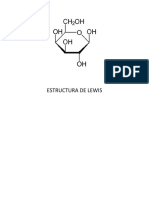
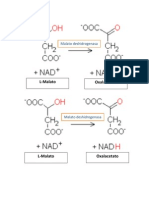
Вам также может понравиться
- CuestionarioДокумент5 страницCuestionarioMaríaGabrielaOrellanaОценок пока нет
- Taller SolucionesДокумент2 страницыTaller SolucionesDaniela0% (1)
- Mecanismos de Regulación EnzimáticaДокумент2 страницыMecanismos de Regulación EnzimáticaRobert Isaias Mendoza CardenasОценок пока нет
- Metabolismo y Regulación Del GlucógenoДокумент6 страницMetabolismo y Regulación Del GlucógenoJose RamirezОценок пока нет
- Cuestionario Carboxilicos y Ruta de Las PentosasДокумент2 страницыCuestionario Carboxilicos y Ruta de Las PentosasAlfonso Lopez PeñafordОценок пока нет
- Solemne 3 BioquimicaДокумент2 страницыSolemne 3 BioquimicaalexОценок пока нет
- Examen Unidad III de Bioquimica GeneralДокумент4 страницыExamen Unidad III de Bioquimica GeneralDiego ChecaОценок пока нет
- Cuestionario ResueltoДокумент13 страницCuestionario ResueltoJose Bastidas0% (1)
- Metabolismo de Otras HexosasДокумент4 страницыMetabolismo de Otras HexosasMaria Zimbrelo Alcarria0% (1)
- Ciclo de Cori: la circulación de glucosa y lactato entre músculo y hígadoДокумент7 страницCiclo de Cori: la circulación de glucosa y lactato entre músculo y hígadoJeanette PaulinaОценок пока нет
- Tarea AminoacidosДокумент3 страницыTarea AminoacidosFelipe MercatelliОценок пока нет
- Bioquimica PreguntasДокумент2 страницыBioquimica PreguntasJesusNavaОценок пока нет
- Gomezniño Adriana Ejerciciosu1 B4aДокумент6 страницGomezniño Adriana Ejerciciosu1 B4aAdriana GomezОценок пока нет
- 1.introduccion Al MetabolismoДокумент9 страниц1.introduccion Al MetabolismoAndres Felipe Prieto AlarconОценок пока нет
- Estrategia Catalítica de La LisozimaДокумент17 страницEstrategia Catalítica de La LisozimaLudwig Omar Julca Salazar100% (4)
- Taller Proteínas 3Документ2 страницыTaller Proteínas 3Juanjo BotiaОценок пока нет
- Enzimas catalizan reacciones acelerando velocidadДокумент64 страницыEnzimas catalizan reacciones acelerando velocidadKaren Cuenca100% (1)
- Rutas de Pentosas FosfatoДокумент19 страницRutas de Pentosas FosfatoAlvaro FloresОценок пока нет
- GalactosaДокумент9 страницGalactosaGUTIÉRREZ SÁNCHEZ OSCARОценок пока нет
- Digestión de proteínas: Acción de enzimas pancreáticas sobre la caseínaДокумент8 страницDigestión de proteínas: Acción de enzimas pancreáticas sobre la caseínaCarlos CC100% (1)
- Metabolismo de NucleótidosДокумент8 страницMetabolismo de NucleótidosAnonymous woR2GSОценок пока нет
- GlucogenolisisДокумент7 страницGlucogenolisisantonioОценок пока нет
- Regulación de La GlucólisisДокумент6 страницRegulación de La GlucólisisLorik90Оценок пока нет
- Bioquimica II (Guia III)Документ9 страницBioquimica II (Guia III)LindaОценок пока нет
- Clase IX - Bioquimica GeneralДокумент102 страницыClase IX - Bioquimica GeneralHiRodriguez MarkОценок пока нет
- PRACTICA 2 Curva de Tolerancia A La GlucosaДокумент6 страницPRACTICA 2 Curva de Tolerancia A La GlucosaalexisОценок пока нет
- Cuestionario 3Документ2 страницыCuestionario 3AlmendraTapia100% (2)
- Discusion de Grupo N 3Документ14 страницDiscusion de Grupo N 3RubenPerez100% (4)
- Amilasa 15Документ3 страницыAmilasa 15Samiri SamiriОценок пока нет
