Вам также может понравиться
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (119)
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (399)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2219)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (344)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (890)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- NaBH4 CuCl2 ReductionOfNitrostyrenes ThesisДокумент63 страницыNaBH4 CuCl2 ReductionOfNitrostyrenes ThesisjoebobОценок пока нет
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- Major Radiation Exposure - What To Expect and How To RespondДокумент8 страницMajor Radiation Exposure - What To Expect and How To RespondMiguel Angel VegaОценок пока нет
- 101 Science Experiments (Gnv64)Документ155 страниц101 Science Experiments (Gnv64)Bogdan Raul100% (1)
- Vitamin D in Dermatology PDFДокумент373 страницыVitamin D in Dermatology PDFelfiana100% (1)
- Cell DifferentiationДокумент26 страницCell DifferentiationQusai Haimone100% (1)
- Margo - 3 NewДокумент71 страницаMargo - 3 NewKhushman KaurОценок пока нет
- Clinical Oncology For StudentsДокумент133 страницыClinical Oncology For StudentslillyanjdeoОценок пока нет
- Chapter 7 Neoplasia 1 2 Robbins and Cotran Pathologic Basis of Disease PDFДокумент9 страницChapter 7 Neoplasia 1 2 Robbins and Cotran Pathologic Basis of Disease PDFChethranОценок пока нет
- Cell Signaling - ImpДокумент11 страницCell Signaling - Impkrishnarajagopal2011Оценок пока нет
- Mucosal Immunity: Dono Indarto, DR., M.Biotech. ST., PHDДокумент34 страницыMucosal Immunity: Dono Indarto, DR., M.Biotech. ST., PHDsekar tyasОценок пока нет
- Gene-Based Vaccines and ImmunotherapeuticsДокумент5 страницGene-Based Vaccines and ImmunotherapeuticsMiguel Angel VegaОценок пока нет
- How To Give A Sensational Scientific TalkДокумент17 страницHow To Give A Sensational Scientific TalkMiguel Angel VegaОценок пока нет
- Plasmid DNA and Viral Vector-Based Vaccines For The Treatment of CancerДокумент11 страницPlasmid DNA and Viral Vector-Based Vaccines For The Treatment of CancerMiguel Angel VegaОценок пока нет
- Immune Surveillance of TumorsДокумент10 страницImmune Surveillance of TumorsMiguel Angel VegaОценок пока нет
- Novel Amyloid-Beta Specific Fragments From Human Antibody LibrariesДокумент24 страницыNovel Amyloid-Beta Specific Fragments From Human Antibody LibrariesMiguel Angel VegaОценок пока нет
- Phage-Displayed T-Cell Epitope Grafted Into Immunoglobulin Heavy-Chain Complementarity-Determining Regions: An Effective Vaccine Design Tested in Murine CysticercosisДокумент8 страницPhage-Displayed T-Cell Epitope Grafted Into Immunoglobulin Heavy-Chain Complementarity-Determining Regions: An Effective Vaccine Design Tested in Murine CysticercosisMiguel Angel VegaОценок пока нет
- AMYLOID-β PEPTIDE BINDS TO MICROTUBULE-ASSOCIATED PROTEIN 1B (MAP1B)Документ12 страницAMYLOID-β PEPTIDE BINDS TO MICROTUBULE-ASSOCIATED PROTEIN 1B (MAP1B)Miguel Angel VegaОценок пока нет
- Peptide Mimotopes of Mycobacterium Tuberculosis Carbohydrate ImmunodeterminantsДокумент7 страницPeptide Mimotopes of Mycobacterium Tuberculosis Carbohydrate ImmunodeterminantsMiguel Angel VegaОценок пока нет
- Towards A Taenia Solium Cysticercosis Vaccine: An Epitope Shared by Taenia Crassiceps and Taenia Solium Protects Mice Against Experimental Cysticercosis.Документ10 страницTowards A Taenia Solium Cysticercosis Vaccine: An Epitope Shared by Taenia Crassiceps and Taenia Solium Protects Mice Against Experimental Cysticercosis.Miguel Angel VegaОценок пока нет
- Vaccine Delivery Methods Using Viral VectorsДокумент15 страницVaccine Delivery Methods Using Viral VectorsMiguel Angel VegaОценок пока нет
- Bacteriophages As Tools For Vaccine and Drug DevelopmentДокумент3 страницыBacteriophages As Tools For Vaccine and Drug DevelopmentMiguel Angel VegaОценок пока нет
- Gene-Based Vaccines and ImmunotherapeuticsДокумент5 страницGene-Based Vaccines and ImmunotherapeuticsMiguel Angel VegaОценок пока нет
- Surface Mapping of Proteins Via Structure-Based Peptide Library Design.Документ7 страницSurface Mapping of Proteins Via Structure-Based Peptide Library Design.Miguel Angel VegaОценок пока нет
- Runt Related Proteins RUNXДокумент2 страницыRunt Related Proteins RUNXMiguel Angel VegaОценок пока нет
