Вам также может понравиться
- DownloadДокумент9 страницDownloadbrkica2011Оценок пока нет
- Thesis Proposal 1Документ11 страницThesis Proposal 1Jhon Mar Eco GataoОценок пока нет
- RSPB 20091076Документ7 страницRSPB 20091076ΧαράλαμποςΧατζηκανέλλουОценок пока нет
- Journal Pone 0025630Документ20 страницJournal Pone 0025630sitvensonОценок пока нет
- Ajol-File-Journals 769 Articles 259606 655c79d6896f0Документ18 страницAjol-File-Journals 769 Articles 259606 655c79d6896f0Efosa OdiaОценок пока нет
- Botryosphaeriaceae Associated With Eucalyptus Canker Diseases in ColombiaДокумент14 страницBotryosphaeriaceae Associated With Eucalyptus Canker Diseases in Colombiachaveslg5755Оценок пока нет
- Practical Manual - Insect Ecology & IPMДокумент81 страницаPractical Manual - Insect Ecology & IPMPrakash Kumar Kumar67% (15)
- Article: LepanthesДокумент12 страницArticle: LepanthesCésar RuedaОценок пока нет
- Grapevine Trunk Diseases: Complex and Still Poorly UnderstoodДокумент23 страницыGrapevine Trunk Diseases: Complex and Still Poorly UnderstoodzuilinhaОценок пока нет
- Plant+Disease++Vol +94+no +8Документ12 страницPlant+Disease++Vol +94+no +8Virgilio UlloaОценок пока нет
- J Funeco 2021 101087Документ12 страницJ Funeco 2021 101087Raymundo Lopez NОценок пока нет
- 2019 ToussaintДокумент12 страниц2019 ToussaintMaristela VolpatoОценок пока нет
- Fungal Ecology and Succession On Phragmites Australis in A Brackish Tidal Marsh. II. StemsДокумент25 страницFungal Ecology and Succession On Phragmites Australis in A Brackish Tidal Marsh. II. StemsDinoMarićОценок пока нет
- Machine Vision - Idenfication of PlantsДокумент21 страницаMachine Vision - Idenfication of PlantsrphmiОценок пока нет
- Vulpia MyurosДокумент5 страницVulpia MyurosOana MikyОценок пока нет
- Parasitological Survey On Mangrove Community in Sitio Bayug, Brgy. Hinaplanon, Iligan CityДокумент12 страницParasitological Survey On Mangrove Community in Sitio Bayug, Brgy. Hinaplanon, Iligan CityJissah Mae Bahan100% (1)
- Disease Progression in Plantations of Acacia Mangium A Ffected by Red Root Rot (Ganoderma Philippii)Документ14 страницDisease Progression in Plantations of Acacia Mangium A Ffected by Red Root Rot (Ganoderma Philippii)rizky rajabillahОценок пока нет
- Phymatotrichum (Cotton) Root Rot Caused by Phymatotrichopsis OmnivoraДокумент10 страницPhymatotrichum (Cotton) Root Rot Caused by Phymatotrichopsis OmnivoraM.C. BRAULIO ALBERTO LEMUS SORIANOОценок пока нет
- Spore Dispersal Patterns of The Ascomycete FungusДокумент14 страницSpore Dispersal Patterns of The Ascomycete Funguslauraa21Оценок пока нет
- A Global Test of The Pollination Syndrome HypothesisДокумент10 страницA Global Test of The Pollination Syndrome HypothesisConstanza Andía AvendañoОценок пока нет
- Ex Hier - PartДокумент12 страницEx Hier - PartastrolopitequusОценок пока нет
- Crop Protection: SciencedirectДокумент13 страницCrop Protection: SciencedirectIlija MileticОценок пока нет
- Understanding The Diversity of Foliar Endophytic Fungi: Progress, Challenges, and FrontiersДокумент16 страницUnderstanding The Diversity of Foliar Endophytic Fungi: Progress, Challenges, and FrontiersNancy HutchinsonОценок пока нет
- The Effect of Urban Ground Covers On Arthropods: An ExperimentДокумент24 страницыThe Effect of Urban Ground Covers On Arthropods: An ExperimentGina Lorena SánchezОценок пока нет