- Relación Entre Ciclo de La Urea y KrebsДокумент1 страницаRelación Entre Ciclo de La Urea y KrebsDavid Vergara67% (3)
- Discusion Nº3Документ22 страницыDiscusion Nº3christian8diaz-59207Оценок пока нет
- Extraccion de Lipidos de Cerebro Brain LДокумент4 страницыExtraccion de Lipidos de Cerebro Brain LJosue AlvarezОценок пока нет
- La Insulina y Sus Efectos MetabólicosДокумент12 страницLa Insulina y Sus Efectos MetabólicosOscar Vasquez CorreaОценок пока нет
- Cuestionario - Tema 10Документ5 страницCuestionario - Tema 10María del Carmen Ramírez SalinasОценок пока нет
- La Enzima Malato DeshidrogenasaДокумент2 страницыLa Enzima Malato DeshidrogenasaKaren Torres JuárezОценок пока нет
- Guia Fisiologia IДокумент151 страницаGuia Fisiologia Ijosmiguel_bv100% (1)
- Act12 de BioquimicaДокумент5 страницAct12 de BioquimicaLucy Judith Cortes MartinezОценок пока нет
- Glucólisis y destinos del piruvatoДокумент14 страницGlucólisis y destinos del piruvatoLuisianaОценок пока нет
- Agar TioglicolatoДокумент2 страницыAgar TioglicolatoAdrianaОценок пока нет
- Etapas Del Ciclo de La UreaДокумент2 страницыEtapas Del Ciclo de La UreaMary CamacaroОценок пока нет
- Guía Práctica 3. Cuantificación de Proteínas Totales y AlbúminaДокумент7 страницGuía Práctica 3. Cuantificación de Proteínas Totales y AlbúminaEnoc MoránОценок пока нет
- Cap 18 Glucolisis y Ox Del Piruvato HARPER Bioquimica Ilustrada 28a Ed-10ArchivoДокумент8 страницCap 18 Glucolisis y Ox Del Piruvato HARPER Bioquimica Ilustrada 28a Ed-10ArchivoAbigail GajardoОценок пока нет
- Determinación bilirrubinaДокумент10 страницDeterminación bilirrubinaJoanie StewartОценок пока нет
- Metabolismo de La GlucosaДокумент15 страницMetabolismo de La GlucosaDayana EstefaniaОценок пока нет
- Tecnicas de Microscopia y Uso Correcto Del Microscopio Binocular CompuestoДокумент4 страницыTecnicas de Microscopia y Uso Correcto Del Microscopio Binocular CompuestoAndrés Felipe MonsalveОценок пока нет
- Lab 2a (2012) - Fisiologia Del Sistema EndocrinoДокумент18 страницLab 2a (2012) - Fisiologia Del Sistema EndocrinoCidAlexanderRami0% (1)
- LDH-40Документ2 страницыLDH-40Patricia Donaires100% (1)
- Medios de cultivo Yersinia y AeromonasДокумент9 страницMedios de cultivo Yersinia y AeromonasFRANCISCO JAVIER CANO CHAVEZОценок пока нет
- Inf4BQ Miercoles10am Ciclo de Krebs GeanCarlos SaavedraValenciaДокумент5 страницInf4BQ Miercoles10am Ciclo de Krebs GeanCarlos SaavedraValenciaJuan Diego CamachoОценок пока нет
- Glucólisis, Gluconeogénesis y Ruta de Las Pentosas FosfatoДокумент26 страницGlucólisis, Gluconeogénesis y Ruta de Las Pentosas FosfatoMarcelo Kim100% (1)
- La GlucólisisДокумент27 страницLa Glucólisisrock2312Оценок пока нет
- Enzimas HepaticasДокумент15 страницEnzimas HepaticasHERNÁNDEZ VALENCIANO KAREN CECILIAОценок пока нет
- Laboratorio de Bioquimica EnzimasДокумент21 страницаLaboratorio de Bioquimica EnzimasAndrea Zambrano TorresОценок пока нет
- Informe de LipidosДокумент2 страницыInforme de LipidosMaria Victoria AriñaОценок пока нет
- Clasificacion de Medios de CultivoДокумент3 страницыClasificacion de Medios de CultivoEdna Hdz50% (2)
