Вам также может понравиться
- J. Biol. Chem.-1992-Sadoshima-10551-60Документ10 страницJ. Biol. Chem.-1992-Sadoshima-10551-60Yusa ResaОценок пока нет
- Creatina - Suplementacao TerapeuticaДокумент14 страницCreatina - Suplementacao TerapeuticaMagno FilhoОценок пока нет
- Mecanismos Moleculares de Adaptacion Cardiaca Al Ejercicio 2017Документ29 страницMecanismos Moleculares de Adaptacion Cardiaca Al Ejercicio 2017Christian JaraОценок пока нет
- Ds RNA1Документ10 страницDs RNA1bharatkapoorОценок пока нет
- Moghadam2020 Article TheEffectsOfConcurrentTrainingДокумент9 страницMoghadam2020 Article TheEffectsOfConcurrentTrainingamirreza jmОценок пока нет
- Exercise Physiology for the Pediatric and Congenital CardiologistОт EverandExercise Physiology for the Pediatric and Congenital CardiologistОценок пока нет
- 1 s2.0 S155041311630225X MainДокумент14 страниц1 s2.0 S155041311630225X Mainadi pranotoОценок пока нет
- Review Journal Experimental Model of Myocardial Infarction Induced by Isoproterenol in RatsДокумент5 страницReview Journal Experimental Model of Myocardial Infarction Induced by Isoproterenol in RatsM.Rizky irawanОценок пока нет
- 2019 - Myofibrillar Protein Synthesis and MuscleДокумент10 страниц2019 - Myofibrillar Protein Synthesis and MuscleDiego Duarte CavalcanteОценок пока нет
- Adipogenesis: Kelesha Sarjeant and Jacqueline M. StephensДокумент20 страницAdipogenesis: Kelesha Sarjeant and Jacqueline M. Stephenswhitedove31Оценок пока нет
- Villeda, 2011 PDFДокумент7 страницVilleda, 2011 PDFDaniel TorresОценок пока нет
- An Evidence Based Narrative Review of Mechanisms.16Документ14 страницAn Evidence Based Narrative Review of Mechanisms.16Philippus Von HohenheimОценок пока нет
- Muscle Damage LTEДокумент6 страницMuscle Damage LTEAnna PylypchukОценок пока нет
- Stem Cell ReportsДокумент13 страницStem Cell ReportsLuisa FernandaОценок пока нет
- Cells: Molecular Mechanisms of Cardiac Remodeling and Regeneration in Physical ExerciseДокумент17 страницCells: Molecular Mechanisms of Cardiac Remodeling and Regeneration in Physical ExerciseCristian OyarzoОценок пока нет
- Impacto Del Ejercicio FisicoДокумент27 страницImpacto Del Ejercicio FisicoOscarDavidGordilloGonzalezОценок пока нет
- Blood Flow Restriction During Low-Intensity ResistДокумент11 страницBlood Flow Restriction During Low-Intensity ResistKatrina KimОценок пока нет
- Senescence Suppresses The Integrated Stress Response and Activates A Stress-Enhanced Secretory PhenotypeДокумент29 страницSenescence Suppresses The Integrated Stress Response and Activates A Stress-Enhanced Secretory Phenotypenicocarro1995Оценок пока нет
- Cardiac HypertrophyДокумент9 страницCardiac HypertrophyJulia WindiОценок пока нет
- Exercise and Cardiac Health Physiological and Molecular InsightsДокумент11 страницExercise and Cardiac Health Physiological and Molecular InsightsAngélica MuñozОценок пока нет
- Advances in Exercise, Fitness, and Performance.2Документ9 страницAdvances in Exercise, Fitness, and Performance.2Francisco Júlio de SousaОценок пока нет
- Pasti FixДокумент17 страницPasti FixMuhammad LutfiОценок пока нет
- Psychological Stress During Exercise Lymphocyte Subset Redistribution in FirefightersДокумент7 страницPsychological Stress During Exercise Lymphocyte Subset Redistribution in FirefightersFlorentina SisuОценок пока нет
- NIH Public AccessДокумент26 страницNIH Public AccessEmy Noviana SandyОценок пока нет
