Академический Документы
Профессиональный Документы
Культура Документы
Estructuras Reproduct
Загружено:
Isabel TrujilloИсходное описание:
Авторское право
Доступные форматы
Поделиться этим документом
Поделиться или встроить документ
Этот документ был вам полезен?
Это неприемлемый материал?
Пожаловаться на этот документАвторское право:
Доступные форматы
Estructuras Reproduct
Загружено:
Isabel TrujilloАвторское право:
Доступные форматы
Captulo 9 Estructuras reproductivas Las plantas terrestres presentan una gran variedad de estructuras reproductivas, las cuales pueden
ser muy sencillas, como esporas desnudas con limitada capacidad de dispersin, o muy complejas, como los frutos de las angiospermas. En todo caso, los rganos reproductores vegetales han permitido la exitosa dispersin de las plantas, ya que ellas han colonizado prcticamente la totalidad del planeta. En este captulo se revisa la estructura bsica de los rganos reproductores de las plantas terrestres, tanto en aspectos anatmicos como morfolgicos, desde las ms sencillas, las brifitas hasta las ms elaboradas, las angiospermas.
9.1. Generalidades acerca de las estructuras y ciclos reproductivos de las plantas terrestres y las algas verdes. Todas las plantas terrestres presentan un ciclo de vida con alternancia de dos generaciones, una productora de esporas denominada esporfita y otra productora de gametos o generacin gametfita (Figura 9.1). Estas generaciones siempre son interdependientes, ya que una de ellas es dominante y sostiene nutritivamente a la otra, a diferencia de las algas verdes, sus parientes ms cercanos, donde es posible que ambas generaciones sean de vida libre (ej: Ulva, Ulothrix). En las plantas sin sistema vascular (brifitas), la generacin dominante es la gametfita, mientras que en las plantas vasculares es la esporfita. Estas dos generaciones tambin difieren en su ploida, ya que el esporfito es diploide (2n) y el gametfito es siempre haploide (n). En contraste con las algas, el esporfito y el gametfito difieren en su apariencia externa, es decir, son heteromorfos, mientras que en las algas verdes es comn que las dos generaciones sean de similar apariencia o isomorfas, como en Ulva y Cladophora.
Figura 9.1. Esquema mostrando la alternancia de generaciones en el reino Viridiplantae. A) En las algas la generacin diploide es simplemente una clula, mientras que la etapa haploide es la dominante y puede ser mucho ms grande, B) en las brifitas el gametfito es fotosinttico y es la generacin dominante, C) en las pteridfitas ya el esporfito es fotosinttico y dominante, mientras que el gametfito es de corta duracin y pequeo, D) en las angiospermas se ha llegado al mximo estado de reduccin del gametfito, el cual est representado por el saco embrionario y el grano de polen y depende completamente de la generacin diploide o esporfito, que es la fase dominante y puede alcanzar proporciones gigantescas. Los rganos reproductores de las plantas terrestres estn construidos con un mismo patrn bsico, en el cual hay un tejido interno frtil y una cubierta externa de clulas estriles, que puede tener una o varias capas de grosor. Slo dentro del grupo de las algas del orden Charales hay una estructura protectora estril similar, por lo que esta cubierta protectora debe considerarse una importante innovacin evolutiva. Los rganos reproductivos de las plantas son los esporangios y los gametangios (Figura 9.2 y portada del captulo). Los primeros producen esporas, por lo que son propios de la generacin esporfita, mientras que los segundos son productores de gametos y estn presentes en el gametfito. La diferencia entre los esporfitos de las plantas se aprecia en la portada, donde un sencillo esporangio de una brifita es comparado con un complicado arreglo de esporangios y estructuras anexas en una angiosperma.
Figura 9.2. Esquema de los rganos reproductivos en las plantas inferiores y ciclo de vida de un alga verde. A) Cubierta estril de los gametangios en cuyo interior se desarrolla una sola ovoclula grande en los arquegonios y muchas clulas espermticas pequeas en los anteridios, B) parte superior del esporangio de un musgo en el que se muestra la capucha protectora (b1) y el tejido esporgeno interno (b2), rodeado de una cubierta estril. D) Ciclo de vida de un alga verde isomorfa: 1) cigoto, 2) esporfito, 3), esporas flageladas haploides que germinan originando los gametfitos, 4) gametos flagelados haploides que se fusionan y dan origen a un nuevo cigoto diploide. En muchas especies el esporfito y el gametfito son iguales morfolgicamente, por lo que se dice que son isomorfas. Las esporas y los gametos tambin son similares pero se diferencian funcionalmente, ya que los gametos se fusionan entre s para dar origen a una nueva clula, mientras que las esporas germinan directamente y producen un individuo. Dado que en las plantas hay alternancia de generaciones, existen dos tipos de clulas reproductivas, el gameto y la espora, los cuales pueden ser similares morfolgicamente pero con una gran diferencia funcional. En efecto, el primero debe fusionarse con otro gameto para generar un nuevo individuo, mientras que la espora no se une a otra clula similar, sino que germina para dar lugar a un nuevo individuo directamente. Es muy importante tener en cuenta que en las plantas terrestres las esporas nunca tienen flagelos, a diferencia de las algas cuyas esporas pueden ser flageladas. Adems, las esporas de las plantas son haploides porque son producidas por meiosis a partir de clulas esporgenas diploides. En cuanto a los gametos vegetales, stos tambin son haploides, pero se producen por mitosis ya que la generacin gametfita es completamente haploide. Los gametos de las plantas terrestres pueden ser flagelados en algunos grupos. Otro aspecto en el que es importante contrastar la biologa reproductiva de las plantas terrestres con las algas verdes, es en la presencia del embrin. ste es el cuerpo primordial o rudimento del esporfito, que se forma a partir del cigoto diploide y es retenido temporalmente en los tejidos maternos, el cual puede entrar en estado de latencia antes de proseguir su desarrollo. En las algas el cigoto no pasa por estado embrionario, sino que empieza a multiplicarse produciendo directamente el cuerpo del organismo adulto en su forma definitiva, bien sea filamentoso o formando un talo. Por ello varios autores se refieren a las plantas terrestres con el apelativo de embrifitas, para enfatizar esta notable diferencia con las algas verdes. Para finalizar este apartado introductorio, es necesario hacer una aclaracin en cuanto a la terminologa referente al gametfito y al esporfito. Usualmente se emplean los prefijos micro y mega para designar las estructuras masculinas y femeninas, como por ejemplo
microgametfito, que es el organismo productor de gametos masculinos, o megagametfito, que es el individuo productor de gametos femeninos. Sin embargo, estos prefijos tambin se usan en la generacin esporfita, ya que en ella se habla de microsporas y megasporas, pero es necesario aclarar que en ese caso no hacen referencia a la sexualidad, dado que slo los individuos que producen gametos tienen sexo, bien sea masculino o femenino. El esporfito, por definicin, es un individuo productor de esporas, no de gametos, por lo tanto no puede tener ni clulas ni rganos reproductivos masculinos o femeninos. Al usar las palabras megaspora y microspora se hace referencia al tamao de las esporas y al hecho que las primeras darn origen a los megagametfitos, estos s de sexo femenino ya que producirn ovoclulas, mientras que las microsporas originarn los microgametfitos, de sexo masculino.
Figura 9.3. Gametfito femenino de Marchantia sp. En esta foto se aprecia claramente la presencia de los arquegoniforos, con sus caractersticas estructuras digitiformes en la parte superior. 9.2 Las plantas terrestres sin sistema vascular poseen las estructuras reproductivas ms sencillas del reino vegetal. Las plantas que carecen de un sistema vascular (avasculares), son conocidas en conjunto como brifitas y muestran total dependencia del agua lquida para completar su ciclo reproductivo, a pesar de ser organismos terrestres. Su ciclo de vida consta de generaciones heteromorfas que se alternan, siendo la generacin gametfita la dominante. Los rganos reproductores tanto de la generacin esporfita como de la gametfita, son bsicamente un conjunto de clulas con potencial reproductivo, envueltas en unas pocas capas de tejidos estriles cuya funcin es proteger a las clulas reproductivas en formacin. En la generacin gametfita se puede formar gametangios de ambos sexos en el mismo individuo (especies monoicas) o en individuos diferentes (especies dioicas), los cuales producen gametos masculinos mviles o gametos femeninos inmviles. Los gametos masculinos nadan hacia los femeninos y, luego de la fertilizacin, se forma un cigoto que es retenido dentro del arquegonio y alimentado por los tejidos maternos, fenmeno al que se denomina matrotrofia. No hay conexiones de tipo plasmodesmo entre el embrin en desarrollo y el tejido materno, lo que implica que el transporte de nutrientes y otras sustancias se realiza slo por la va apoplstica. En la interfase de las generaciones se ubican clulas de transferencia que forman una placenta, las cuales facilitan la movilizacin de nutrientes hacia el embrin, debido a que poseen numerosas invaginaciones en su pared que aumentan la superficie de transporte. El embrin se desarrolla produciendo un nuevo individuo de la generacin esporfita, el cual al
madurar producir esporangios en los que se generar las esporas por meiosis. Cuando las esporas estn maduras sern liberadas y germinan dando origen a los gametfitos, con lo que se completa el ciclo reproductivo.
