Вам также может понравиться
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (895)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (344)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (121)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- BIO213 Fall 2016 SyllabusДокумент4 страницыBIO213 Fall 2016 SyllabusJosephParkОценок пока нет
- GFP TrapДокумент4 страницыGFP TrapAlleleBiotechОценок пока нет
- String DatabaseДокумент10 страницString DatabaseAnirudh PottipalliОценок пока нет
- A New Tool Based On Two Micromanipulators Facilitates The Handling of Ultrathin Cryosection RibbonsДокумент4 страницыA New Tool Based On Two Micromanipulators Facilitates The Handling of Ultrathin Cryosection Ribbonsrehu1976Оценок пока нет
- Inducible Expression of Claudin-1-Myc But Not occludin-VSV-G Results in Aberrant Tight Junction Strand Formation in MDCK CellsДокумент12 страницInducible Expression of Claudin-1-Myc But Not occludin-VSV-G Results in Aberrant Tight Junction Strand Formation in MDCK CellsKarina B Hernandez AОценок пока нет
- Tuesday, August 18, 2009Документ47 страницTuesday, August 18, 2009zorbaxОценок пока нет
- Corelative Fluorescence and Electron MicrosДокумент10 страницCorelative Fluorescence and Electron MicrosEmma StepinacОценок пока нет
- Conservation of BiodiversityДокумент19 страницConservation of BiodiversityAnkit AgarwalОценок пока нет
- XXXДокумент134 страницыXXXrhlzuraОценок пока нет
- Effective Microbial Bioremediation Via The Multi-Omics Approach: An Overview of Trends, Problems and ProspectsДокумент19 страницEffective Microbial Bioremediation Via The Multi-Omics Approach: An Overview of Trends, Problems and ProspectsUMYU Journal of Microbiology Research (UJMR)Оценок пока нет
- Bla Bla Bla New 2014Документ11 страницBla Bla Bla New 2014Georgiana GeoОценок пока нет
- Amino Acid StructureДокумент26 страницAmino Acid StructureSaumya SK100% (2)
- Neural TubeДокумент4 страницыNeural TubeRuth Danielle GasconОценок пока нет
- Obesity 1. Basic Science of ObesityДокумент10 страницObesity 1. Basic Science of ObesitygraceОценок пока нет
- As91603 Plant and Animal ResponsesДокумент2 страницыAs91603 Plant and Animal Responsesapi-323107386Оценок пока нет
- Ex3 Abo Forward Grouping Group7Документ19 страницEx3 Abo Forward Grouping Group7marielle roseОценок пока нет
- HTLA AntibodiesДокумент1 страницаHTLA Antibodieshakima.hassan.79Оценок пока нет
- Physiology and Biochemistry of Extremophiles-ASM Press (2007)Документ451 страницаPhysiology and Biochemistry of Extremophiles-ASM Press (2007)amine2263100% (1)
- Teaching Biology and Science in The FieldДокумент10 страницTeaching Biology and Science in The FieldIvan IvanovicОценок пока нет
- Muscle and Nervous Tissues LabДокумент4 страницыMuscle and Nervous Tissues LabSip BioОценок пока нет
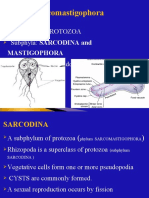
- Sarcomastigophora: A Phylum of PROTOZOA Subphyla: SARCODINA and Have Flagella or PseudopodiaДокумент20 страницSarcomastigophora: A Phylum of PROTOZOA Subphyla: SARCODINA and Have Flagella or Pseudopodiaabel semuОценок пока нет
- Biology ExamДокумент2 страницыBiology ExamZephieОценок пока нет
- Breast Cancer in The Views of Foods, Herbs & VitaminsДокумент66 страницBreast Cancer in The Views of Foods, Herbs & VitaminsKyle J. NortonОценок пока нет
- Cell MembranesДокумент34 страницыCell MembranesayfilonОценок пока нет
- Cell - Structure and Functions: World of CellsДокумент5 страницCell - Structure and Functions: World of CellsPriyanshu Priyadarshi MeherОценок пока нет
- Role of Free Radicals in Health and DiseaseДокумент49 страницRole of Free Radicals in Health and DiseaseKuzhandai VeluОценок пока нет
- Grade 5 Experiment No. 1 Animal and Plant ReproductionДокумент3 страницыGrade 5 Experiment No. 1 Animal and Plant ReproductionPaul James Abelardo TayagОценок пока нет
- Sbi4u1-Cell Bio-Cell Transport 4Документ15 страницSbi4u1-Cell Bio-Cell Transport 4alathena alathenaОценок пока нет
- PG BookДокумент5 страницPG BookUnnithan Rendezvous100% (2)
- Biodiversity of Flora and FaunaДокумент13 страницBiodiversity of Flora and FaunaLasmaenita SiahaanОценок пока нет