- Resistance Exercise-Induced Regulation of Muscle Protein Synthesis To Intraset RestДокумент35 страницResistance Exercise-Induced Regulation of Muscle Protein Synthesis To Intraset RestMacarena Beatriz Artigas AriasОценок пока нет
- Scandinavian Med Sci Sports - 2022 - Angleri - Resistance Training Variable Manipulations Is Less Relevant Than IntrinsicДокумент26 страницScandinavian Med Sci Sports - 2022 - Angleri - Resistance Training Variable Manipulations Is Less Relevant Than IntrinsicDaniloОценок пока нет
- Sanchis Gomar2013Документ2 страницыSanchis Gomar2013Ariadna BarretoОценок пока нет
- Ni Hms 601547Документ29 страницNi Hms 601547fidaОценок пока нет
- Generation of Functional Brown Adipocytes From Human Pluripoten - 2020 - Cell STДокумент26 страницGeneration of Functional Brown Adipocytes From Human Pluripoten - 2020 - Cell STBandar Ternama XBОценок пока нет
- 2015 Understanding The Cellular and Molecular Mechanisms of Physical Activity-Induced Health BenefitsДокумент8 страниц2015 Understanding The Cellular and Molecular Mechanisms of Physical Activity-Induced Health Benefitsteja aryudhaОценок пока нет
- Osteoporosis Literature ReviewДокумент5 страницOsteoporosis Literature Reviewafmzinuvouzeny100% (1)
- Skeletal Muscle Protein Synthesis and mTORC1 Nutritional and Contractile Regulation of HumanДокумент12 страницSkeletal Muscle Protein Synthesis and mTORC1 Nutritional and Contractile Regulation of HumankalikОценок пока нет
- Marco Sandri: This Article CitesДокумент12 страницMarco Sandri: This Article CitesjackmstrОценок пока нет
- Mechanisms Behind Blood Flow Restricted Training and Its Effect Towards Muscle GrowthДокумент40 страницMechanisms Behind Blood Flow Restricted Training and Its Effect Towards Muscle GrowthFranciscoJavierRoblesОценок пока нет
- The Effects of Aerobic and Anaerobic Exercises On Circulating Soluble Klotho and Igf I in Young and Elderly Adults and in Cad PatientsДокумент8 страницThe Effects of Aerobic and Anaerobic Exercises On Circulating Soluble Klotho and Igf I in Young and Elderly Adults and in Cad PatientsPJ DSОценок пока нет
- Targeted Ablation of TRAF6 Inhibits Skeletal Muscle Wasting in MiceДокумент17 страницTargeted Ablation of TRAF6 Inhibits Skeletal Muscle Wasting in MiceDiana DangОценок пока нет
- Melo2018 CV Adaptation by RTДокумент8 страницMelo2018 CV Adaptation by RTMumtaz MaulanaОценок пока нет
- Endothelial Cell CenescenceДокумент6 страницEndothelial Cell CenescenceMumtaz MaulanaОценок пока нет
- Associations Between Borgs' Rating of Perceived Exertion and Physiological Measures of Exercise Intensity # ControlДокумент9 страницAssociations Between Borgs' Rating of Perceived Exertion and Physiological Measures of Exercise Intensity # ControlVinicius Fallux QueirozОценок пока нет
- 1985-Cardiac Adaptations To Chronic ExerciseДокумент28 страниц1985-Cardiac Adaptations To Chronic ExerciseJavier Mora BaizОценок пока нет
- Cross-Talk Between Gut and Brain Elicited by Physical ExerciseДокумент19 страницCross-Talk Between Gut and Brain Elicited by Physical ExercisecontactoviphyaОценок пока нет
- almeida2022CRF As A Mediator in The Relation Between Lung Function and Blood PressureДокумент10 страницalmeida2022CRF As A Mediator in The Relation Between Lung Function and Blood PressureMumtaz MaulanaОценок пока нет
- The Endocrine Role of The Skeleton: Background and Clinical EvidenceДокумент9 страницThe Endocrine Role of The Skeleton: Background and Clinical EvidenceRuth BritoОценок пока нет