Figura 9.4. Corte transversal del gametfito de una heptica (Marchantia sp.). A) Las estructuras redondeadas en la parte inferior de la capucha son arquegonios, B) cuerpo del arquegoniforo, C) rizoides, hebras de una clula de grosor en la parte inferior del talo que lo anclan al sustrato. 9.2.1. El gametangio es el rgano encargado de producir gametos. El cuerpo de la generacin gametfita se conoce como gametforo ya que se encarga de producir y sostener los gametangios (Figura 9.2A), que son los rganos generadores de las clulas reproductoras gamticas: clulas espermticas y ovoclulas. Las clulas espermticas son pequeas, biflageladas y se producen en el microgametangio, tambin llamado anteridio, mientras que la ovoclula o gameto femenino, es grande e inmvil y se encuentra dentro del megagametangio o arquegonio. A continuacin se estudia el gametfito en los tres diferentes grupos de brifitas, ya que hay diferencias importantes entre ellas. En los musgos el anteridio posee un corto tallo que lo sostiene y un cuerpo ovalado con una masa interna de clulas espermatgenas cubiertas por una capa de clulas estriles. Las clulas espermticas son producidas por mitosis, ya que el gametfito se origina de una espora haploide. El arquegonio posee una base ensanchada y un cuello largo y esbelto que es hueco en la madurez, slo una ovoclula sobrevive en cada arquegonio, la cual se deposita en la base del rgano. El anteridio y el arquegonio pueden coexistir en el mismo individuo en especies bisexuales o monoicas (Funaria, Pottia), mientras que estn en individuos separados en especies dioicas (Barbula, Polytrichum, Rhacomitrium). Tanto los anteridios como los arquegonios pueden presentarse agrupados en estructuras llamadas cabezuelas (anteridial y arquegonial), en las cuales hay estructuras estriles llamadas parfisis, que se ubican intermezcladas con los gametangios. La apertura del anteridio se realiza bien sea por acumulacin de lquido en el interior, lo que genera una presin y empuja las clulas espermticas hacia fuera, o por contraccin de las clulas de la cubierta protectora estril. La clula espermtica puede ser dispersada por gotas de lluvia, pero siempre tiene que viajar nadando hasta el arquegonio. Se sabe de la secrecin de sacarosa por parte del arquegonio como gua qumica para las clulas espermticas.
Figura 9.5. Esporangios en un musgo. La parte foliosa fotosinttica inferior es el gametfito, mientras que los tallos con unas cpsulas en sus extremos representan la generacin esporfita. En las hepticas la generacin gametfita puede ser monoica o dioica y sus gametangios se ubican intermezclados con hojas vegetativas, o en ramas laterales rodeados por hojas modificadas. Los gametforos ms complejos de las hepticas estn presentes en Marchantia, donde existen anteridiforos y arquegoniforos, rganos portadores de anteridios y arquegonios, respectivamente (Figuras 9.3 y 9.4). El anteridiforo consta de un tallo que se eleva sobre el gametfito y que sostiene una estructura aplanada dentro de la cual estn inmersos los anteridios. El arquegoniforo es palmiforme, con un tallo largo que sostiene una estructura plana con proyecciones apendiformes de las cuales se sostienen a los arquegonios en su superficie abaxial. Los antoceros, el grupo menos estudiado de las brifitas, presentan notables diferencias en los rganos reproductivos con el resto de ellas. El gametfito es aplanado y casi siempre unisexual, con los gametangios ubicados en la parte superior del cuerpo. El anteridio se origina internamente de una clula anteridial inicial y queda recluido en una cavidad, cuyo techo eventualmente se romper quedando expuesto al ambiente y liberando las clulas espermticas al ambiente. Cuando esto sucede las clulas de la capa estril externa cambian de color, bien sea a amarillo o naranja. Los arquegonios son superficiales, a veces dispuestos en filas, pero no rodean completamente a la ovoclula por lo que no son rganos discretos. Ellos estn en contacto directo con las clulas vegetativas del talo en el que estn embebidos. 9.2.2. Las esporas son producidas en el esporangio por meiosis. La generacin esporfita en las plantas avasculares est constituida por un individuo diploide, el cual depende completamente del gametfito para sobrevivir, excepto en los antoceros. El esporfito produce esporas mediante meiosis en un rgano especializado llamado esporangio (Figura 9.5), siendo los musgos el grupo de brifitas en los que ms se ha estudiado. En ellos, el cigoto diploide, que es la primera clula de la generacin esporfita, se divide transversalmente por mitosis generando dos hijas, de las cuales la superior da origen a un nico esporangio apical llamado cpsula. Esta estructura posee una columna interna de tejido estril denominada columela y una cubierta protectora tambin estril, entre las cuales se desarrolla el tejido esporgeno (Figura 9.2B). Las esporas se producen por meiosis y son liberadas slo cuando la tapa protectora de la cpsula, llamada oprculo, se separa del resto del esporangio. Esta separacin deja expuestos los dientes del peristoma, estructuras sensibles a la humedad del aire que pueden doblarse para liberar las esporas que sern dispersadas por el viento. Los dientes del peristoma se disponen en dos capas, una ms interna que la otra. Si las esporas caen en un medio adecuado, germinan produciendo la generacin gametfita.
Figura 9.6. Corte transversal del esporangio de una heptica (Marchantia sp.). A) Pared del esporangio, B) elteres, filamentos que ayudan a la dispersin de las esporas. En las hepticas los esporangios poseen una estructura un poco diferente (Figura 9.6). Por ejemplo, carecen de columela y de oprculo, por lo que la liberacin de las esporas se realiza por dehiscencia del esporangio, la cual se da a lo largo de cuatro suturas longitudinales. En algunas especies de Riccia y Ricciocarpus, la liberacin se da por la muerte y descomposicin del esporangio. Existe adems un grupo de clulas estriles denominadas elteres, las cuales son elongadas, con engrosamientos en la pared interna, que pueden adquirir la forma de resorte para estirarse y encogerse con el objeto de dispersar las esporas, dependiendo de la humedad del ambiente. El esporfito de los antoceros es quiz el ms diferente entre todas las brifitas. Una vez fecunda la ovoclula, sta se divide longitudinalmente y empieza a formarse el cuerpo del esporfito, en cuyo eje central se generar el tejido esporgeno que dar origen a las esporas. La dehiscencia del esporangio se realiza por dos aberturas lineales produciendo la liberacin de las esporas, que pueden ser multicelulares en algunas especies. Los antoceros poseen columela como en los musgos, adems de alteres, pero stos son pluricelulares y sus paredes carecen de engrosamientos espiralados.
Figura 9.7. Cuerpo vegetativo y estructuras reproductivas en Psilotum. A) Espcimen de herbario mostrando el cuerpo dictomo y la posicin de los sinangios, B) ampliacin de los sinangios que muestra su naturaleza trmera, C) representacin esquemtica de un corte transversal de uno de ellos, en la que se aprecia el tejido esporgeno en su interior, distribuido en tres grupos discretos. 9.2.3. Las brifitas tambin pueden reproducirse vegetativamente. En las brifitas es comn encontrar especies que puedan reproducirse asexualmente por fragmentacin, pues pequeos trozos del cuerpo son capaces de regenerar un individuo nuevo. De hecho, para algunas especies se cree que esta es la principal forma de reproduccin. Sin embargo, adems de la fragmentacin tambin hay estructuras multicelulares especializadas en la propagacin asexual de musgos y hepticas, las cuales se denominan yemas (gemmae en latn). Estas estructuras son particularmente conspicuas en hepticas del gnero Marchantia, donde se localizan en la superficie dorsal del gametfito sobre estructuras en forma de cpula denominadas conceptculos y de donde son dispersadas por gotas de lluvia. Tambin han sido llamadas propgulos. 9.3. Las plantas vasculares sin semilla presentan estructuras reproductivas ms complejas que las brifitas. Las plantas vasculares que no producen semillas se conocen en conjunto como pteridfitas, e incluyen a los helechos verdaderos (Pteridophyta), las colas de caballo (Equisetophyta), los licopodios (Lycopodiophyta) y las psilotfitas o helechos primitivos (Psilotophyta). No es un grupo monofiltico, pero se les estudia siempre en conjunto atendiendo propsitos didcticos, ya que se diferencian de las brifitas por poseer verdaderos tejidos conductores, mientras que del resto de plantas vasculares se distinguen porque carecen de semilla. Actualmente hay evidencia fuerte que anida las colas de caballo dentro de los helechos, lo cual se aborda con ms detalle en la seccin de diversidad. En estas y en todas las dems plantas terrestres, el esporfito es la generacin dominante y es comnmente perenne, en contraste con las brifitas. Las pteridfitas presentan la caracterstica de agrupar sus esporangios en estructuras especializadas, siendo esto lo que ms las diferencia de las brifitas, en trminos reproductivos. Las agrupaciones de esporangios se denominan soros en los helechos, mientras que en las psilotfitas se llaman sinangios y en las colas de caballo y licopodios reciben el nombre genrico de estrbilos.
Figura 9.8. Espcimen de herbario de Huperzia. A) Parte vegetativa basal, B) estructura reproductiva apical, en la cual los esporangios estn subtendidos por hojas modificadas (esporfilos). El estrbilo es un rgano generalmente terminal, que consta de un eje central en el que se insertan hojas o pequeos tallos que sostienen a los esporangios, mientras que el sinangio es una agrupacin de tres esporangios que estn fusionados y que son sostenidos por un pequeo tallo. De otra parte, los soros son agrupaciones de esporangios individuales unidos a un mismo punto en el envs de la fronda de los helechos y que pueden presentar o no una estructura protectora. Los gametfitos producidos por la germinacin de las esporas que se producen en las estructuras recientemente mencionadas, poseen gametangios bsicamente iguales a los de las brifitas, ya que el anteridio es casi esfrico, multicelular y cubierto por una capa de tejido estril, mientras que el arquegonio es alargado con la base ensanchada y produce una sola ovoclula. 9.3.1. En las psilotfitas los esporangios se agrupan en rganos ms especializados que en las brifitas. Los esporangios de la divisin Psilotophyta estn fusionados en una estructura denominada sinangio, en la cual se lleva a cabo la meiosis que producir las esporas (Figura 9.7). Los sinangios se originaron por la fusin de tres esporangios y no son sostenidos por hojas modificadas, sino por ramas muy cortas. Cuando las esporas maduran, su liberacin se da por la apertura del sinangio a travs de suturas longitudinales. Una vez liberadas, las esporas germinan dando origen al gametfito, que es monoico y en el que sus anteridios sobresalen de la superficie del cuerpo, mientras que los arquegonios estn parcialmente hundidos. La clula espermtica es flagelada ya que debe nadar para alcanzar la ovoclula. Tanto en sus estructuras reproductoras como vegetativas, las psilotfitas son las plantas vasculares ms sencillas, muy parecidas a los fsiles de las primeras plantas vasculares, pero se cree que llegaron a ese estado mediante una aparente reduccin secundaria de sus rganos.