- Thorp2004 - RS Weeds ReviewДокумент32 страницыThorp2004 - RS Weeds ReviewJose G. GuerraОценок пока нет
- Fungi DivesityДокумент17 страницFungi DivesityArgelia CamachoОценок пока нет
- tmpF285 TMPДокумент20 страницtmpF285 TMPFrontiersОценок пока нет
- Bertsch Et Al 2012 Plant PatholДокумент23 страницыBertsch Et Al 2012 Plant PatholJose Miguel Soto HerediaОценок пока нет
- Anatomical Study of Sansevieria Zeylanica Leaf Affected by Vehicular EmissionsДокумент35 страницAnatomical Study of Sansevieria Zeylanica Leaf Affected by Vehicular EmissionsLowie Soriano Bacunot100% (1)
- Tree Inventory Analysis in Văcăreşti Natural ParkДокумент8 страницTree Inventory Analysis in Văcăreşti Natural ParkVladimir BocОценок пока нет
- Patogeni Quercia Da Sughero - Ragazzi Et AlДокумент10 страницPatogeni Quercia Da Sughero - Ragazzi Et AlmncmarcoОценок пока нет
- Memmotetal 2004 Proc RSoc BДокумент7 страницMemmotetal 2004 Proc RSoc BConstanza Andía AvendañoОценок пока нет
- 4855 9357 1 SMДокумент9 страниц4855 9357 1 SMwonpil kimОценок пока нет
- Linking Ecology and Plant PathologyДокумент16 страницLinking Ecology and Plant PathologyGustavo Cano GallegosОценок пока нет
- Mapping of Shoot y Tolerance Loci in Sorghum Using SSR MarkersДокумент8 страницMapping of Shoot y Tolerance Loci in Sorghum Using SSR MarkersBrent LagarizaОценок пока нет
- Air Quality Monitoring System Using Lichens As Bioindicators in Central ArgentinaДокумент9 страницAir Quality Monitoring System Using Lichens As Bioindicators in Central Argentinabryan160892Оценок пока нет
- Bannari 1995Документ28 страницBannari 1995Klissman Morales OlabarreraОценок пока нет
- 2016 IEEE-fruit-Hybrid Approach For Anthracnose Detection usingTIAR.2016.Документ5 страниц2016 IEEE-fruit-Hybrid Approach For Anthracnose Detection usingTIAR.2016.shripadОценок пока нет
- Cinnamomum Verum (Presl.)Документ4 страницыCinnamomum Verum (Presl.)Husnul SiregarОценок пока нет
- The Role of Insect As Vector of Gall Rust Disease On Albizia Falcataria L. FosbergДокумент6 страницThe Role of Insect As Vector of Gall Rust Disease On Albizia Falcataria L. FosbergAhmad Irgan DamaraОценок пока нет
- Aizen y Feinsinger.1994Документ23 страницыAizen y Feinsinger.1994Alexis Centeno CuellarОценок пока нет
- Dicke 2003Документ3 страницыDicke 2003Luca MuratoriОценок пока нет
- Jurnal IngrisДокумент11 страницJurnal IngrisdewiОценок пока нет
- A Simple Model For Nondestructive Leaf Area Estimation in Bedding PlantsДокумент9 страницA Simple Model For Nondestructive Leaf Area Estimation in Bedding PlantsnazrulОценок пока нет
- Interploidy Hybridization in Sympatric Zones The FormationДокумент14 страницInterploidy Hybridization in Sympatric Zones The FormationTeflon SlimОценок пока нет
- Shade Tree Management Affects Fruit Abortion, Insect Pests and Pathogens of CacaoДокумент5 страницShade Tree Management Affects Fruit Abortion, Insect Pests and Pathogens of CacaoArdiyanti Tiwoek PurwaningsihОценок пока нет
- Schizophyllum Commune - The Main Cause: of Dying Trees of The Banja Luka Arbored Walks and ParksДокумент11 страницSchizophyllum Commune - The Main Cause: of Dying Trees of The Banja Luka Arbored Walks and ParksMiroljub KrsticОценок пока нет
- Conservation Ecology - Ups and Downs in Pollinator Populations - When Is There A DeclineДокумент15 страницConservation Ecology - Ups and Downs in Pollinator Populations - When Is There A DeclineSal BorОценок пока нет