- Undulating Changes in Human Plasma Proteome Profiles Across The LifespanДокумент22 страницыUndulating Changes in Human Plasma Proteome Profiles Across The LifespanЗлойБобрОценок пока нет
- Antibody Appears To Attack Cancer Cells, Leaving Other Cells UnscathedДокумент13 страницAntibody Appears To Attack Cancer Cells, Leaving Other Cells UnscathedAnonymous EdgflnjcОценок пока нет
- Repeated and Interrupted Resistance Exercise Induces The Desensitization and Re-Sensitization of mTOR-Related Signaling in Human Skeletal Muscle FibersДокумент18 страницRepeated and Interrupted Resistance Exercise Induces The Desensitization and Re-Sensitization of mTOR-Related Signaling in Human Skeletal Muscle FibersHugo FalquetoОценок пока нет
- Cardioprotective role of GTS-21 by attenuating the TLR4/NF-κB pathway in streptozotocin-induced diabetic cardiomyopathy in ratsДокумент21 страницаCardioprotective role of GTS-21 by attenuating the TLR4/NF-κB pathway in streptozotocin-induced diabetic cardiomyopathy in ratsvtrang0310Оценок пока нет
- The Effects of High-Intensity Resistance Exercise On The Blood Lipid Profile and Liver Function in Hypercholesterolemic HamstersДокумент7 страницThe Effects of High-Intensity Resistance Exercise On The Blood Lipid Profile and Liver Function in Hypercholesterolemic HamstersRizki NovitasariОценок пока нет
- NIH Public Access: Author ManuscriptДокумент16 страницNIH Public Access: Author ManuscriptFernando De Oliveira SolimanОценок пока нет
- Alternate Day FastingДокумент35 страницAlternate Day FastingJohnny RockermeierОценок пока нет
- Endothelial Progenitor Cells As A New AgentДокумент14 страницEndothelial Progenitor Cells As A New Agentrajeshkundapur123Оценок пока нет
- Debabrata JTM 2013Документ13 страницDebabrata JTM 2013Tarak Nath KhatuaОценок пока нет
- De Paula 2017Документ21 страницаDe Paula 2017TAINAH DE PAULAОценок пока нет
- 742 FullДокумент8 страниц742 FulljoaoevandroОценок пока нет
- Japplphysiol 00277 2018Документ11 страницJapplphysiol 00277 2018Misael VargasОценок пока нет
- BloodДокумент5 страницBloodIstván PortörőОценок пока нет
- mTOR Signaling Pathway and Myostatin in Men Resistance Exercise With Whey Protein Ingestion AffectsДокумент11 страницmTOR Signaling Pathway and Myostatin in Men Resistance Exercise With Whey Protein Ingestion AffectsMarcosViníciusSilvaОценок пока нет
- Articulo 1 TecДокумент16 страницArticulo 1 TecKristina TabaresОценок пока нет
- J. Nutr.-2016-Reidy-jn.114.203208Документ29 страницJ. Nutr.-2016-Reidy-jn.114.203208Nanci GuestОценок пока нет
- Zhao' 07Документ11 страницZhao' 07Javier Cerda InfanteОценок пока нет
- Cambridge University Press, International Labor and Working-Class, Inc. International Labor and Working-Class HistoryДокумент13 страницCambridge University Press, International Labor and Working-Class, Inc. International Labor and Working-Class HistoryDaniel Lee Eisenberg Jacobs100% (1)
- Claudia Jones Nuclear TestingДокумент25 страницClaudia Jones Nuclear TestingDaniel Lee Eisenberg JacobsОценок пока нет
- Theophilus Capital Against: Fisk Labor1Документ9 страницTheophilus Capital Against: Fisk Labor1Daniel Lee Eisenberg Jacobs100% (1)
- Why The Euro Will Rival The Dollar PDFДокумент25 страницWhy The Euro Will Rival The Dollar PDFDaniel Lee Eisenberg JacobsОценок пока нет
- Browder - The American SpiritДокумент2 страницыBrowder - The American SpiritDaniel Lee Eisenberg JacobsОценок пока нет
- Beta Anomaly An Ex-Ante Tail RiskДокумент104 страницыBeta Anomaly An Ex-Ante Tail RiskDaniel Lee Eisenberg JacobsОценок пока нет