9.3.2. Los licopodios presentan hojas especializadas que acompaan a los rganos reproductivos. En este grupo de plantas vasculares, agrupadas en la divisin Lycopodiophyta, hay hojas reproductivas especializadas que sostienen los esporangios y se denominan esporfilos (Figuras 9.8 y 9.9). Pueden estar agrupadas al final de los tallos formando estrbilos y en este caso no son fotosintticas, o pueden estar entre las hojas vegetativas y se distinguen de ellas slo por la presencia del esporangio en su axila. Algunas plantas de este grupo presentan una innovacin importante en sus esporas con respecto a los helechos primitivos y las brifitas, ya que presentan dos tipos de ellas segn su morfologa: las microsporas que darn origen al gametfito masculino y las megasporas que originarn los gametfitos femeninos. A esta condicin se le conoce como heterosporia. Las hojas que las subtienden se denominan microsporfilos y megasporfilos respectivamente, las cuales pueden estar en el mismo estrbilo. El megasporangio produce cuatro megasporas, mientras que en el microsporangio se producen muchas microsporas.
Figura 9.9. Corte longitudinal de un estrbilo de un licopodio hetersporo (Selaginella sp.). A) Esporfilo, hoja modificada que subtiende al esporangio, B) macroesporangio, C) tallo del estrbilo, D) microsporangio. Ambos gametfitos de este grupo crecen dentro de las esporas y stas se abren slo cuando el masculino est listo para liberar las clulas espermticas flageladas y el femenino est receptivo. El cido ctrico parece jugar un papel importante en la atraccin de la clula espermtica por parte del arquegonio y se sabe que la autofertilizacin est poco favorecida. Tras la fecundacin el cigoto se divide y una de las clulas hijas producir el suspensor, otra innovacin de este grupo con respecto a las plantas ms simples. La funcin de este rgano es empujar el embrin dentro del tejido nutricio del gametfito que lo rodea, aunque tambin parece estar involucrado en la transferencia de nutrientes hacia el embrin. La otra clula hija de la primera divisin mittica del cigoto dar origen al embrin, que se desarrollar en el esporfito propiamente dicho.
Figura 9.10. Estrbilo de un equiseto o cola de caballo (Equisetum sp.). A) Tallo articulado del esporfito, B) estrbilo terminal con esporangiforos (puntos negros), uno de los cuales est ampliado a la derecha, C) tallo central del estrbilo, D) pednculo del esporangiforo, E) tejido esporgeno dentro de los esporangios. 9.3.3. Las colas de caballo presentan una innovacin interesante en la disposicin de sus esporangios. Las colas de caballo se agrupan en la divisin Equisetophyta (Figura 9.10), taxn que tiene la caracterstica de agrupar los esporangios en una estructura en forma de sombrilla llamada esporangiforo, la cual posee un tallo corto y una superficie aplanada superior de la que se sostienen entre 5 y 10 esporangios. Los esporangiforos se disponen en espiral en un estrbilo y slo producen un tipo de espora, por lo que se dice que estas plantas son homsporas. Segn la especie, el estrbilo puede estar en el pice de un tallo fotosinttico o de un tallo especial reproductor, carente de clorofila. Las esporas germinan produciendo un gametfito masculino o bisexual. Del anteridio se liberan numerosos espermatozoides multiflagelados que nadan hasta el arquegonio para fecundar la ovoclula. 9.3.4. Los helechos se caracterizan por la presencia de soros abaxiales en el esporfito. El esporfito de algunas especies de helechos posee hojas especializadas en la produccin de esporangios, denominadas esporfilos, las cuales son alargadas como tallos, no son fotosintticas y poseen cerca de su pice muchos esporangios agrupados, como en los gneros Blechnum y Osmunda. En la mayora de las especies de helechos, sin embargo, todas las hojas son fotosintticas y poseen agrupaciones de esporangios denominadas soros, que se ubican en la superficie inferior de las hojas (Figura 9.11). En los esporangios, los cuales constan de un tallo corto que sostiene una estructura globosa con un borde engrosado, son producidas las esporas por meiosis. Los soros pueden estar protegidos por una estructura con forma de sombrilla denominada indusio o pueden estar completamente expuestos al aire. Se pueden ubicar tanto en el centro de la lmina como en sus bordes, denominndose en este caso soros marginales. Cuando el soro es marginal puede estar envuelto por un pliegue del borde de la hoja, el cual imita al indusio y recibe el nombre de pseudoindusio (falso indusio, figura 9.12).
Figura 9.11. Diferentes tipos de soros en los helechos. A) Esporangios en frondas frtiles de Anemia sp., B) soros redondeados en Polypodium sp., C) soros marginales continuos en la superficie inferior de una fronda de un helecho terrestre y D) soros marginales discretos en fronda de Adiantum sp.
Figura 9.12. Detalle de un corte transversal de un soro marginal. A) Epidermis de la fronda, B) esporangios, C) pseudoindusio, estructura que cubre el conjunto de esporangios (soro). La mayora de los helechos son homsporos, es decir, poseen esporas morfolgicamente iguales, pero hay dos grupos de helechos muy particulares que son hetersporos, los rdenes Marsileales y Salvineales, ambos acuticos. En todo caso, las esporas son liberadas tras la apertura del esporangio luego de haber madurado, para as germinar y producir diminutos gametfitos aplanados, en los que se produce arquegonios y anteridios en la superficie inferior,
entre los rizoides unicelulares (Figura 9.13). Se sabe que los gametfitos producen sustancias hormonales como el anteridigeno, que induce la maduracin de los anteridios de otros gametfitos. Cuando las clulas espermticas estn maduras, son liberadas y nadan mediante flagelos hacia el arquegonio para fecundar la ovoclula. Al parecer hay sustancias qumicas que guan a los espermatozoides hacia su destino, al cual pueden llegar slo si existe agua lquida disponible.
Figura 9.13. Gametfito de un helecho. A) Cuerpo aplanado, a menudo en forma de corazn y de pocos milmetros de dimetro, B) gametangios, estructuras productoras de gametos, C) rizoides, filamentos usados para adherirse al sustrato. 9.4. Estructuras reproductivas de las plantas vasculares con semilla desprotegida (gimnospermas). La innovacin reproductora ms notable en las plantas terrestres es la semilla (del latn sperma), razn por la cual a las plantas que la poseen se les denomina espermatfitas. Estas plantas tambin tienen en comn que presentan la mayor reduccin de tamao del gametfito en todo el reino vegetal. La semilla puede estar cubierta por un carpelo, como en las angiospermas, o completamente expuesta al ambiente (sin proteccin o desnuda), como sucede en las gimnospermas. Las plantas con semilla descubierta constituyen un grupo variado cuya monofilia no ha sido demostrada, pero tradicionalmente se han estudiado en conjunto debido al factor comn de producir sus semillas sin la proteccin del mencionado carpelo. Entre las especies vivientes de gimnospermas hay cuatro grupos a los que se les ha dado categora de divisin, que son las conferas (Pinophyta), las ccadas (Cycadophyta), las gnetfitas (Gnetophyta) y el grupo monotpico de las ginkgfitas (Gingkgophyta: Ginkgo biloba). 9.4.1. Las conferas poseen los estrbilos ms grandes del reino vegetal y un complicado ciclo reproductivo. El esporfito de las conferas es un rbol de gran tamao que posee dos tipos de rganos productores de esporas, a los que se denomina estrbilos o conos, que son de mucho mayor tamao que los presentes en brifitas y pteridfitas. Como todas las plantas con semilla son hetersporas, hay conos productores de polen y conos productores de semillas (Figura 9.14). Los conos productores de polen estn generalmente hacia los extremos de las ramas, a menudo en la parte baja del rbol y son de menor tamao que los conos de semillas. El cono de polen consta de un eje central con vascularizacin en el cual se desarrollan los esporfilos, hojas
pequeas que sostienen a los microsporangios. Dentro de los microsporangios las clulas madres de las microsporas producen por meiosis ttradas de microsporas, cada una de las cuales desarrollar un grano de polen o gametfito masculino. Hay dos tipos de conos con semillas en las conferas, el de las coniferales y el de las taxales. Los conos de semillas en las coniferales son compuestos (Figura 9.14), pues hay yemas axilares a diferencia de los conos de polen. En dichos conos se producen brcteas, que son hojas modificadas que sostienen su respectiva yema axilar, la cual produce la escama ovulfera. Sobre la escama se ubican dos vulos que consisten de un megasporangio, tambin llamado nucela, rodeado por integumentos que se abren hacia el eje del cono por un micrpilo. En cada megasporangio hay un megasporocito o clula madre de la megaspora, que se divide por meiosis y produce cuatro esporas de las cuales slo sobrevive una, la ms alejada del micrpilo. La megaspora haploide que sobrevive no es liberada, sino que se desarrollar dentro del megasporangio y producir el gametfito femenino, en cuyo interior se pueden desarrollar dos arquegonios, cada uno de los cuales contendr una ovoclula. De otra parte, en las taxales no hay conos femeninos ya que las semillas se ubican en tallos cortos laterales en las ramas, mientras que los microsporangios se desarrollan en grupos de hasta nueve sobre microsporfilos aplanados. Sus semillas desarrollan un arilo, estructura brillante y carnosa que parece atraer animales dispersores.