- 2018 - Stürmer Et AlДокумент17 страниц2018 - Stürmer Et AlMaristela VolpatoОценок пока нет
- Forests 09 00732Документ9 страницForests 09 00732Eduardo ArroyoОценок пока нет
- Sub Angui en Pasto GolfДокумент6 страницSub Angui en Pasto GolfEdgar Medina GomezОценок пока нет
- Discussion Adult SeedelingДокумент13 страницDiscussion Adult SeedelingAlejandra LlerenaОценок пока нет
- GJESM Volume 7 Issue 3 Pages 473-484Документ12 страницGJESM Volume 7 Issue 3 Pages 473-484GJESMОценок пока нет
- Corn Stalk Borer Infestation On SorghumДокумент3 страницыCorn Stalk Borer Infestation On Sorghumpaspartu121Оценок пока нет
- Int J Appl Earth Obs Geoinformation: SciencedirectДокумент10 страницInt J Appl Earth Obs Geoinformation: SciencedirectMichael WondemuОценок пока нет
- Conl 12674Документ8 страницConl 12674Manuel PérezОценок пока нет
- Influence of Mulching On Virus Disease IДокумент8 страницInfluence of Mulching On Virus Disease ISandra MadukaifeОценок пока нет
- Gauzere Et Al Mating 2013Документ15 страницGauzere Et Al Mating 2013cubanosОценок пока нет
- Scientia 2007Документ6 страницScientia 2007Liv CBОценок пока нет
- 1Документ8 страниц110sgОценок пока нет
- NIH Public Access: p21 Protects "Super p53" Mice From The Radiation-Induced Gastrointestinal SyndromeДокумент7 страницNIH Public Access: p21 Protects "Super p53" Mice From The Radiation-Induced Gastrointestinal Syndrome10sgОценок пока нет
- Sex-Dependent Novelty Response in Neurexin-1 A Mutant MiceДокумент6 страницSex-Dependent Novelty Response in Neurexin-1 A Mutant Mice10sgОценок пока нет
- A Research Agenda For Helminth Diseases of Humans: Modelling For Control and EliminationДокумент11 страницA Research Agenda For Helminth Diseases of Humans: Modelling For Control and Elimination10sgОценок пока нет
- NIH Public Access: Author ManuscriptДокумент16 страницNIH Public Access: Author Manuscript10sgОценок пока нет
- Is Axis Position Within Tree Architecture A Determinant of Axis Morphology, Branching, Flowering and Fruiting? An Essay in MangoДокумент12 страницIs Axis Position Within Tree Architecture A Determinant of Axis Morphology, Branching, Flowering and Fruiting? An Essay in Mango10sgОценок пока нет
- Field Attractants For Pachnoda Interrupta Selected by Means of GC-EAD and Single Sensillum ScreeningДокумент14 страницField Attractants For Pachnoda Interrupta Selected by Means of GC-EAD and Single Sensillum Screening10sgОценок пока нет
- Marine Drugs: Immense Essence of Excellence: Marine Microbial Bioactive CompoundsДокумент29 страницMarine Drugs: Immense Essence of Excellence: Marine Microbial Bioactive Compounds10sgОценок пока нет
- Role of Oxidative Stress and The Activity of Ethylene Biosynthetic Enzymes On The Formation of Spongy Tissue in Alphonso' MangoДокумент5 страницRole of Oxidative Stress and The Activity of Ethylene Biosynthetic Enzymes On The Formation of Spongy Tissue in Alphonso' Mango10sgОценок пока нет
- Allergy: Hypersensitivity Manifestations To The Fruit MangoДокумент7 страницAllergy: Hypersensitivity Manifestations To The Fruit Mango10sgОценок пока нет
- Activation of Ethylene-Responsive P-Hydroxyphenylpyruvate Dioxygenase Leads To Increased Tocopherol Levels During Ripening in MangoДокумент11 страницActivation of Ethylene-Responsive P-Hydroxyphenylpyruvate Dioxygenase Leads To Increased Tocopherol Levels During Ripening in Mango10sgОценок пока нет