- Prognostic Implications of Cell Cycle Apoptosis and Angiogenesis BiomarkersДокумент13 страницPrognostic Implications of Cell Cycle Apoptosis and Angiogenesis BiomarkersMiguel Angel VegaОценок пока нет
- Tumour-Initiating Cells ChallengesДокумент18 страницTumour-Initiating Cells Challengessatya_chagantiОценок пока нет
- Mechanisms of Bone MetastasisДокумент10 страницMechanisms of Bone MetastasisMiguel Angel VegaОценок пока нет
- Theories of CarcinogenesisДокумент12 страницTheories of CarcinogenesisMiguel Angel VegaОценок пока нет
- Does Tumor Growth Follow A Universal LawДокумент14 страницDoes Tumor Growth Follow A Universal LawMiguel Angel VegaОценок пока нет
- Imaging of Ultraweak Spontaneous Photon Emission From Human Body Displaying Diurnal Rhythm (M Kobayashi)Документ4 страницыImaging of Ultraweak Spontaneous Photon Emission From Human Body Displaying Diurnal Rhythm (M Kobayashi)THE NIKOLA TESLA INSTITUTEОценок пока нет
- The Neurobiology of Decision - Kableglimcher09Документ13 страницThe Neurobiology of Decision - Kableglimcher09pancholarpancholarОценок пока нет
- Conversational American EnglishДокумент222 страницыConversational American EnglishDiana Vasiliniak96% (25)
- Insulin Resistance: Definition and Consequences: H.E. LebovitzДокумент14 страницInsulin Resistance: Definition and Consequences: H.E. LebovitzCarlos Andre Hernández EléspuruОценок пока нет
- 10 1021@acs Jmedchem 0c01195Документ25 страниц10 1021@acs Jmedchem 0c01195Khairunnisa Salsabila LutfiОценок пока нет
- Invitrogen 2012Документ251 страницаInvitrogen 2012Deepak Ranjan SahooОценок пока нет
- SITOKINДокумент16 страницSITOKINRidwan OorОценок пока нет
- Nino Gachechiladze ImunitetiДокумент268 страницNino Gachechiladze ImunitetiSaba GabadzeОценок пока нет
- Transcription Factor-Based Biosensors in Biotechnology - CurrentДокумент12 страницTranscription Factor-Based Biosensors in Biotechnology - CurrentConcetta SantiniОценок пока нет
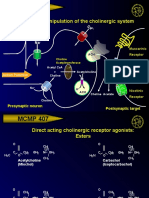
- MCMP 407: Pharmacologic Manipulation of The Cholinergic SystemДокумент13 страницMCMP 407: Pharmacologic Manipulation of The Cholinergic System16_dev5038Оценок пока нет
- Sy Compdiv1 Genes - t1Документ9 страницSy Compdiv1 Genes - t1Shreeya GanjiОценок пока нет
- Sitokin: I Wayan M. Santika, S.Farm., M.Si.,Apt Departement of Pharmacy, FMIPA, Udayana UniversityДокумент27 страницSitokin: I Wayan M. Santika, S.Farm., M.Si.,Apt Departement of Pharmacy, FMIPA, Udayana Universityweni diahОценок пока нет
- Topic: Cytokine and Their Roles in The Immune Regulation: Md. Jiyaul MustafaДокумент21 страницаTopic: Cytokine and Their Roles in The Immune Regulation: Md. Jiyaul MustafaAisha Siddiq100% (1)
- Compendium of Selected Recent Publications Cell and Molecular Biology ResearchДокумент16 страницCompendium of Selected Recent Publications Cell and Molecular Biology ResearchXyza Kim OlivaОценок пока нет
- 2 PharmacodynamicsДокумент51 страница2 PharmacodynamicsKriziaoumo P. Orpia100% (1)
- PG - ZOOLOGY-New CBCS Syllabus-NBU-2022-23Документ35 страницPG - ZOOLOGY-New CBCS Syllabus-NBU-2022-23Subhrangshu Ray Sarkar IX B 34Оценок пока нет
- Free Radicals and Antioxidants in Normal PhysiologicalДокумент41 страницаFree Radicals and Antioxidants in Normal PhysiologicalleosabreuОценок пока нет
- Biochem - Hormone and Signal TransductionДокумент36 страницBiochem - Hormone and Signal TransductionPrincess Jeyan PagatpatanОценок пока нет
- Apoptosis by Dr. Sanjiv Kumar-1Документ27 страницApoptosis by Dr. Sanjiv Kumar-1चौधरी हरिओम सौरोतОценок пока нет
- Targeting Tumor Associated Macrophages (TAMs) Via NanocarriersДокумент15 страницTargeting Tumor Associated Macrophages (TAMs) Via Nanocarriersyuvrajsingh3Оценок пока нет
- KrishPlex Assays FlyerДокумент2 страницыKrishPlex Assays Flyerkrishgen biosystemsОценок пока нет
- Targeting The IL-17 Pathway in Inflammatory Disease: Pierre MiossecДокумент1 страницаTargeting The IL-17 Pathway in Inflammatory Disease: Pierre Miossecanirbanmanna88320100% (1)
- Pathophysiology of Epilepsy: An Updated Review: International Journal of Medical and Health ResearchДокумент5 страницPathophysiology of Epilepsy: An Updated Review: International Journal of Medical and Health ResearchGayathiri GovindarajuОценок пока нет
- Kuliah Tumor Biology 2016Документ100 страницKuliah Tumor Biology 2016izulОценок пока нет
- Vol 6 No 3 Page 287 To 292Документ6 страницVol 6 No 3 Page 287 To 292ddemashОценок пока нет