Вам также может понравиться
- 97 312 PDFДокумент5 страниц97 312 PDFdupuytrenОценок пока нет
- Patofisiologi StrkeДокумент14 страницPatofisiologi StrkeTeuku ILza Nanta SatiaОценок пока нет
- Differences: Biological Between Ischemia, Hypoglycemia, EpilepsyДокумент9 страницDifferences: Biological Between Ischemia, Hypoglycemia, EpilepsyAyie JayaОценок пока нет
- Qin, Et Al. 2022Документ29 страницQin, Et Al. 2022Ignacio BarbieriОценок пока нет
- Neural ApoptosisДокумент6 страницNeural ApoptosisJay HardkickОценок пока нет
- Pathophysiology, Bioelectricidad, Review Stroke TINS99Документ7 страницPathophysiology, Bioelectricidad, Review Stroke TINS99Carla Saramy MachillandaОценок пока нет
- Cell Death Pathways in Ischemic Stroke and Targeted PharmacotherapyДокумент18 страницCell Death Pathways in Ischemic Stroke and Targeted PharmacotherapyHeath HensleyОценок пока нет
- Khoshnam2017 Article PathogenicMechanismsFollowingIДокумент20 страницKhoshnam2017 Article PathogenicMechanismsFollowingILovin BiancaОценок пока нет
- Saraf Kel 4 AlzheimerДокумент15 страницSaraf Kel 4 AlzheimertirtapinkanОценок пока нет
- Excitotoxicity and neuronal death mechanisms in epilepsyДокумент8 страницExcitotoxicity and neuronal death mechanisms in epilepsyKhrîîz Ëvïi BïbiBüûОценок пока нет
- Pa Tho Physiology of Focal Cerebral Ischemia A Therapeutic PerspectiveДокумент10 страницPa Tho Physiology of Focal Cerebral Ischemia A Therapeutic PerspectiveMalisa LukmanОценок пока нет
- Cell Death Mechanisms and Emerging TherapiesДокумент23 страницыCell Death Mechanisms and Emerging TherapiesPaola MontenegroОценок пока нет
- Mecanismos de Muerte Celular en Las Enfermedades Neurodegenerativas Apoptosis o NecrosisДокумент16 страницMecanismos de Muerte Celular en Las Enfermedades Neurodegenerativas Apoptosis o NecrosisAle Romero AguirreОценок пока нет
- Dapus 2Документ8 страницDapus 2Beirnes Fernando Sembiring MОценок пока нет
- 2010 Hypoxic Ischemic Brain Injury Pathophysiology Neuropathology and Mechanisms KM Busl PDFДокумент9 страниц2010 Hypoxic Ischemic Brain Injury Pathophysiology Neuropathology and Mechanisms KM Busl PDFDaniel OUОценок пока нет
- 17 - KULIAH PANEL - Neuronal Injury and Adaptation - PDFXДокумент56 страниц17 - KULIAH PANEL - Neuronal Injury and Adaptation - PDFXluthfianisa azhariОценок пока нет
- Contribution of TRPC Channels in Neuronal Excitotoxicity Associated With Neurodegenerative Disease and Ischemic StrokeДокумент17 страницContribution of TRPC Channels in Neuronal Excitotoxicity Associated With Neurodegenerative Disease and Ischemic Strokediego.battiatoОценок пока нет
- Pathophysiology of StrokeДокумент11 страницPathophysiology of StrokeAgronaSlaughterОценок пока нет
- 1 Apoptosis DesДокумент7 страниц1 Apoptosis DessashiwiiОценок пока нет
- Apoptosis in The NcsДокумент8 страницApoptosis in The NcsMiguel Angel Villarreal YbazetaОценок пока нет
- C R B S: Urrent Eview in Asic CienceДокумент5 страницC R B S: Urrent Eview in Asic CienceNurul FajriОценок пока нет
- BR Med Bull 1997 Rubin 617 31Документ15 страницBR Med Bull 1997 Rubin 617 31karinadapariaОценок пока нет
- Jurnal PDFДокумент9 страницJurnal PDFFerdy Arif FadhilahОценок пока нет
- BNIP3L NIX degradation leads to mitophagy deficiency in ischemic brainsДокумент14 страницBNIP3L NIX degradation leads to mitophagy deficiency in ischemic brainseОценок пока нет