- Optimization of Serine Protease Purification From Mango (Mangifera Indica Cv. Chokanan) Peel in Polyethylene Glycol/Dextran Aqueous Two Phase SystemДокумент14 страницOptimization of Serine Protease Purification From Mango (Mangifera Indica Cv. Chokanan) Peel in Polyethylene Glycol/Dextran Aqueous Two Phase System10sgОценок пока нет
- Contrasting Diversity Patterns of Crenarchaeal, Bacterial and Fungal Soil Communities in An Alpine LandscapeДокумент7 страницContrasting Diversity Patterns of Crenarchaeal, Bacterial and Fungal Soil Communities in An Alpine Landscape10sgОценок пока нет
- Limits of Nuclear Ribosomal DNA Internal Transcribed Spacer (ITS) Sequences As Species Barcodes For FungiДокумент1 страницаLimits of Nuclear Ribosomal DNA Internal Transcribed Spacer (ITS) Sequences As Species Barcodes For Fungi10sgОценок пока нет
- Shifts in Soil Microorganisms in Response To Warming Are Consistent Across A Range of Antarctic EnvironmentsДокумент11 страницShifts in Soil Microorganisms in Response To Warming Are Consistent Across A Range of Antarctic Environments10sgОценок пока нет
- Validation and Application of A PCR Primer Set To Quantify Fungal Communities in The Soil Environment by Real-Time Quantitative PCRДокумент13 страницValidation and Application of A PCR Primer Set To Quantify Fungal Communities in The Soil Environment by Real-Time Quantitative PCR10sgОценок пока нет
- The History, Fungal Biodiversity, Conservation, and Future Perspectives For Mycology in EgyptДокумент20 страницThe History, Fungal Biodiversity, Conservation, and Future Perspectives For Mycology in Egypt10sgОценок пока нет
- Soil Eukaryotic Microorganism Succession As Affected by Continuous Cropping of Peanut - Pathogenic and Beneficial Fungi Were SelectedДокумент9 страницSoil Eukaryotic Microorganism Succession As Affected by Continuous Cropping of Peanut - Pathogenic and Beneficial Fungi Were Selected10sgОценок пока нет
- Evolution and Organization of A Highly Dynamic, Subtelomeric Helicase Gene Family in The Rice Blast Fungus Magnaporthe GriseaДокумент10 страницEvolution and Organization of A Highly Dynamic, Subtelomeric Helicase Gene Family in The Rice Blast Fungus Magnaporthe Grisea10sgОценок пока нет
- New Strategy For Rapid Diagnosis and Characterization of Fungal Infections: The Example of Corneal ScrapingsДокумент10 страницNew Strategy For Rapid Diagnosis and Characterization of Fungal Infections: The Example of Corneal Scrapings10sgОценок пока нет
- Special Issue Paper: Olorode Afr. J. Trad. CAM (2007) 4 (4) : 532 - 540Документ9 страницSpecial Issue Paper: Olorode Afr. J. Trad. CAM (2007) 4 (4) : 532 - 54010sgОценок пока нет
- Diazotrophicus On Solid Media To Changes in AtmosphericДокумент7 страницDiazotrophicus On Solid Media To Changes in Atmospheric10sgОценок пока нет
- Relationships of Cereal Crops and Other Grasses: Vol. 95, Pp. 2005-2010, March 1998 Colloquium PaperДокумент6 страницRelationships of Cereal Crops and Other Grasses: Vol. 95, Pp. 2005-2010, March 1998 Colloquium Paper10sgОценок пока нет
- Yukot,+houkelin 2505 11892735 Final+Paper+Group+41Документ17 страницYukot,+houkelin 2505 11892735 Final+Paper+Group+410191720003 ELIAS ANTONIO BELLO LEON ESTUDIANTE ACTIVOОценок пока нет
- AdverbsДокумент10 страницAdverbsKarina Ponce RiosОценок пока нет