Figura 9.14. Vista macroscpica y anatmica de estrbilos de gimnospermas. A) Cono de semillas de una gimnosperma ornamental introducida a Colombia (Cryptomeria japonica), B) cono de polen, ms pequeo y totalmente independiente del cono de semillas, ya que todas las plantas con semilla son hetersporas (producen dos tipos de esporas en rganos separados en el mismo individuo). A la derecha hay un detalle del corte longitudinal de un cono de semillas en el ciprs (Cupressus sp.), en el que se muestra el megasporocito en desarrollo (C) y la escama ovulfera (D). La generacin gametfita en las conferas es muy pequea y depende completamente del esporfito. Est constituida por el grano de polen (Figura 9.15), tambin llamado gametfito masculino o microgametfito y el megagametfito o gametfito femenino. El grano de polen posee cuatro clulas: dos protlicas (aparentes remanentes del talo del gametfito que se desintegran antes de la fertilizacin), una clula del tubo y una clula generativa, la cual se multiplica en dos produciendo una clula tallo (aparente vestigio del anteridio) y una clula del cuerpo, que es la que originar las clulas espermticas. La dispersin del polen se da luego de la dehiscencia del microsporfilo y es debida al viento (anemfila). Este mtodo requiere la produccin de una gran cantidad de granos ya que muchos de ellos caen fuera de los conos femeninos y son desperdiciados. Inicialmente, el gametfito femenino es cenoctico y puede contener ms de 7000 ncleos. La formacin de paredes celulares puede durar hasta un ao y
dar lugar a un megagametfito celular, en el que se desarrollan dos o tres arquegonios con una ovoclula cada uno, la cual est llena de sustancias nutritivas como glcidos y protenas. Las conferas generalmente crecen formando grandes agregaciones de individuos, lo que es necesario si dependen de la polinizacin por el viento, ya que de otra forma, con individuos aislados, las probabilidades de fecundacin seran mnimas. El polen llega al cono femenino antes que el huevo est maduro y alcanza el vulo debido a que las escamas ovulferas estn ligeramente separadas entre s. Luego se cierran haciendo del cono una estructura ms compacta y puede pasar hasta un ao para que se verifique la fertilizacin. La forma del grano de polen es redondeada, con dos prolongaciones laterales que sirven para la dispersin anemfila. La germinacin del polen produce un tubo polnico que debe digerir los tejidos para llegar a la ovoclula, ya que el megasporangio no se abre. Es posible que se fertilice a los dos o tres gametos femeninos presentes en sus respectivos arquegonios, produciendo el fenmeno de poliembriona, pero generalmente slo uno es viable. De las primeras divisiones del cigoto se origina el suspensor, que empuja el grupo menor de clulas que originarn el embrin dentro del tejido del megagametfito. Las clulas empujadas reciben el nombre de proembrin y generarn el embrin verdadero. El gametfito femenino contina creciendo y proveer el tejido nutritivo para el embrin, el cual consta de una radcula, el hipoctilo, el epictilo y los cotiledones, que pueden ser muchos. La semilla es retenida en el cono, el cual generalmente es seco pero puede ser carnoso en algunas especies de Podocarpus y Juniperus, hasta cuando est madura y lista para la dispersin.
Figura 9.15. Microgametfito o gametfito masculino o grano de polen de una gimnosperma (Pinus sp.). El polen es producido en los microsporangios y posee una caracterstica estructura alada, la cual es til para la dispersin por el viento. 9.4.2. Las ccadas son gimnospermas con sexos separados. Todas las ccadas actuales son dioicas. Sus rganos reproductores son conos situados en el pice de la planta o cerca de l y pueden ser de polen o de semillas (Figura 9.16). Los conos de polen pueden alcanzar ms de 30 cm de longitud, poseen microsporfilos arreglados en espiral con grupos de microsporangios en los cuales se generan los granos de polen. Una vez germinado, el polen produce un tubo polnico que puede ser ramificado pero que no penetra el arquegonio. A lo largo del tubo polnico se mueven las dos clulas espermticas multiflageladas en busca de la ovoclula para fecundarla. De otra parte, los conos de semilla son ms variables en su morfologa, siendo algunos el resultado de la agregacin de megasporfilos foliosos, unidos de manera poco compacta, con 6 a 8 vulos basales. Otras especies poseen conos ms compactos,
con megasporfilos en forma de escudo y con slo dos vulos, a los cuales se les considera los conos ms derivados.
Figura 9.16. Cono de una ccada. Los conos de las ccadas se dan en la parte terminal de la planta, entre las hojas que los rodean a manera de corona. Cuando maduran, los conos de semillas como el de esta figura, liberan pasivamente los propgalos que darn origen a la siguiente generacin. 9.4.3. Las gnetfitas poseen caractersticas reproductivas de difcil interpretacin. Este grupo de plantas comprende los gneros Ephedra, Gnetum y Welwitschia (Figura 9.17), los cuales poseen estrbilos compuestos. Los conos de polen de las gnetfitas poseen pequeas brcteas y el polen es alargado con bordes angostos, sin flagelos. Existe un verdadero tubo polnico que conduce las clulas espermticas hasta la cercana de las ovoclulas. Los conos de semillas poseen tejido accesorio alrededor de los vulos y la interpretacin de esta caracterstica es compleja. Gnetum y Welwitschia carecen de arquegonios y la clula generativa del polen produce los espermatozoides directamente, al igual que las angiospermas, con la particularidad que Welwitschia produce un crecimiento tubular a partir de sus vulos, de forma similar a como lo hace el tubo polnico. De otra parte, Ephedra y Gnetum presentan doble fecundacin, al igual que las angiospermas, pero se producen dos embriones, en lugar de un embrin y un endospermo como sucede en las plantas con flores. Por todas estas similitudes reproductivas repartidas entre los miembros del grupo, hay varias hiptesis acerca de cual de ellos est ms cercanamente emparentado con las angiospermas, as que hay bastante trabajo por hacer antes de clarificar este asunto.
Figura 9.17. Estructuras reproductivas en Welwitschia, gnero que posee una sola especie dioica. Conos de semillas a la izquierda y conos de polen a la derecha. Todas las gnetfitas (Ephedra, Gnetum y Welwitschia), presentan el fenmeno de doble fecundacin. Foto cortesa de M OBrien.
Figura 9.18. Semillas de ginkgo. Esta especie es una gimnosperma dioica, as que las semillas aqu mostradas slo se hallan en los rboles que producen vulos y carecen de envoltura protectora externa. 9.4.4. rganos reproductivos del Ginkgo biloba. El ginkgo, la nica especie viviente de las ginkgfitas, es dioica, sin conos, con vulos desnudos (Figura 9.18), situados en pares al final de un eje corto llamado braquiblasto, los cuales producen compuestos de azufre y desarrollan una semilla con una cubierta de tres capas. El ftido olor que producen los vulos que caen por no ser fecundados hace que slo rboles machos, es decir, que produzcan polen, sean usados con fines ornamentales. El polen es producido en un eje de las ramas cortas que posee microsporfilos y algunos esporangiforos, los cuales portan un par de esporangios cada uno. Tanto los microsporfilos como los megasporfilos son sostenidos en sus respectivos individuos en ramas cortas, intermezclados con las hojas. Una vez el grano de polen ha llegado al vulo, desarrolla un tubo polnico
ramificado que se torna haustorial y crece entre las clulas de la nucela, sin penetrar el arquegonio. Luego, la parte basal del polen se agranda y libera dos espermatozoides que nadan hasta alcanzar el gametfito femenino para fecundar las ovoclulas. 9.5. La mxima especializacin reproductiva se da en las plantas vasculares con semilla protegida, mejor conocidas como angiospermas. Las angiospermas obtienen su nombre de dos palabras griegas que significan vasija o recipiente (angeion) y semilla (sperma), queriendo esto decir que son plantas con semillas guardadas en un recipiente, es decir, protegidas. La estructura protectora es el carpelo y en las plantas modernas est confinado a la flor. Una vez polinizadas, las flores pierden sus rganos externos (cliz, corola y estambres), tras lo cual el carpelo se modifica para producir el fruto con las semillas en su interior, que son las encargadas de la dispersin de los nuevos individuos. 9.5.1. La flor es un conjunto de hojas modificadas que alberga a los esporangios. La definicin de flor no es muy clara a pesar de ser el rgano caracterstico de las angiospermas, pero se puede describir como una estructura del vstago (parte area de la planta) con una disposicin de los esporangios que difiere de las otras plantas, la cual adems posee estructuras accesorias que protegen los rganos reproductores o atraen vectores para la dispersin de los granos de polen. La flor se compone de un eje caulinar llamado pednculo, el cual est ensanchado apicalmente formando una plataforma que se denomina tlamo o receptculo, sobre la cual se insertan cuatro grupos de rganos denominados verticilos florales, que son el cliz, la corola, el androceo y el gineceo (Figura 9.19). Todos los verticilos son antfilos, es decir, hojas modificadas que sirven para contribuir al proceso reproductor de la planta. Uno o varios verticilos florales pueden estar ausentes o muy modificados segn la especie. Los verticilos ms externos son estriles y reciben en conjunto el nombre de perianto, haciendo alusin a que rodean a la parte reproductiva. Son ellos el cliz y la corola. El cliz est constituido por spalos, los cuales son casi siempre verdes y de apariencia foliosa, si bien algunos spalos pueden ser casi idnticos a los ptalos y slo se distinguen de ellos por su posicin ms externa. En tal caso ambos verticilos se denominan tpalos. Los spalos a veces reemplazan a los ptalos en la funcin de atraer polinizadores y pueden estar fusionados y presentar variaciones de simetra. Cuando la flor no ha abierto, los spalos constituyen la cubierta protectora ms externa del capullo floral. En algunas plantas los spalos se pierden en el momento de la abertura floral. El siguiente verticilo hacia adentro es la corola, la cual est formada por los ptalos y su funcin es generalmente atraer polinizadores. Los ptalos pueden ser libres o estar fusionados y determinan el tipo de simetra floral, la cual puede ser radial (actinomorfa) y bilateral (cigomorfa). Los ptalos presentan muy variadas modificaciones morfolgicas entre las especies, lo cual se supone es el resultado de la coevolucin entre las plantas y sus visitantes florales. Sus muy variados colores se dan porque las clulas de los ptalos son especialmente ricas en cromoplastos, plastidios especializados en almacenar pigmentos. El tercer verticilo es conocido como androceo y lo forman los estambres, rganos reproductores constituidos por un filamento y una antera (Figura 9.20). El filamento est constituido por epidermis, parnquima y una traza vascular, es generalmente de forma alargada y sostiene en su extremo superior la antera. sta consta de dos masas denominadas tecas, cada una de las cuales posee dos sacos polnicos o microsporangios, en los cuales se generan las microsporas. Las tecas estn unidas entre s por un tejido denominado conectivo y segn la forma en que el filamento las sostiene se clasifica a las anteras en basifijas (sostenidas por la base), dorsifijas (sostenidas por el dorso) y verstiles o en balancn (sostenidad por la parte media y pueden moverse a lado y lado).