- Human Molecular Genetics study reveals multi-systemic mitochondrial disorder due to dominant POLG mutationДокумент16 страницHuman Molecular Genetics study reveals multi-systemic mitochondrial disorder due to dominant POLG mutationMihai LazarОценок пока нет
- Pathophysiology of Stroke 2Документ12 страницPathophysiology of Stroke 2AgronaSlaughterОценок пока нет
- Ciclo Celular y CáncerДокумент12 страницCiclo Celular y CáncerAlfonso De la FuenteОценок пока нет
- Ischemic StrokeДокумент8 страницIschemic StrokeRinavi Adrin RirinОценок пока нет
- J Gerontol A Biol Sci Med Sci-2001-Pollack-B475-82Документ8 страницJ Gerontol A Biol Sci Med Sci-2001-Pollack-B475-82divinebutler13Оценок пока нет
- Stem Cell Treatment Alzheimer's Disease - ASCI - Asian Stem Cell InstituteДокумент39 страницStem Cell Treatment Alzheimer's Disease - ASCI - Asian Stem Cell Institutea73072Оценок пока нет
- Contribucion de La SepsisДокумент2 страницыContribucion de La SepsisRichard Sossa VillarrealОценок пока нет
- Subarachnoid Hemorrhage and Cerebral Vasospasm - Literature ReviewДокумент6 страницSubarachnoid Hemorrhage and Cerebral Vasospasm - Literature ReviewzixzaxoffОценок пока нет
- Pa Tho Physiology of Traumatic Brain Injury2Документ22 страницыPa Tho Physiology of Traumatic Brain Injury2n2s_nqОценок пока нет
- European Journal of Internal Medicine: Stephanie Seneff, Glyn Wainwright, Luca MascitelliДокумент7 страницEuropean Journal of Internal Medicine: Stephanie Seneff, Glyn Wainwright, Luca MascitelliApril ayuОценок пока нет
- Cytotoxic Edema Mechanisms in Brain InjuryДокумент17 страницCytotoxic Edema Mechanisms in Brain InjuryYopi ArdhiaswariОценок пока нет
- Receptor HNДокумент7 страницReceptor HNLeilane GlienkeОценок пока нет
- Neuropathologic Changes in ADДокумент4 страницыNeuropathologic Changes in ADscientist786Оценок пока нет
- 5-Aminolevulinic Acid-Mediated Sonodynamic Therapy Inhibits RIPK1/RIPK3-Dependent Necroptosis in THP-1-Derived Foam CellsДокумент13 страниц5-Aminolevulinic Acid-Mediated Sonodynamic Therapy Inhibits RIPK1/RIPK3-Dependent Necroptosis in THP-1-Derived Foam Cellsdevi eraОценок пока нет
- Ad ReviewДокумент14 страницAd ReviewKhushОценок пока нет
- Neurodegeneration: Progressive Loss of Neurons and Their FunctionsДокумент8 страницNeurodegeneration: Progressive Loss of Neurons and Their FunctionsRommelBaldagoОценок пока нет
- Dutta Et Al Mitochondrial Dysfunction in MS AnnNeurol06Документ12 страницDutta Et Al Mitochondrial Dysfunction in MS AnnNeurol06WZОценок пока нет
- Brain Injury Following Cardiac Arrest: Pathophysiology For Neurocritical CareДокумент10 страницBrain Injury Following Cardiac Arrest: Pathophysiology For Neurocritical CareDianis DiОценок пока нет
- Current Status and Challenges of Stem Cell Treatment For Alzheimer's DiseaseДокумент19 страницCurrent Status and Challenges of Stem Cell Treatment For Alzheimer's DiseaseHelical Nail FemurОценок пока нет
- Cellular Injury, Necrosis and ApoptosisДокумент34 страницыCellular Injury, Necrosis and ApoptosisSuraj UsmanОценок пока нет
- Stem cell therapy for spinal cord repair: a review of the past 10 yearsДокумент6 страницStem cell therapy for spinal cord repair: a review of the past 10 yearsOkhuarobo SamuelОценок пока нет
- Lee 2008Документ14 страницLee 2008Laura Duarte RojasОценок пока нет
- Pathophysiology of Epilepsy: An Updated Review: International Journal of Medical and Health ResearchДокумент5 страницPathophysiology of Epilepsy: An Updated Review: International Journal of Medical and Health ResearchGayathiri GovindarajuОценок пока нет
