Вам также может понравиться
- PruebaДокумент11 страницPruebaGustavo Barrera LindaoОценок пока нет
- Caso Practico de Auditoria FinancieraДокумент53 страницыCaso Practico de Auditoria FinancieraRafafel Jorge Ucharico Uruchi84% (102)
- Guia 01 - Deberes FormalesДокумент40 страницGuia 01 - Deberes FormalesLuis Santos Enríquez0% (1)
- Banco de Preguntas CiudadaníaДокумент7 страницBanco de Preguntas CiudadaníaKenedRomanОценок пока нет
- 5 Brasil Colonia Portuguesa 119689770766965 2Документ8 страниц5 Brasil Colonia Portuguesa 119689770766965 2Gustavo Barrera LindaoОценок пока нет
- Plan Anual 10 Egb - 2016Документ11 страницPlan Anual 10 Egb - 2016hemelendez100Оценок пока нет
- Guía completa derivadas ejercicios resueltos propuestosДокумент14 страницGuía completa derivadas ejercicios resueltos propuestosClau TorrejónОценок пока нет
- 361 2012D RHH353 IessДокумент18 страниц361 2012D RHH353 IessGustavo Barrera LindaoОценок пока нет
- Guía completa derivadas ejercicios resueltos propuestosДокумент14 страницGuía completa derivadas ejercicios resueltos propuestosClau TorrejónОценок пока нет
- Unidad IДокумент43 страницыUnidad IGustavo Barrera LindaoОценок пока нет
- Banco de Preguntas CiudadaníaДокумент7 страницBanco de Preguntas CiudadaníaKenedRomanОценок пока нет
- Ejerciciosresueltosdetributaria 121122145543 Phpapp01Документ16 страницEjerciciosresueltosdetributaria 121122145543 Phpapp01Gustavo Barrera LindaoОценок пока нет
- Imperialismo y sus causasДокумент16 страницImperialismo y sus causasjavoluceroОценок пока нет
- La Diferencia Entre Vencedores y PerdedoresДокумент15 страницLa Diferencia Entre Vencedores y Perdedoresgonzo1484100% (1)
- Conquista América siglo XVIДокумент18 страницConquista América siglo XVIGustavo Barrera LindaoОценок пока нет
- Guía para elaborar el Anexo de Gastos Personales en líneaДокумент28 страницGuía para elaborar el Anexo de Gastos Personales en línealib3llula2210Оценок пока нет
- CIUDADANIAДокумент2 страницыCIUDADANIAGustavo Barrera LindaoОценок пока нет
- Marketing MixДокумент40 страницMarketing MixslipkbetoОценок пока нет
- Talller de La Guerra Frc3adaДокумент1 страницаTalller de La Guerra Frc3adaGustavo Barrera LindaoОценок пока нет
- Marketing MixДокумент40 страницMarketing MixslipkbetoОценок пока нет
- Bancodepreguntasexamenremedialfisica 150325100528 Conversion Gate01 PDFДокумент5 страницBancodepreguntasexamenremedialfisica 150325100528 Conversion Gate01 PDFGustavo Barrera LindaoОценок пока нет
- T-ESPE-021513-5 Marketin Mix PDFДокумент65 страницT-ESPE-021513-5 Marketin Mix PDFGary OucОценок пока нет
- Elmovimientoobrero 111205052144 Phpapp02Документ19 страницElmovimientoobrero 111205052144 Phpapp02Gustavo Barrera LindaoОценок пока нет
- Proyecto-De Arte DramáticoДокумент50 страницProyecto-De Arte DramáticoGustavo Barrera Lindao67% (3)
- Pruebadehistoria 7Документ3 страницыPruebadehistoria 7Juan Jose HidalgoОценок пока нет
- Almacenamiento GeneralidadesДокумент41 страницаAlmacenamiento GeneralidadesgeorgevandykeОценок пока нет
- Pruebaimperialismocolonialismoygranguerra 141031180025 Conversion Gate02 PDFДокумент6 страницPruebaimperialismocolonialismoygranguerra 141031180025 Conversion Gate02 PDFGustavo Barrera LindaoОценок пока нет
- Ecuaciones exponencialesДокумент1 страницаEcuaciones exponencialesGustavo Barrera LindaoОценок пока нет