Figura 9.19. Flor del borrachero (Brugmansia sp.). En la parte superior la flor est en estado natural: A) cliz con sus spalos fusionados (sinspalo), B) corola de ptalos tambin fusionados (simptala), en la parte inferior est abierta para mostrar detalles: C) Punto de unin entre el ovario y el estilo, D) cliz disectado, E) corola abierta, F) estambre sealado cerca de la insercin de la antera en el filamento. Esta solancea posee los cuatro verticilos en sus flores, razn por la cual se dice que es una flor completa. Al desarrollarse, cada microspora genera un grano de polen o gametfito masculino, tambin llamado microgametfito. El grano de polen es de tal importancia en biologa vegetal, que en torno a l se ha desarrollado una disciplina propia dedicada a su estudio, denominada palinologa. El polen est constituido por dos capas protectoras, la externa denominada exina y la interna llamada intina. La exina est compuesta de esporopolenina, sustancia extremadamente resistente, compuesta al parecer de carotenoides polimerizados, que permite la preservacin de los granos de polen durante muchos de aos, razn por la cual los hace tiles en estudios de paleobiologa. En la exina se encuentran las aperturas que permitirn la germinacin del polen, denominadas colpos si son alargadas o poros si son redondeadas. La ornamentacin de la exina prcticamente define la morfologa del grano de polen y es de gran importancia taxonmica (Figura 9.20). La capa interna, la intina, est constituida por celulosa y pectina principalmente y es menos resistente que la exina.
Figura 9.20. Detalle mediante fotografa electrnica de barrido de los estambres y el polen. La figura de la parte izquierda muestra la parte terminal de los filamentos y su insercin basifija en las anteras de Penta sp. Se observa algunos granos de polen esparcidos. A la derecha hay una composicin en la que se muestra las diversas morfologas de los granos de polen. La escultura y forma del polen es de utilidad taxonmica y est determinada por la cubierta externa, denominada exina. Fotos cortesa del Dartmouth Electron Microscope Facility, Darmouth College.
Figura 9.21. Esquema en el que se muestra la disposicin de los estambres en la flor. A) Estambres libres, B) monadelfos (fusionados en un solo grupo), C) diadelfos (fusionados en dos grupos), D) triadelfos (en tres grupos). Los estambres varan en nmero desde uno en algunas gramneas hasta cientos en las mimosceas y pueden estar libres o fusionados entre s o con otros rganos florales (Figura 9.21). Cuando estn fusionados en un solo grupo son monadelfos, en dos grupos son diadelfos, en tres triadelfos y en ms de tres poliadelfos. La liberacin del polen se hace por lneas de dehiscencia longitudinal o transversal o por poros en el extremo de la antera (dehiscencia poricida). Comnmente se habla de los estambres como los rganos masculinos de las plantas, pero la verdad es que ellos carecen de sexo ya que pertenecen a la generacin esporfita, por lo que producen esporas y no gametos. Slo puede hablarse de sexo en rganos o individuos que
produzcan clulas reproductoras sexuales destinadas a fusionarse con otras clulas similares (gametos). Esto quiere decir que los estambres y las otras estructuras del esporfito, incluyendo el ovario y sus vulos, carecen de sexualidad ya que slo la generacin gametfita produce gametos: el gametfito femenino producir ovoclulas mientras que el masculino generar clulas espermticas. El cuarto verticilo es el pistilo o gineceo (Figura 9.19), el cual est formado por uno o varios carpelos, que son las estructuras reproductoras tradicionalmente llamadas femeninas, pero al igual que con los estambres, producen esporas y no gametos femeninos. El carpelo es una hoja modificada que sostena los esporangios y que se pleg sobre s misma para cerrarse y originar una cavidad protectora que actualmente cubre el megasporangio y la megaspora que se origina de l. Esta megaspora y su producto, el gametfito femenino, jams son liberados de la planta madre. El carpelo se modific mucho durante la evolucin y en la actualidad se considera que est compuesto de tres partes: 1) el ovario, que es la parte basal que yace sobre el receptculo y contiene los vulos, 2) el estilo, rgano generalmente alargado de posicin intermedia y caracterizado por el tejido transmisor que facilita el crecimiento del tubo polnico, aunque puede estar ausente en algunas especies, y 3) el estigma, que es la parte superior y presenta gran variedad formas, cuya funcin es recibir el grano de polen. La superficie del estigma es de naturaleza glandular en muchas especies y secreta sustancias ricas en azcares (Figura 9.22).
Figura 9.22. Fragmento del estigma de una monocotilednea de la familia lilicea. Se aprecia el conjunto de prolongaciones papilares en su epidermis, las cuales estn involucradas en el apropiado recono-cimiento del grano de polen. El gineceo es la parte ms interna de la flor y al desarrollarse dar origen al fruto, que es el ovario maduro, el cual en su interior contendr las semillas u vulos fecundados. El vulo consta de un tallo que lo une a la pared del ovario, denominado funculo, adems de dos envolturas protectoras llamadas tegumentos que se encuentran interrumpidas en uno de los polos formando el micrpilo. En el centro del vulo hay una masa de clulas denominada megasporangio o nucela, en la cual se desarrollar una clula madre de la megaspora que dar origen al megagametfito (Figura 9.23). Esta es la estructura que presentan la mayora de los vulos en las plantas modernas y la disposicin espacial de ellos dentro del ovario es usada para clasificarlos as: antropos, cuando el micrpilo se orienta cercano al funculo que lo sostiene y orttropos cuando el micrpilo y el funculo quedan opuestos (Figura 9.24). Hay incluso una posicin intermedia en los vulos anftropos, pero en las angiospermas el vulo se caracteriza por ser mayoritariamente antropo.
Figura 9.23. Esquema de la estructura del vulo de una angiosperma. A) Pared del ovario, B) placenta, C) funculo, D), tegumentos, E) micrpilo, F) extremo de la claza. Rodeado por los tegumentos est el saco embrionario, el cual posee tres antpodas en el extremo de la claza, dos ncleos polares en el centro y dos sinrgidas flanqueando a la ovoclula (en negro), en el extremo micropilar. Esta es la caracterstica estructura de 7 clulas y 8 ncleos del gametfito femenino o saco embrionario.
Figura 9.24. Esquema que muestra la insercin de los vulos en el ovario vegetal (placentacin), as como su orientacin espacial. A) vulos de placentacin parietal, B) placentacin libre central, C) placentacin axilar, D) vulo antropo y E) orttropo. Las flores pueden ser solitarias o pueden estar dispuestas en grupos densos llamados inflorescencias, las cuales son de importancia taxonmica. El eje principal de la inflorescencia es el pednculo, mientras que los ejes menores que se originan de l y que sostienen las flores individuales se denominan pedicelos. La disposicin de los pedicelos y la secuencia de maduracin de las flores son diferentes para cada uno de los tipos de inflorescencias (Figura 9.25). Segn su tipo de desarrollo las inflorescencias se clasifican en: 1) indefinidas o
racemosas (racimo, espiga, espdice, corimbo, umbela y captulo), 2) definidas o cimosas (cima bpara, cima escorpioide, drepanio, cima helicoidal y ripidio).