- NSAIDs' Role in Alzheimer's Disease via Anti-Amyloidogenic and Anti-Inflammatory EffectsДокумент16 страницNSAIDs' Role in Alzheimer's Disease via Anti-Amyloidogenic and Anti-Inflammatory Effectsdhiraj.2367Оценок пока нет
- Pathophysiology of Encephalopathy: N. G.F. AДокумент7 страницPathophysiology of Encephalopathy: N. G.F. Adoğancan uzunОценок пока нет
- Concepts of Brain DeathДокумент6 страницConcepts of Brain DeathBen OkangaОценок пока нет
- Mitochondrial Membrane PermeabilizationДокумент66 страницMitochondrial Membrane PermeabilizationGus Tavo BrugesОценок пока нет
- Dhami Et Al-2019-Journal of Neurochemistry PDFДокумент18 страницDhami Et Al-2019-Journal of Neurochemistry PDFJawad A. KhanОценок пока нет
- Non-Coding RNA Mediates Endoplasmic Reticulum Stress-Induced Apoptosis in Heart DiseaseДокумент11 страницNon-Coding RNA Mediates Endoplasmic Reticulum Stress-Induced Apoptosis in Heart DiseaseWaldo IzyОценок пока нет
- NIH Public Access: Caspase-1 Induced Pyroptotic Cell DeathДокумент15 страницNIH Public Access: Caspase-1 Induced Pyroptotic Cell DeathcutlisaafriannaОценок пока нет
- Quantitative Proteomic Analyses of Dynamic Signalling Events in Cortical Neurons Undergoing Excitotoxic Cell DeathДокумент19 страницQuantitative Proteomic Analyses of Dynamic Signalling Events in Cortical Neurons Undergoing Excitotoxic Cell DeathSaqr Al KhazalehОценок пока нет
- 1 s2.0 S0021915021003269 MainДокумент1 страница1 s2.0 S0021915021003269 MainIoanna NikdimaОценок пока нет
- New Concepts in Treatment of Pediatric Traumatic Brain InjuryДокумент28 страницNew Concepts in Treatment of Pediatric Traumatic Brain InjuryAlexander BenavidesОценок пока нет
- NHE6 PaperДокумент28 страницNHE6 PaperJon DrdОценок пока нет
- Cancer Epidemiol Biomarkers Prev 2008 Lee 2647 55Документ10 страницCancer Epidemiol Biomarkers Prev 2008 Lee 2647 55Kahubbi Fatimah Wa'aliyОценок пока нет
- LIPOMAДокумент21 страницаLIPOMAKahubbi Fatimah Wa'aliyОценок пока нет
- Autopsy Manual - 5 - 06Документ54 страницыAutopsy Manual - 5 - 06Kahubbi Fatimah Wa'aliyОценок пока нет
- NICE Guidelines PDFДокумент37 страницNICE Guidelines PDFKahubbi Fatimah Wa'aliyОценок пока нет
- Autopsy Manual - 5 - 06Документ54 страницыAutopsy Manual - 5 - 06Kahubbi Fatimah Wa'aliyОценок пока нет
- List of Anesthetic, Analgesic and Tranquilizer Drugs Veterinary)Документ25 страницList of Anesthetic, Analgesic and Tranquilizer Drugs Veterinary)drbadman77100% (2)
- WHO Dengue Guidelines 2013Документ160 страницWHO Dengue Guidelines 2013Jason MirasolОценок пока нет
- Space LesionsДокумент18 страницSpace LesionsKahubbi Fatimah Wa'aliyОценок пока нет
- June07-ACLS Provider Course PapersДокумент7 страницJune07-ACLS Provider Course PapersKahubbi Fatimah Wa'aliyОценок пока нет
- List of Anesthetic, Analgesic and Tranquilizer Drugs Veterinary)Документ25 страницList of Anesthetic, Analgesic and Tranquilizer Drugs Veterinary)drbadman77100% (2)
- NMJ Chemical Synapse Between Neurons and MuscleДокумент21 страницаNMJ Chemical Synapse Between Neurons and Musclekunjbihari sulakhiyaОценок пока нет
- The Nervous System: Lecture Presentation by Patty Bostwick-Taylor Florence-Darlington Technical CollegeДокумент196 страницThe Nervous System: Lecture Presentation by Patty Bostwick-Taylor Florence-Darlington Technical CollegeTrisha100% (8)