- Bioquímica I examen unidadДокумент3 страницыBioquímica I examen unidadRicardo Gustavo Iparraguirre CortavitarteОценок пока нет
- Proteína Visceral y EsqueléticaДокумент7 страницProteína Visceral y EsqueléticaNicole RamirezОценок пока нет
- Informe de Glucogenolisis Uu12Документ24 страницыInforme de Glucogenolisis Uu12dieg0280% (1)
- Tema 17. Metabolismo Del Glucógeno y Su RegulaciónДокумент13 страницTema 17. Metabolismo Del Glucógeno y Su RegulaciónÁngel Parra CominoОценок пока нет
- Metabolismo Del GlucógenoДокумент48 страницMetabolismo Del GlucógenoJoel CruzОценок пока нет
- Procesados FrutasДокумент79 страницProcesados FrutasDANGEROUSCATОценок пока нет
- DS-L01 Lineamientos para La Elaboracion y Control Doc SG - PuДокумент31 страницаDS-L01 Lineamientos para La Elaboracion y Control Doc SG - PurobertcotrinaОценок пока нет
- FileДокумент2 страницыFilerobertcotrinaОценок пока нет
- Dopamina Síntesis, Liberación y Receptores en El Sistema Nervioso Central Bio001f PDFДокумент22 страницыDopamina Síntesis, Liberación y Receptores en El Sistema Nervioso Central Bio001f PDFJoelBen50% (2)
- Jefe de LaboratorioДокумент3 страницыJefe de LaboratoriorobertcotrinaОценок пока нет
- ComasДокумент81 страницаComasRemy SP100% (1)
- GestiÓ N Por Procesos e IndicadoresДокумент5 страницGestiÓ N Por Procesos e IndicadoresErick Joel JesusОценок пока нет
- Planeamiento Y Control Del Sistema de Producción: TemarioДокумент1 страницаPlaneamiento Y Control Del Sistema de Producción: TemarioAnimalitoОценок пока нет
- Catalago de Normas ActualizadasДокумент62 страницыCatalago de Normas ActualizadasrobertcotrinaОценок пока нет
- GC-PR-001 Procedimiento Elaboracion y Control de DocumentosДокумент16 страницGC-PR-001 Procedimiento Elaboracion y Control de DocumentosNaylé LopézОценок пока нет
- Stock de Licores 03Документ6 страницStock de Licores 03robertcotrinaОценок пока нет
- Fermentación VegetalesДокумент10 страницFermentación VegetalesrobertcotrinaОценок пока нет
- ECA-MC-C12 Criterios para La Ev de La Norma 17020 2012 V02 PDFДокумент16 страницECA-MC-C12 Criterios para La Ev de La Norma 17020 2012 V02 PDFrobertcotrinaОценок пока нет
- Imforme Liofilizacion en Los AlimentosДокумент9 страницImforme Liofilizacion en Los AlimentosrobertcotrinaОценок пока нет
- BioquímicaPescadoДокумент36 страницBioquímicaPescadorobertcotrinaОценок пока нет
- Fusion de Grasas y AceitesДокумент9 страницFusion de Grasas y AceitesrobertcotrinaОценок пока нет
- Fritura AlimentosДокумент13 страницFritura Alimentosrobertcotrina100% (1)
- Sensores de PresionДокумент4 страницыSensores de PresionrobertcotrinaОценок пока нет
- Seminario 10Документ42 страницыSeminario 10Aracely Escarleth Aguirre de la CruzОценок пока нет
- Biomoléculas: carbohidratos, lípidos, proteínas y ácidos nucleicosДокумент12 страницBiomoléculas: carbohidratos, lípidos, proteínas y ácidos nucleicosYamila MardonesОценок пока нет
- Epigenética en Obesidad y Diabetes Tipo 2 - Papel de La Nutrición, Limitaciones y Futuras AplicacionesДокумент1 страницаEpigenética en Obesidad y Diabetes Tipo 2 - Papel de La Nutrición, Limitaciones y Futuras AplicacionesTANIA CECILIA VASQUEZ ROMEROОценок пока нет
- Examen Nivelación CélulaДокумент4 страницыExamen Nivelación CélulaRonaldo Castillo PayaresОценок пока нет