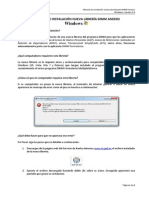
- Manual Instalacion Libreria Windows V1Документ2 страницыManual Instalacion Libreria Windows V1Javier CuzcoОценок пока нет
- Cultura IslámicaДокумент21 страницаCultura IslámicaGustavo Barrera Lindao100% (1)
- Cuadernillo 21 Desde El 19 Al 30 de OCTUBRE MEDIO MAYORДокумент16 страницCuadernillo 21 Desde El 19 Al 30 de OCTUBRE MEDIO MAYORvanessa olmos sОценок пока нет
- Adolf Hitler Tenía RazónДокумент2 страницыAdolf Hitler Tenía RazónJ'Car Gaez RouzОценок пока нет
- Cubo DrenДокумент8 страницCubo DrenAlexis Riveros CortesОценок пока нет
- Usted S A PDFДокумент28 страницUsted S A PDFNabil A M SilvaОценок пока нет
- 3° Básico Historia Guía 3 Ciudades GriegasДокумент6 страниц3° Básico Historia Guía 3 Ciudades Griegasdebyta salinasОценок пока нет
- Discurso Del Sr. Cmdte. Ascenso-2013Документ4 страницыDiscurso Del Sr. Cmdte. Ascenso-2013Farid Gamarra Floreano82% (11)
- Chile Indomito PDFДокумент56 страницChile Indomito PDFAle T.JОценок пока нет
- Demanda MarianaДокумент7 страницDemanda MarianaMARIANA DE BOJORQUEZ 1330Оценок пока нет
- Ripley Grupo10Документ40 страницRipley Grupo10Felipe Brante100% (1)
- KOPPITZДокумент22 страницыKOPPITZLesly Noemi Santisteban SalvadorОценок пока нет
- Listas ProvisionalesДокумент6 страницListas ProvisionalesNieves García MoránОценок пока нет
- Correos 2022 1.3.1.1 Carta OrdinariaДокумент7 страницCorreos 2022 1.3.1.1 Carta OrdinariaJASPОценок пока нет
- Katherina - Rodriguez - Tarea S3Документ5 страницKatherina - Rodriguez - Tarea S3Katherina RodriguezОценок пока нет
- Guia Formulario IseДокумент9 страницGuia Formulario IseNELSA SAENZ SAAVEDRAОценок пока нет
- Influencia Fases Lun@rsДокумент4 страницыInfluencia Fases Lun@rssusana maria hernandez urdanetaОценок пока нет
- 01 Lost and Found - Elle CaseyДокумент394 страницы01 Lost and Found - Elle CaseyclaudiaОценок пока нет
- El Sector de Las TICs en ColombiaДокумент155 страницEl Sector de Las TICs en ColombiaPromoMadridОценок пока нет
- Etica y Estilo Forense PDFДокумент75 страницEtica y Estilo Forense PDFJanice Vargas HinojosaОценок пока нет
- Baño ElectroquimicoДокумент3 страницыBaño ElectroquimicoLuis GustavoОценок пока нет
- ReciclajeДокумент29 страницReciclajeSebax Gamer10Оценок пока нет
- GAFA - LENTE CLARO UVEX I-Vo 9160Документ1 страницаGAFA - LENTE CLARO UVEX I-Vo 9160JORGE ALBERTO PEREZ OSPINAОценок пока нет
- Suelo y TopografiaДокумент12 страницSuelo y TopografiaFabiola Pinto GrandaОценок пока нет
- Posicion EsДокумент4 страницыPosicion EsMaJo RiveraОценок пока нет
- Diagnóstico Diferencial, Evaluacion, EpidemiologiaДокумент12 страницDiagnóstico Diferencial, Evaluacion, EpidemiologiaChriztian Serón TuerosОценок пока нет
- Chikunguña en PediatríaДокумент6 страницChikunguña en Pediatríaangela abrilОценок пока нет
- Compressed Air Filters: Atlas CopcoДокумент30 страницCompressed Air Filters: Atlas Copcoeng_m_ismaileОценок пока нет
- Nro7072 - 2022 06 27Документ40 страницNro7072 - 2022 06 27Diario Primera LíneaОценок пока нет
- La decoración geométrica de los zócalos andalusíesДокумент114 страницLa decoración geométrica de los zócalos andalusíesAnonymous IMjVlUTVОценок пока нет
- ExposiciónДокумент21 страницаExposiciónLuisОценок пока нет
- CoquelucheДокумент9 страницCoquelucheHermes VJОценок пока нет