- Chapter 12 Social Structural Theories of CrimeДокумент5 страницChapter 12 Social Structural Theories of CrimeKaroline Thomas100% (1)
- Marketing Mix of Van HeusenДокумент38 страницMarketing Mix of Van HeusenShail PatelОценок пока нет
- Final Presentation BANK OF BARODA 1Документ8 страницFinal Presentation BANK OF BARODA 1Pooja GoyalОценок пока нет
- NCPДокумент3 страницыNCPchesca_paunganОценок пока нет
- Bakhtin's Chronotope On The RoadДокумент17 страницBakhtin's Chronotope On The RoadLeandro OliveiraОценок пока нет
- KAHOOT - Assignment 4.1 Lesson PlanДокумент3 страницыKAHOOT - Assignment 4.1 Lesson PlanJan ZimmermannОценок пока нет
- E TN SWD Csa A23 3 94 009 PDFДокумент5 страницE TN SWD Csa A23 3 94 009 PDFRazvan RobertОценок пока нет
- War Thesis StatementsДокумент8 страницWar Thesis StatementsHelpPaperRochester100% (2)
- Contents Serbo-Croatian GrammarДокумент2 страницыContents Serbo-Croatian GrammarLeo VasilaОценок пока нет
- List de VerbosДокумент2 страницыList de VerbosmarcoОценок пока нет
- City/ The Countryside: VocabularyДокумент2 страницыCity/ The Countryside: VocabularyHương Phạm QuỳnhОценок пока нет
- 28Документ12 страниц28physicsdocsОценок пока нет
- Playwriting Pedagogy and The Myth of IntrinsicДокумент17 страницPlaywriting Pedagogy and The Myth of IntrinsicCaetano BarsoteliОценок пока нет
- Zero Power Factor Method or Potier MethodДокумент1 страницаZero Power Factor Method or Potier MethodMarkAlumbroTrangiaОценок пока нет
- Case Study 1 (Pneumonia)Документ13 страницCase Study 1 (Pneumonia)Kate EscotonОценок пока нет
- In Mein KampfДокумент3 страницыIn Mein KampfAnonymous t5XUqBОценок пока нет
- MasterMind 1 Unit 5 Extra LifeSkills Lesson 2Документ2 страницыMasterMind 1 Unit 5 Extra LifeSkills Lesson 2Hugo A FEОценок пока нет
- Approach To Vaginal Discharge in ChildrenДокумент12 страницApproach To Vaginal Discharge in ChildrensujataОценок пока нет
- Lista Verbelor Regulate - EnglezaДокумент5 страницLista Verbelor Regulate - Englezaflopalan100% (1)
- Simple Future Tense & Future Continuous TenseДокумент2 страницыSimple Future Tense & Future Continuous TenseFarris Ab RashidОценок пока нет
- Star QuizДокумент3 страницыStar Quizapi-254428474Оценок пока нет
- GAJ Mod 18 Ace Your InterviewДокумент12 страницGAJ Mod 18 Ace Your InterviewAnjela SantiagoОценок пока нет
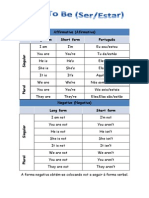
- Affirmative (Afirmativa) Long Form Short Form PortuguêsДокумент3 страницыAffirmative (Afirmativa) Long Form Short Form PortuguêsAnitaYangОценок пока нет
- #6 Decision Control InstructionДокумент9 страниц#6 Decision Control InstructionTimothy King LincolnОценок пока нет
- MCN Drill AnswersДокумент12 страницMCN Drill AnswersHerne Balberde100% (1)
- SUBSET-026-7 v230 - 060224Документ62 страницыSUBSET-026-7 v230 - 060224David WoodhouseОценок пока нет
- Infusion Site Selection and Infusion Set ChangeДокумент8 страницInfusion Site Selection and Infusion Set ChangegaridanОценок пока нет
- "What Is A Concept Map?" by (Novak & Cañas, 2008)Документ4 страницы"What Is A Concept Map?" by (Novak & Cañas, 2008)WanieОценок пока нет
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessОт Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessРейтинг: 4 из 5 звезд4/5 (33)
