Вам также может понравиться
- The 2ND Training of Scientific Paper Writing of Biodiversitas, Journal of Biological DiversityДокумент106 страницThe 2ND Training of Scientific Paper Writing of Biodiversitas, Journal of Biological DiversityBiodiversitas, etcОценок пока нет
- Alternative Supplementary Biochemic Food For Growing Up The Fresh Water Lobster (Cherax Quadricarinatus)Документ8 страницAlternative Supplementary Biochemic Food For Growing Up The Fresh Water Lobster (Cherax Quadricarinatus)Biodiversitas, etcОценок пока нет
- Volume 11 - Number 2 - April 2010: Enetic Diversity Pecies DiversityДокумент3 страницыVolume 11 - Number 2 - April 2010: Enetic Diversity Pecies DiversityBiodiversitas, etcОценок пока нет
- Variation of Morphological and Protein Pattern of Cassava (ManihotДокумент9 страницVariation of Morphological and Protein Pattern of Cassava (ManihotBiodiversitas, etcОценок пока нет
- Domestica Val.)Документ6 страницDomestica Val.)Biodiversitas, etc100% (4)
- F 020202Документ5 страницF 020202Biodiversitas, etc100% (2)
- Jurnal CurcuminnnnnnДокумент6 страницJurnal CurcuminnnnnnKezia CandraОценок пока нет
- Oxysporum Schlecht F.sp. CubenseДокумент7 страницOxysporum Schlecht F.sp. CubenseBiodiversitas, etcОценок пока нет
- F 010201xxxДокумент5 страницF 010201xxxMuhammad Riv'at NalОценок пока нет
- Asam Giberelat-KhlorofilДокумент8 страницAsam Giberelat-KhlorofilPratama Irwin TalentaОценок пока нет
- Keanekaragaman Dan Potensi Jenis Tumbuhan Hutan Sekunder Di Kuala Ran, Kabupaten Bulungan, Kalimantan TimurДокумент7 страницKeanekaragaman Dan Potensi Jenis Tumbuhan Hutan Sekunder Di Kuala Ran, Kabupaten Bulungan, Kalimantan TimurBiodiversitas, etcОценок пока нет
- 197102211997022001bioscience 1Документ6 страниц197102211997022001bioscience 1Desti WibowoОценок пока нет
- 196607141999032001bioscience 4Документ5 страниц196607141999032001bioscience 4drunkpunchloveОценок пока нет
- Jurnal Nutrisi PDFДокумент5 страницJurnal Nutrisi PDFMuwahid IzhharОценок пока нет
- Effects of Physical Form of Barley Straw and Urea Supplementation On Intake and Digestion of Sheep Kept Under High Ambient TemperatureДокумент4 страницыEffects of Physical Form of Barley Straw and Urea Supplementation On Intake and Digestion of Sheep Kept Under High Ambient TemperatureBiodiversitas, etcОценок пока нет
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5795)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (895)
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (400)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (345)
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2259)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (266)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (121)
- Hormones NotesДокумент14 страницHormones NotesTeena SheikhОценок пока нет
- Fatigue AssignmentДокумент9 страницFatigue AssignmentRishbha TikuОценок пока нет
- 4 CYCLE FAT LOSS - Week 1 - Advanced Carb Depletion PDFДокумент17 страниц4 CYCLE FAT LOSS - Week 1 - Advanced Carb Depletion PDFDanCurtisОценок пока нет
- Caregiving TG - 10Документ31 страницаCaregiving TG - 10Amer Dalanda100% (2)
- Absorption & AssimilationДокумент16 страницAbsorption & AssimilationNaan Mahan AlleОценок пока нет
- Asnwer Scheme June 2015Документ11 страницAsnwer Scheme June 2015Nurul HaziqahОценок пока нет
- GI TrackДокумент125 страницGI TrackAndika HОценок пока нет
- Question Bank - New Questions: Chapter 18 HomeostasisДокумент5 страницQuestion Bank - New Questions: Chapter 18 HomeostasisAnders HuiОценок пока нет
- Creatine The Power Supplement LIVROДокумент295 страницCreatine The Power Supplement LIVROAndré Codea100% (2)
- GlucagonДокумент4 страницыGlucagonElenanana100% (1)
- Handbook of Meat ProcessingДокумент584 страницыHandbook of Meat ProcessingNguyễn Quang93% (15)
- QuizДокумент14 страницQuizmnouma1612Оценок пока нет
- Meeusen Et Al 2013 Overtraining Consensus ECSS ACSMДокумент20 страницMeeusen Et Al 2013 Overtraining Consensus ECSS ACSMBrandon HeatОценок пока нет
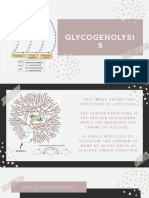
- GLYCOGENOLYSISДокумент12 страницGLYCOGENOLYSISDanah RosalesОценок пока нет
- Intensive Program - 2nd Group Session - 2020 - Batch 60Документ66 страницIntensive Program - 2nd Group Session - 2020 - Batch 60Tarun Kumar100% (1)
- Operation Bulk UpДокумент52 страницыOperation Bulk UpPhilip Choo100% (2)
- Chronic Adaptations To Training (Aerobic)Документ39 страницChronic Adaptations To Training (Aerobic)api-234153555100% (1)
- Pharmacy ScienceДокумент48 страницPharmacy ScienceMickey BrownОценок пока нет
- Gcse Biology Answers and Mark Schemes Questionsheet 1: Nervous & Hormonal Co-OrdinationДокумент9 страницGcse Biology Answers and Mark Schemes Questionsheet 1: Nervous & Hormonal Co-OrdinationKomalesh TheeranОценок пока нет
- Diet NutritionДокумент37 страницDiet Nutritionsowmyasrinivas25Оценок пока нет
- Glycogen Storage Disorders PDFДокумент4 страницыGlycogen Storage Disorders PDFAHMEDОценок пока нет
- Conditioning of AsilДокумент8 страницConditioning of AsilWorrenbuffet Rang60% (5)
- Organic Compounds in Living Things: PropertiesДокумент96 страницOrganic Compounds in Living Things: PropertiesTUĞRUL ERGÜLОценок пока нет
- Nutritional Needs of Critically Ill PatientsДокумент7 страницNutritional Needs of Critically Ill Patientstaner_soysurenОценок пока нет
- Carbohydrates ERTДокумент10 страницCarbohydrates ERTCharlotte WhiteОценок пока нет
- Lipid and Fat MetabolismДокумент25 страницLipid and Fat MetabolismCassy WalkerОценок пока нет
- Topic 10 - Homeostasis Notes o LevelДокумент6 страницTopic 10 - Homeostasis Notes o LeveldhanyaashinesОценок пока нет
- Jurnal ObesitasДокумент20 страницJurnal ObesitasTan Endy100% (1)
- Chapter 4 Carbohydrates QuizДокумент3 страницыChapter 4 Carbohydrates QuizAileen A. Monares100% (1)