- Clase 10Документ34 страницыClase 10Valeria Marcela Gutierrez VelasquezОценок пока нет
- Tarea de Generalidades Enfermeria 2021Документ6 страницTarea de Generalidades Enfermeria 2021JUAN JOSE VARGAS MAMANIОценок пока нет
- Elementos de Análisis Compartimental y Balance Hidroelectrolítico CorregidoДокумент31 страницаElementos de Análisis Compartimental y Balance Hidroelectrolítico CorregidoyuniorОценок пока нет
- Informe Fenilcetonuria Albarran TorresДокумент10 страницInforme Fenilcetonuria Albarran Torreschat gptОценок пока нет
- Las CélulasДокумент3 страницыLas CélulasBenilda mendoza roqueОценок пока нет
- Clase 14 y 15 BBДокумент50 страницClase 14 y 15 BBRitter López JaraОценок пока нет
- Tema 5. Membrana CelularДокумент28 страницTema 5. Membrana CelularMiguel ÁngelОценок пока нет
- Enzimas Del VinoДокумент16 страницEnzimas Del VinoJesus ManuelОценок пока нет
- Organización Del ADNДокумент6 страницOrganización Del ADNMARTHA VARGASОценок пока нет
- 21 - FarmacoquimДокумент9 страниц21 - FarmacoquimMiguel Machaca Flores (QuimioFarma)Оценок пока нет
- Antihipertensivo IecasДокумент7 страницAntihipertensivo IecasVidal AbadОценок пока нет
- Patomolecular 2 PCR PDFДокумент13 страницPatomolecular 2 PCR PDFDalmar RodriguezОценок пока нет
- Tipos de Mutaciones-Genetica 2019a4Документ29 страницTipos de Mutaciones-Genetica 2019a4Ochoa Cuevas SarahíОценок пока нет
- Karol Resumen Clase 13Документ3 страницыKarol Resumen Clase 13Moises Castillo GОценок пока нет
- Relacion Entre Minerales y EnzimasДокумент6 страницRelacion Entre Minerales y EnzimasYenny Suxe SánchezОценок пока нет
- Silabus de Nutricion AnimalДокумент3 страницыSilabus de Nutricion AnimalC J Chagua MachaguaiОценок пока нет
- Guía No.5 Genética Molecular - Arn 2023Документ8 страницGuía No.5 Genética Molecular - Arn 2023Blogamers cracksОценок пока нет
- Actividad 1.5 - Factores Que Determinan La Utilización de Substratos Durante El Ejercicio Físicoadalberto de J Hdz.Документ4 страницыActividad 1.5 - Factores Que Determinan La Utilización de Substratos Durante El Ejercicio Físicoadalberto de J Hdz.hernandezalexa4339Оценок пока нет
- Partes Del Sistema DigestivoДокумент3 страницыPartes Del Sistema DigestivoAdriana RebolledoОценок пока нет
- Trabajo Práctico de Biología Nº1: "Extracción Del ADN Vegetal"Документ7 страницTrabajo Práctico de Biología Nº1: "Extracción Del ADN Vegetal"Jimena CamachoОценок пока нет
- BIOLOGÍA 1 (B) - Biomoléculas Orgánicas - GlúcidosДокумент3 страницыBIOLOGÍA 1 (B) - Biomoléculas Orgánicas - GlúcidosJorge RebazaОценок пока нет
- Propiedades proteínasДокумент6 страницPropiedades proteínasJoicy AlejandroОценок пока нет
- Clasificacion BacterianaДокумент20 страницClasificacion BacterianaBrenda Dalila Barrientos MartinezОценок пока нет
- Quiz 1 - Semana 3 - Ra - Primer Bloque-Biologia Humana - (Grupo2)Документ11 страницQuiz 1 - Semana 3 - Ra - Primer Bloque-Biologia Humana - (Grupo2)ErikaОценок пока нет
- Átomo Molecula Celula OrganismosДокумент9 страницÁtomo Molecula Celula OrganismosJean Tejada TrejoОценок пока нет
- Sistema GastrointestinalДокумент12 страницSistema GastrointestinalMilagro Ester HerreraОценок пока нет