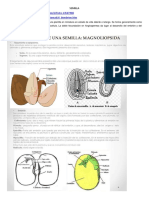
Figura 9.25. Esquema que muestra los principales tipos de inflorescencias. Los puntos negros son las flores, representando los ms grandes a las flores que maduran primero. A) Racimo, B) corimbo, C) pancula (racimo de racimos), D) umbela compuesta, E) cima helicoidal, F) cima escorpioide, G) espiga, H) espdice (espiga con eje carnoso), I) cima bpara, J) umbela, K) captulo. (Dibujo cortesa de M. Morales). 9.5.2. La semilla trajo numerosas ventajas reproductivas a las plantas. La semilla se origina del vulo fecundado y es probablemente el rgano de dispersin ms especializado que existe en el reino vegetal. Ella consta de un embrin en estado de latencia que est protegido por una estructura especializada denominada cubierta seminal (Figura 9.26). Es comn que la semilla posea material nutritivo de reserva, el cual es conocido tambin como albumen. Dicho material de reserva puede constituir el endospermo, tejido triploide o a veces poliploide, cuya primera clula es producida por la unin de un ncleo espermtico con los dos
(o ms) ncleos polares del megagametfito, proceso que se conoce con el nombre de doble fecundacin. Las semillas que presentan endospermo se denominan albuminosas y en general poseen embrin pequeo como en el maz. Las que carecen de l se denominan exalbuminosas y su embrin es generalmente grande. Se ha visto que algunas semillas poseen el endospermo con su superficie agrietada, producto de invaginaciones de la cubierta seminal, denominndose en este caso ruminado, pero en la mayora de semillas es de superficie lisa. Algunas especies carecen de endospermo porque transfieren completamente el material nutritivo de l hacia los cotiledones del embrin, como las leguminosas. Alternativamente, las reservas alimenticias pueden estar en el perispermo, tejido nutritivo diploide de origen nucelar. Por lo regular el perispermo es consumido por el embrin durante su desarrollo, pero algunas semillas maduras pueden poseerlo, como en algunas especies de las familias Quenopodiaceae, Amaranthaceae y Phytolaccaceae. La cubierta seminal, tambin llamada episperma, se deriva de los tegumentos del vulo y tiene como funcin proteger al embrin contenido en su interior. Puede poseer dos capas, siendo la ms externa la testa y la ms interna el tegmen. Segn el origen del tejido mecnico de la cubierta seminal, la semilla se clasifica en exotestal, mesotestal o endotestal si dicho tejido deriva de la testa. Tambin puede ser exotgmica, mesotgmica y endotgmica si la cubierta protectora proviene del tegmen. Rara vez hay tejido mecnico en la testa y el tegmen simultneamente. El tejido mecnico puede estar constituido por clulas esclerticas en empalizada, fibras en variada orientacin, clulas cuboidales u osteobraquiesclereidas. Esta cubierta de la semilla protege al embrin de la deshidratacin, las lesiones mecnicas y la entrada de plagas y agentes infecciosos. Las semillas poseen muchas variaciones en tamao y forma, adems de ser comn que presenten estructuras accesorias (Figura 9.27). Una caracterstica muy visible es el hilo, cicatriz que se ubica en el sitio en que el funculo una la semilla a la placenta, el cual es muy notable en algunas plantas como las leguminosas. El micrpilo es observable en algunas semillas y su posicin con respecto al hilo depende del tipo de vulo (orttropo, antropo, etc.). Algunas semillas de leguminosas y cucurbitceas poseen una lnea curvada en ambas caras de la cubierta seminal denominada pleurograma, que delimita un rea con diferencias de color y textura con respecto al resto de la semilla. La cubierta seminal puede estar fusionada a veces al funculo, originando una estructura denominada rafe. Es comn encontrar semillas con partes asociadas a manera de excrecencias que sirven para la dispersin. Por ejemplo, de la parte externa de la testa se origina la sarcotesta, de consistencia carnosa y jugosa como en las guamas (Inga spp.) y el mamoncillo (Melicoccus bijugatus). Del tegumento externo tambin se origina la carncula, estructura oleosa, situada cerca al micrpilo. Cuando es el funculo el que origina la excrecencia, se denomina arilo, como en la badea, maracuy, curuba y granadilla, donde cubre totalmente la semilla, o como en el trompeto (Bocconia frutescens), donde la envuelve parcialmente. Adicionalmente, en la superficie seminal puede haber pelos como en el algodn (Gossypium) o expansiones de la testa que forman alas para facilitar la dispersin anemfila, como en las bignoniceas.
Figura 9.26. Esquema de un corte longitudinal de la semilla de una planta monocotilednea (maz, A-F) y una dicotilednea (frjol, G-I). A) Endospermo, B) Escutelo (cotiledn), C) pericarpo fusionado con la cubierta seminal, D) coleptilo, E) embrin, F), coleorriza, G) cotiledn, H) cubierta seminal, I) embrin (Dibujo cortesa de M Morales). El embrin es la razn de ser de la semilla y resulta de la multiplicacin celular del cigoto diploide originado en la fertilizacin. Constituye el esporfito joven y su grado de desarrollo en el momento de la dispersin vara mucho entre las especies. En muchas orqudeas, por ejemplo, el embrin es simplemente una masa de clulas indiferenciadas y sin tejido nutricio alguno, por lo que depende de una asociacin simbitica mutualista con un hongo para su germinacin. El caso opuesto de desarrollo embrionario se ve en otra monocotilednea, el maz, cuyo embrin presenta una clara diferenciacin en radcula y vstago, los cuales poseen estructuras protectoras denominadas coleorriza y coleptilo, respectivamente. Adems, el embrin del maz posee un cotiledn especializado en la absorcin de nutrientes a partir del endospermo en digestin, que se denomina escutelo, el cual puede presentar hasta seis pequeas hojas que empiezan a hacer fotosntesis inmediatamente despus de la germinacin. La estructura del embrin maduro se basa en un eje longitudinal con polaridad, diferenciado en una parte basal y una apical. El punto de separacin entre ellas est marcado por el nudo cotiledonar, punto de insercin de los cotiledones. Arriba del nudo se halla el epictilo, que presenta un meristemo y a veces una yema desarrollada denominada plmula. Por debajo del nudo se encuentra el hipoctilo, cuyo extremo forma la radcula o raz embrionaria. La transicin entre el hipoctilo y la radcula es a veces difcil de distinguir, por lo que se puede denominar eje hipoctilo radicular. La posicin del embrin dentro de la semilla hace que se les clasifique en semillas con embriones basales, axiales y perifricos.
Figura 9.27. Algunas semillas y frutos tropicales. En la guama (Inga sp.), la semilla posee una excrecencia que sirve como atractivo para los animales dispersores, al igual que en el mamoncillo (Melicoccus sp.). El chontaduro (Bactris gasipaes), en la parte superior derecha de la foto, es un fruto indehiscente con una sola semilla (Foto cortesa U.S.D.A.), mientras que el araz (Psidium cattleianum), es una baya con muchas semillas.
Figura 9.28. Fruto agregado en la guanbana (Annona). Estos frutos preceden de varios carpelos de una misma flor. Varias anonceas son de valor comercial por sus frutos, como A. muricata, A. squamosa y A. cherimola. (cambiar foto Foto cortesa del U.S.D.A.). La proporcin entre el endospermo y los cotiledones vara mucho entre las semillas de las diferentes especies. En muchas dicotiledneas, el endospermo desaparece gradualmente mientras alimenta al embrin y ayuda a crecer los cotiledones. stos se llenan de almidn protenas o aceites que alimentarn la plntula en sus primeras etapas de crecimiento. En las plantas monocotiledneas, sin embargo, el endospermo est presente en la semilla madura y ayuda a digerir y absorber nutrientes al embrin. De otra parte, algunas dicotiledneas presentan un estado intermedio, pues desarrollan cotiledones pero parte del endospermo est presente y es funcional durante la germinacin y el desarrollo de la plntula. 9.5.3. Con el fruto se optimiz la dispersin de la semilla.
Las semillas desnudas dependen de la gravedad, el viento o el agua para su dispersin, ya que carecen de atractivo para animales dispersores. Con la aparicin del fruto, las plantas desarrollaron variadas estrategias de dispersin de sus semillas, como la presencia de estructuras en forma de gancho que se adhieren a la piel de animales, o la provisin de tejidos comestibles que aument el inters de los animales por ese tipo de especies (Figuras 9.28-9.32). Ontogenticamente hablando, el fruto es el resultado del proceso de maduracin del ovario luego de la fertilizacin de la ovoclula. En dicho proceso las paredes del carpelo experimentan grandes modificaciones por accin de las fitohormonas, que originan la forma y tamao definitivos del fruto maduro. La pared del fruto est formada por tres capas denominadas endocarpo, mesocarpo y exocarpo, que en conjunto reciben el nombre de pericarpo. La naturaleza seca o carnosa del pericarpo es comnmente usada para clasificar los frutos. Aunque el objetivo del fruto es la dispersin de las semillas, se sabe del desarrollo de algunos que no las poseen, a los cuales se denomina frutos partenocrpicos. Algunos de ellos, sin embargo, son el resultado de procesos de hibridacin artificial con fines comerciales, como sucede con los frutos del banano y los pltanos (Musa spp.).
