíaДокумент24 страницыCristalografía y MineralogíaKevin ChavezОценок пока нет
- EXPO (BJT) Transistor de Unión BipolarДокумент3 страницыEXPO (BJT) Transistor de Unión Bipolarjesus natoОценок пока нет