- Why We Die: The New Science of Aging and the Quest for ImmortalityОт EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityРейтинг: 4.5 из 5 звезд4.5/5 (6)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisОт EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisРейтинг: 3.5 из 5 звезд3.5/5 (2)
- Tales from Both Sides of the Brain: A Life in NeuroscienceОт EverandTales from Both Sides of the Brain: A Life in NeuroscienceРейтинг: 3 из 5 звезд3/5 (18)
- The Rise and Fall of the Dinosaurs: A New History of a Lost WorldОт EverandThe Rise and Fall of the Dinosaurs: A New History of a Lost WorldРейтинг: 4 из 5 звезд4/5 (597)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomОт EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomРейтинг: 4 из 5 звезд4/5 (216)
- Return of the God Hypothesis: Three Scientific Discoveries That Reveal the Mind Behind the UniverseОт EverandReturn of the God Hypothesis: Three Scientific Discoveries That Reveal the Mind Behind the UniverseРейтинг: 4.5 из 5 звезд4.5/5 (52)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsОт EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsРейтинг: 4.5 из 5 звезд4.5/5 (6)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceОт EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceРейтинг: 4.5 из 5 звезд4.5/5 (517)
- Gut: the new and revised Sunday Times bestsellerОт EverandGut: the new and revised Sunday Times bestsellerРейтинг: 4 из 5 звезд4/5 (393)
- Seven and a Half Lessons About the BrainОт EverandSeven and a Half Lessons About the BrainРейтинг: 4 из 5 звезд4/5 (111)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedОт EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedРейтинг: 4 из 5 звезд4/5 (11)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesОт EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesРейтинг: 4.5 из 5 звезд4.5/5 (397)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionОт EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionРейтинг: 4 из 5 звезд4/5 (812)
- Who's in Charge?: Free Will and the Science of the BrainОт EverandWho's in Charge?: Free Will and the Science of the BrainРейтинг: 4 из 5 звезд4/5 (65)
- Good Without God: What a Billion Nonreligious People Do BelieveОт EverandGood Without God: What a Billion Nonreligious People Do BelieveРейтинг: 4 из 5 звезд4/5 (66)
- Moral Tribes: Emotion, Reason, and the Gap Between Us and ThemОт EverandMoral Tribes: Emotion, Reason, and the Gap Between Us and ThemРейтинг: 4.5 из 5 звезд4.5/5 (116)
- Remnants of Ancient Life: The New Science of Old FossilsОт EverandRemnants of Ancient Life: The New Science of Old FossilsРейтинг: 3 из 5 звезд3/5 (3)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildОт EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildРейтинг: 4.5 из 5 звезд4.5/5 (44)
- Darwin's Doubt: The Explosive Origin of Animal Life and the Case for Intelligent DesignОт EverandDarwin's Doubt: The Explosive Origin of Animal Life and the Case for Intelligent DesignРейтинг: 4 из 5 звезд4/5 (19)
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindОт EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindРейтинг: 4.5 из 5 звезд4.5/5 (93)
- Human: The Science Behind What Makes Your Brain UniqueОт EverandHuman: The Science Behind What Makes Your Brain UniqueРейтинг: 3.5 из 5 звезд3.5/5 (38)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorОт EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorОценок пока нет