- Guide To General Histology and Microscopic AnatomyДокумент62 страницыGuide To General Histology and Microscopic AnatomybjuangorenabОценок пока нет
- Unit 5 Biology NotesДокумент21 страницаUnit 5 Biology Notes098789100% (1)
- 6 5 Nerves Hormones and HomeostasisДокумент19 страниц6 5 Nerves Hormones and Homeostasisapi-235355872Оценок пока нет
- Histo Lec Module 5 BookДокумент54 страницыHisto Lec Module 5 Bookchuchu lalaОценок пока нет
- Datasheet of Pc16f87xaДокумент2 страницыDatasheet of Pc16f87xasnehaОценок пока нет
- Pulsed Magnetic Therapy-Drug Free Healing and Pain Relief WebДокумент145 страницPulsed Magnetic Therapy-Drug Free Healing and Pain Relief WebMaryury Quintero Ramírez100% (5)
- BIO 2311 Lab Test 4 Study Guide Fall 2020Документ1 страницаBIO 2311 Lab Test 4 Study Guide Fall 2020Annabell CheungОценок пока нет
- MCQ of Neurochemistry and Signal Transduction-2022-FinalДокумент6 страницMCQ of Neurochemistry and Signal Transduction-2022-FinalKaif AliОценок пока нет
- Barker Revision NotesДокумент22 страницыBarker Revision NotesWaqas HaleemОценок пока нет
- NBSS AIA - Ivan Christian ChannelДокумент128 страницNBSS AIA - Ivan Christian ChannelASTAGINA NAURAHОценок пока нет
- Thalamus: What Is The Thalamus and What Are Its Main Functions?Документ8 страницThalamus: What Is The Thalamus and What Are Its Main Functions?sandeep1699Оценок пока нет
- Bossche1985 PDFДокумент57 страницBossche1985 PDFMohamad MostafaОценок пока нет
- Cholinergic Anticholinergic DrugsДокумент60 страницCholinergic Anticholinergic DrugsMD. RASEL MAHMUD MIMОценок пока нет
- @MBS MedicalBooksStore 2019 GettingДокумент364 страницы@MBS MedicalBooksStore 2019 GettingCF Ponce100% (4)
- Bioquimica Da DorДокумент92 страницыBioquimica Da DorLUCIENE CRISTALDO ALBUQUERQUEОценок пока нет
- Thoughts, Consciousness and MemoryДокумент28 страницThoughts, Consciousness and MemoryJorge De VeraОценок пока нет
- Principles of Neural Science: Eric R James H. Schwartz, M.D.,PanДокумент24 страницыPrinciples of Neural Science: Eric R James H. Schwartz, M.D.,PanThiago Alves SilvaОценок пока нет
- Pyc1501 PDFДокумент44 страницыPyc1501 PDFAlicial Olwandle40% (5)
- The Central Nervous System - Per Brodal, MD, PHDДокумент1 368 страницThe Central Nervous System - Per Brodal, MD, PHDHenryk BazalarОценок пока нет
- Basic Neuroscience CompiledДокумент71 страницаBasic Neuroscience CompiledDr.Rathod RanjithОценок пока нет
- A2 BIOLOGY CORE PRACTICAL SUMMARYДокумент3 страницыA2 BIOLOGY CORE PRACTICAL SUMMARYSQ100% (2)
- Pearson Nervous System ReviewerДокумент8 страницPearson Nervous System ReviewerViaBОценок пока нет
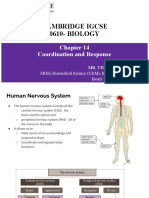
- CAMBRIDGE IGCSE BIOLOGY Chapter 14 Coordination and ResponseДокумент203 страницыCAMBRIDGE IGCSE BIOLOGY Chapter 14 Coordination and ResponseVentus Tan100% (1)
- Comparative Pharmacology For AnesthetistДокумент162 страницыComparative Pharmacology For AnesthetistGayatri PalacherlaОценок пока нет
- Neuromuscular Junction: Dr. Mustafa Al-Shehabat MSC, PH.DДокумент32 страницыNeuromuscular Junction: Dr. Mustafa Al-Shehabat MSC, PH.Dمحمد السنجلاوي100% (1)
- pharmacology-II Practical ManualДокумент35 страницpharmacology-II Practical ManualJunaed Ahmed0% (1)
- Ebook PDF Vanders Human Physiology The Mechanisms of Body Function 13th Edition PDFДокумент40 страницEbook PDF Vanders Human Physiology The Mechanisms of Body Function 13th Edition PDFmarie.simons156100% (27)
- MC. Midterm Set 5Документ18 страницMC. Midterm Set 5Wibert TrenggonoОценок пока нет