Figura 9.29. Fruto mltiple del anan o pia (Ananas comosus). Este tipo de fruto se forma por la fusin de los carpelos junto con otras estructuras como el eje de la inflorescencia y las brcteas de muchas flores de la inflorescencia. Dichos carpelos corresponden a cada uno de los casquetes hexagonales que forman la superficie de este fruto. Es importante mencionar que los frutos presentan gran cantidad de modificaciones y adaptaciones, que incluyen la presencia de partes florales diferentes del ovario, denominndose en tal caso frutos accesorios. La forma ms comnmente empleada en la clasificacin de los frutos es la que los agrupa en simples, agregados y mltiples, como se describe a continuacin. Los frutos simples se desarrollan de un carpelo o de varios carpelos fusionados, los agregados se originan de carpelos separados de un solo gineceo, mientras que los frutos mltiples provienen de gineceos de ms de una flor (inflorescencia), por lo que tambin pueden ser llamados infrutescencias. Ejemplos de frutos agregados son las polidrupas de las moras (Rubus spp.), y los sincarpos de la guanbana (Annona muricata) y los anones (Figura 9.28). Los frutos mltiples son el resultado de la fusin de las flores de una inflorescencia y algunos ejemplos son la sorosis del anan o pia (Ananas comosus) (Figura 9.29), y el scono de los brevos. Antes de pasar a los frutos simples, es necesario mencionar que existe un variado grupo de frutos llamados complejos o accesorios, los cuales derivan de uno a varios ovarios simples o compuestos y de algn otro tejido que procede de partes de la flor como el cliz, la corola, el
receptculo, etc. Los ms conocidos son el cinorrodon de las rosas, que est conformado por aquenios o nculas incluidos en una cavidad formada por el cliz y el receptculo floral; la bellota, que es una nuez modificada por una cpula acartonada; el pseudocarpo de las fresas (Fragaria), en el que varios aquenios estn inmersos en el carnoso receptculo cnico; la pseudodrupa, en la que hay un involucro como en el nogal cafetero (Juglans neotropica). Cabe anotar que el pomo de peras y manzanas (ver ms adelante), es considerado por algunos autores en este tipo de frutos tambin.
Figura 9.30. Frutos simples secos dehiscentes. A) Vaina de una leguminosa que al abrirse expone las vistosas semillas, B) cpsula de una orqudea, la cual puede liberar miles de semillas muy pequeas por sus suturas longitudinales cuando el fruto haya madurado completamente. Los frutos simples son los ms abundantes, se originan de una flor monocarpelar o pluricarpelar sincrpica y se les divide en secos y carnosos. Los frutos secos son de dos tipos, dehiscentes (Figura 9.30), si se abren cuando alcanzan la madurez, e indehiscentes, que no se abren. El folculo es un fruto seco que se abre por un solo lado, el de la sutura ventral, como sucede en las magnolias. En la vaina o legumbre la apertura es por ambos lados, como pasa en las leguminosas, aunque hay un tipo especial denominado lomento, en el que el fruto tiene septos transversales que delimitan segmentos con una semilla que se desprenden individualmente como sucede en el pega-pega (Desmodium spp.). La silicua es un fruto alargado que en la madurez tambin se abre por ambos lados pero cuyas semillas quedan adheridas a una estructura central y laminar llamada replo, tal como sucede en las brasicceas. Una variante de ella es la silcula, de forma redondeada y caracterstica del medalln del Papa (Lunnaria annua) y la bolsa del pastor (Capsella bursa-pastoris). La cpsula es otro fruto seco dehiscente que se abre por lneas longitudinales, transversales o por poros terminales, como en las papaverceas, familia a la cual pertenece la amapola (Papaver somniferum). Algunos frutos secos hacen dehiscencia parcial que a veces no es funcional, ya que la apertura del fruto no es lo suficientemente grande para permitir la salida de las semillas, como sucede en Eschweilera bogotensis (Figura 9.31). Entre los frutos secos indehiscentes el ms comn es el aquenio, que consta de una sola semilla que est adherida por el funculo a la pared del fruto, como en las ranunculceas y
poligonceas. Cuando el fruto seco posee prolongaciones aplanadas a manera de alas, se le conoce como smara y es tpico en gneros como Ulmus y en el urapn (Fraxinus chinensis). En las gramneas el fruto tiene fuertemente adherida la semilla en su interior y se conoce como grano o cariopsis. Otro ejemplo de fruto indehiscente es la nuez, caracterizada por poseer una pared ptrea, que est representada en los trpicos en el gnero Quercus de las fagceas. Cuando la nuez es pequea se le denomina ncula, como en el maran (Anacardium occidentale). El esquizocarpo es un fruto seco pluricarpelar que en la madurez desprende trozos o segmentos de su cuerpo (mericarpos), cada uno de los cuales posee una semilla en su interior, como en los geranios y algunas malvceas. La cipsela es otro fruto seco indehiscente proveniente de un gineceo bicarpelar, unilocular y con ovario nfero. Posee una sola semilla y es tpico de las asterceas como el diente de len, el girasol y las margaritas.
Figura 9.31. Fruto en cpsula de Eschweilera bogotensis, una Lecythidaceae nativa de Colombia. Este fruto es dehiscente por una sutura transversal pero la dehiscencia no es funcional siempre, ya que algunas veces las semillas slo caen cuando los agentes dispersores las toman o cuando el fruto se descompone y cae al suelo. El otro gran grupo de frutos simples es el de lo carnosos, entre los que estn las bayas, las drupas y los pomos. Las bayas poseen varias semillas y son completamente carnosas, como en el tomate (Lycopersicon esculentum, recientemente renombrado Solanum lycopersicon) y las uvas (Vitis vinifera). Un tipo especial de baya es el pepnide, carnoso en su interior pero con una cubierta coricea (dura y gruesa), tpico de las cucurbitceas. Otro tipo de baya especial es el hesperidio, el fruto de los ctricos, el cual es una baya pluricarpelar. Las drupas poseen su capa interna muy dura y gruesa, adherida a la semilla que generalmente es individual, como el durazno (Prunus sp.) y el coco (Cocos nucifera). Los pomos son tpicos de algunas rosceas y su parte carnosa proviene de la base agrandada del perianto, mientras que la parte interna del fruto (endocarpo) es membranosa y acartonada, como en la manzana (Malus sylvestris) y la pera (Pyrus communis). Aunque la norma es que los frutos carnosos no sean dehiscentes, hay algunas especies tropicales en la familia Monimiaceae que s los abren en la madurez (Figura 9.32).
Figura 9.32. Fruto carnoso dehiscente. Esta especie neotropical de la familia Monimiaceae muestra uno de los pocos casos en que un fruto carnoso se abre para exponer sus semillas a los agentes dispersores. Los frutos representaron una gran ventaja para las plantas, ya que protegan a las semillas y a la vez constituyeron un gran atractivo para los animales dispersores, por lo que es apenas lgico que las especies que los poseen son las que ms exitosamente han colonizado el planeta. La biologa reproductiva es un tema de investigacin inacabado, que ser necesario agotar si se quiere entender completamente la dinmica de la vegetacin moderna. BIBLIOGRAFA Becerra N & Chaparro de Valencia M. 1999. Morfologa y anatoma vegetal. Facultad de Ciencias, Universidad Nacional de Colombia, Bogot, DC., Colombia. Bello-G MA. 2004. Esporognesis, gametognesis y embriognesis: Hacia la reproduccin sexual en plantas. Departamento de Biologa, Universidad Nacional de Colombia, Bogot, DC., Colombia. Esau K. 1976. Anatoma vegetal. Ediciones Omega, S.A., Barcelona, Espaa. Izco J, Barreno E, Brugus M, Costa M, Devesa JA, Fernndez F, Gallardo T, Llimona X, Prada C, Talavera S & Valds B. 2004. Botnica 2 ed. McGraw-Hill Interamericana de Espaa, S.A.U. Madrid, Espaa. Judd WS, Campbell CS, Kellogg EA & Stevens PF. 2002. Plant systematics. A phylogenetic approach. Sinauer Associates Inc., Sunderland, MA, EUA. Mauseth JD. 2003. Botany. An introduction to plant biology, 3 ed. Jones and Barlett Publishers, Sudbury, EUA. Moore R, Clark WD, Stern KR & Vodopich D. 1995. Botany. WCM Brown Communications, Inc., Dubuque, EUA. Raven PH, Evert RF & Eichorn SE. 1999. Biology of plants, 6 ed. W.H. Freeman and co. and Worth Publishers, Nueva York, NY, EUA.
Rost TL, Barbour MG, Thornton RM, Weier TE & Stocking CR. 1985. Botnica. Introduccin a la biologa vegetal. Editorial Limusa, Mxico DF., Mxico. Sitte P, Ziegler H, Ehrendorfer & Bresinsky A. 1994. Strasburger tratado de botnica, 8 ed. Ediciones Omega, S.A., Barcelona, Espaa.
Вам также может понравиться
- Especialidad en Flores z4Документ8 страницEspecialidad en Flores z4Jeremias Mesa Medina EncarnacionОценок пока нет
- Examen Especialidad de Semillas - Print - QuizizzДокумент3 страницыExamen Especialidad de Semillas - Print - QuizizzGessica MoreiraОценок пока нет
- Semiología PDFДокумент1 597 страницSemiología PDFIsabel TrujilloОценок пока нет
- GIMNOSPERMASДокумент10 страницGIMNOSPERMASMariana CondeОценок пока нет
- Diagrama FloralДокумент7 страницDiagrama FloralAnais Briggit Jacinto ChavezОценок пока нет
- Guia Completa BriofitasДокумент10 страницGuia Completa BriofitasAnGyziitha HdezОценок пока нет
- Estructuras ReproductivasДокумент27 страницEstructuras ReproductivasJorge RiveraОценок пока нет
- EMBRIOFITOSДокумент12 страницEMBRIOFITOSClaudia GuerreroОценок пока нет
- Teoría Musgos Hepáticas y AntocerosДокумент14 страницTeoría Musgos Hepáticas y AntocerosJOSUE DANIEL CERDAN ALVAREZОценок пока нет
- BriófitosДокумент4 страницыBriófitosMiguel Angel SarmientoОценок пока нет
- Ferriol López - Ciclo de Vida de Los MusgosДокумент6 страницFerriol López - Ciclo de Vida de Los MusgosValeria GonzalezОценок пока нет
- BriófitosДокумент10 страницBriófitosSebastian ArdilaОценок пока нет
- Biologia Laboratorio, Diversidad Ded La Vida Plantas y Animales, F1Документ13 страницBiologia Laboratorio, Diversidad Ded La Vida Plantas y Animales, F1Alondra Quispe AvalosОценок пока нет
- TEMA 3 - Briofitos Y PteridofitosДокумент121 страницаTEMA 3 - Briofitos Y PteridofitosClaireОценок пока нет
- BG Tema 13B - Reproducción VegetalДокумент22 страницыBG Tema 13B - Reproducción VegetalIsabel BelloОценок пока нет
- Ciclos de Vida de PlantaeДокумент11 страницCiclos de Vida de PlantaeIsmael Castillo ReyesОценок пока нет
- 5Документ12 страниц5Carlos AlcaОценок пока нет
- Tpn6 - Angiospermas Basales y Clado Magnoliides - 2021Документ17 страницTpn6 - Angiospermas Basales y Clado Magnoliides - 2021Juan FretesОценок пока нет
- RodofitasДокумент7 страницRodofitasAndre RodriguezОценок пока нет
- HepaticasДокумент27 страницHepaticasNicolas Cotrino VaquiroОценок пока нет
- PlantasguiafinalДокумент11 страницPlantasguiafinalLucía CampbellОценок пока нет
- MUSGOДокумент2 страницыMUSGOcieloОценок пока нет
- Laboratorio 2. BriófitosДокумент9 страницLaboratorio 2. BriófitosManuel Felipe Echeverry GracésОценок пока нет
- Práctica BriofitasДокумент2 страницыPráctica Briofitastoro_strongОценок пока нет
- EMBRIOFITOSДокумент22 страницыEMBRIOFITOSClaudia GuerreroОценок пока нет
- ALGASДокумент5 страницALGAS.:("*"BLacK BuLLeT"*"):.0% (1)
- 15 Plantas Semillas Texto PDFДокумент6 страниц15 Plantas Semillas Texto PDFneyОценок пока нет
- Tema 10Документ11 страницTema 10María Gómez RomeroОценок пока нет
- TEMA 9 (AyM) AcabadoДокумент24 страницыTEMA 9 (AyM) AcabadoSara Domínguez AlonsoОценок пока нет
- Reino VegetalДокумент36 страницReino VegetalAlex rodrigo MontenegroОценок пока нет
- MicrobiologiaДокумент6 страницMicrobiologiaTatiana LambrañoОценок пока нет
- Actividad+de+plantas 3Документ18 страницActividad+de+plantas 3Geovani RodriguezОценок пока нет
- 8°actividad Sobre Musgos y HelechosДокумент4 страницы8°actividad Sobre Musgos y Helechosvanessa paola perez vivanqueОценок пока нет
- Rebigo 2008 08Документ10 страницRebigo 2008 08JOSE MIGUEL GIRALDO AYERBEОценок пока нет
- Los ProtistasДокумент5 страницLos ProtistasJULIOCRODRIGUEZОценок пока нет
- Musgos y HepaticasДокумент21 страницаMusgos y Hepaticasparared0% (1)
- Tema36 Biologia y EcologiaДокумент16 страницTema36 Biologia y EcologiaMaría Jesús RuizОценок пока нет
- Informe - Nematodos en Vid-1Документ6 страницInforme - Nematodos en Vid-1Mayra Guzmán IbañezОценок пока нет
- Informe de Labo. BiologiaДокумент4 страницыInforme de Labo. Biologiamanuel delacruzОценок пока нет
- Division CloropitaДокумент47 страницDivision CloropitaLuiggi Ronaldo Rios DiazОценок пока нет
- Las AngiospermasДокумент30 страницLas AngiospermasKAREN LIZETH QUINTERO PERALTAОценок пока нет
- Cuestionario BiologiaДокумент2 страницыCuestionario BiologiaEzequias100% (2)
- Informe 6 CharophytasДокумент8 страницInforme 6 CharophytasJosselyn Alvarez MarquezОценок пока нет
- Alternancia de Generaciones DescripcionДокумент10 страницAlternancia de Generaciones DescripcionhernanОценок пока нет
- Cuestionario N°08Документ1 страницаCuestionario N°08Sofía ValentinaОценок пока нет
- SemillasДокумент60 страницSemillasLaura GuerreroОценок пока нет
- Ciclos de VidaДокумент19 страницCiclos de Vidaanystar20dem.15Оценок пока нет
- HT PlantasДокумент2 страницыHT PlantasDIEGO ANDRES ALONZO MEDINILLAОценок пока нет
- Seminario Biodiversidad y Ciclos de VidaДокумент4 страницыSeminario Biodiversidad y Ciclos de VidaSilvina B TamburiniОценок пока нет
- 2 Propagación de Plantas 2023Документ8 страниц2 Propagación de Plantas 2023Pablo Vasquez cruzОценок пока нет
- Cuestionario 1 Sobre El Tema de La SemillaДокумент3 страницыCuestionario 1 Sobre El Tema de La SemillaJose MarcelinoОценок пока нет
- Evol SemillaДокумент8 страницEvol SemillaElFaboОценок пока нет
- Briofitas 2015 DiapoДокумент20 страницBriofitas 2015 DiapoMiguel Angel SarmientoОценок пока нет
- U.3 Investigación Documental AgronomíaДокумент26 страницU.3 Investigación Documental AgronomíaJesus Ernesto VerdugoОценок пока нет
- Tema 17Документ3 страницыTema 17Fernando LuengoОценок пока нет
- Ensayo de La Fauna GuatemaltecaДокумент16 страницEnsayo de La Fauna GuatemaltecaOscar ArguetaОценок пока нет
- Biologia LeonelisДокумент17 страницBiologia LeonelisLeonelis MoleroОценок пока нет
- Embryo Phy TaДокумент15 страницEmbryo Phy TaWalter Mija MarchanОценок пока нет
- MuДокумент27 страницMusu kiОценок пока нет
- 11 Briofitos TextoДокумент4 страницы11 Briofitos TextoAnGyziitha HdezОценок пока нет
- Porifera 2010Документ10 страницPorifera 2010Isabel TrujilloОценок пока нет
- Genes Simples IIДокумент8 страницGenes Simples IIIsabel TrujilloОценок пока нет
- Obtención de Cloruro de TerbutiloДокумент5 страницObtención de Cloruro de TerbutiloIsabel TrujilloОценок пока нет
- POLENESДокумент7 страницPOLENESItzel MartinezОценок пока нет
- Practica 8 BotanicaДокумент7 страницPractica 8 BotanicaMacias Guadalupe LizbethОценок пока нет
- QUINUAДокумент6 страницQUINUAAna Lizeth Marín GómezОценок пока нет
- Germinacion de Semillas en Plantas DicotiledoneasДокумент6 страницGerminacion de Semillas en Plantas DicotiledoneasBRANDON FELIPE MOSQUERAОценок пока нет
- Polinización de Las Orquídeas Zapatilla - PDF Versión 1Документ6 страницPolinización de Las Orquídeas Zapatilla - PDF Versión 1Diego GutierrezОценок пока нет
- Por Qué Los Polinizadores Visitan A Las FloresДокумент3 страницыPor Qué Los Polinizadores Visitan A Las FloresLewisKarlОценок пока нет
- Glosario Términos BotánicosДокумент4 страницыGlosario Términos BotánicosLaura PrestonОценок пока нет
- Exposicion de SemillaДокумент28 страницExposicion de SemillaRaul RangelОценок пока нет
- Se MillaДокумент10 страницSe MillaShirley Yesenia Heredia AltamiranoОценок пока нет
- Multiplicación Sexual Del DuraznoДокумент12 страницMultiplicación Sexual Del DuraznoVilma RodriguezОценок пока нет
- Práctica 7 La Flor y Sus PartesДокумент3 страницыPráctica 7 La Flor y Sus PartesKatheryn Dahiana Chamorro Correa100% (1)
- Evol SemillaДокумент8 страницEvol SemillaElFaboОценок пока нет
- ArticCientGilmerMacco CAMUCAMUДокумент17 страницArticCientGilmerMacco CAMUCAMUJulio César Segura SifuentesОценок пока нет
- Información de Parcial 2 de SemillasДокумент8 страницInformación de Parcial 2 de SemillasYeni Paola Giron JoaquiОценок пока нет
- Monografia de Forraje Verde HidroponicoДокумент6 страницMonografia de Forraje Verde HidroponicoIvan Martinez Huallpa0% (1)
- Infome Laboratorio 1-Germinación SemillasДокумент9 страницInfome Laboratorio 1-Germinación Semillasmartinacos21100% (3)
- Estructura de Los Órganos VegetalesДокумент35 страницEstructura de Los Órganos VegetalesDaniel Jonatan VelazquezОценок пока нет
- Flora de Veracruz ActinidiaceaeДокумент28 страницFlora de Veracruz ActinidiaceaeLiebre Messi BonaparteОценок пока нет
- FlorДокумент8 страницFlorAllisonSanchezGuzmanОценок пока нет
- Monocots y EudicotsДокумент16 страницMonocots y Eudicotsjacobo caicedo angelОценок пока нет
- Reproducción Por EsporasДокумент9 страницReproducción Por EsporasJuanjo Vanini0% (1)
- Informe de Laboratorio 2Документ6 страницInforme de Laboratorio 2Re Ramirez TumbaoОценок пока нет
- Germinación de Las Semillas de PalmaДокумент14 страницGerminación de Las Semillas de PalmaHugo MoraОценок пока нет
- Separata MangoДокумент12 страницSeparata MangoEileen WilsonОценок пока нет
- Actividad de Reforzamiento - Formas de Reproducción en Las PlantasДокумент1 страницаActividad de Reforzamiento - Formas de Reproducción en Las PlantasRocio Ricse GalarzaОценок пока нет
- Tema 10. Reproducción VegetalДокумент80 страницTema 10. Reproducción VegetalLydia Pérez OrdoñezОценок пока нет