- Karl Kautsky Republic and Social Democra PDFДокумент4 страницыKarl Kautsky Republic and Social Democra PDFDaniel Lee Eisenberg JacobsОценок пока нет
- Conference Group For Central European History of The American Historical AssociationДокумент9 страницConference Group For Central European History of The American Historical AssociationDaniel Lee Eisenberg JacobsОценок пока нет
- Necessary and Sufficient Conditions For Dynamic OptimizationДокумент18 страницNecessary and Sufficient Conditions For Dynamic OptimizationDaniel Lee Eisenberg JacobsОценок пока нет
- 1996 ElectionДокумент84 страницы1996 ElectionDaniel Lee Eisenberg JacobsОценок пока нет
- Continuity Change State of Process of Task ofДокумент1 страницаContinuity Change State of Process of Task ofDaniel Lee Eisenberg JacobsОценок пока нет
- Morningstar GammaДокумент4 страницыMorningstar GammaDaniel Lee Eisenberg JacobsОценок пока нет
- Kant Conflict of FacultiesДокумент11 страницKant Conflict of FacultiesDaniel Lee Eisenberg JacobsОценок пока нет
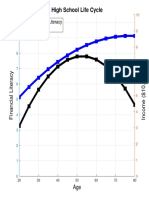
- High School Life Cycle: Financial Literacy IncomeДокумент1 страницаHigh School Life Cycle: Financial Literacy IncomeDaniel Lee Eisenberg JacobsОценок пока нет
- Incentive Contracts in Delegated Portfolio ManagementДокумент41 страницаIncentive Contracts in Delegated Portfolio ManagementDaniel Lee Eisenberg JacobsОценок пока нет
- Statics and Dynamics in Socialist Economics PDFДокумент19 страницStatics and Dynamics in Socialist Economics PDFDaniel Lee Eisenberg JacobsОценок пока нет
- 1st International: Djacobs November 2020Документ5 страниц1st International: Djacobs November 2020Daniel Lee Eisenberg JacobsОценок пока нет
- Long Term Care Insurance PuzzleДокумент17 страницLong Term Care Insurance PuzzleDaniel Lee Eisenberg JacobsОценок пока нет
- University of Oxford Draft-COVID-19-Model - 2020Документ7 страницUniversity of Oxford Draft-COVID-19-Model - 2020Eric L. VanDussenОценок пока нет
- Notes On Forbidden RegressionsДокумент5 страницNotes On Forbidden RegressionsZundaОценок пока нет
- Time Is Money - Rational Life Cycle InertiaДокумент39 страницTime Is Money - Rational Life Cycle InertiaDaniel Lee Eisenberg JacobsОценок пока нет
- Vanguard How Investors Select Advisors PDFДокумент49 страницVanguard How Investors Select Advisors PDFDaniel Lee Eisenberg JacobsОценок пока нет
- Vanguard How Investors Select Advisors PDFДокумент49 страницVanguard How Investors Select Advisors PDFDaniel Lee Eisenberg JacobsОценок пока нет
- Borel Sets PDFДокумент181 страницаBorel Sets PDFDaniel Lee Eisenberg Jacobs100% (1)
- Individualization of Robo-AdviceДокумент8 страницIndividualization of Robo-AdviceDaniel Lee Eisenberg JacobsОценок пока нет
- Cover FERCДокумент1 страницаCover FERCDaniel Lee Eisenberg JacobsОценок пока нет
- A Multiobjective Model For Passive Portfolio ManagementДокумент32 страницыA Multiobjective Model For Passive Portfolio ManagementDaniel Lee Eisenberg JacobsОценок пока нет
- Gpebook PDFДокумент332 страницыGpebook PDFDaniel Lee Eisenberg JacobsОценок пока нет
- 09 MihatovДокумент14 страниц09 MihatovRaphael T. SprengerОценок пока нет
- Genetic Algorithm in Economics and Agent Based ModelsДокумент23 страницыGenetic Algorithm in Economics and Agent Based ModelsDaniel Lee Eisenberg JacobsОценок пока нет
- Flax and Flaxseed Oil: An Ancient Medicine and Modern Functional FoodДокумент21 страницаFlax and Flaxseed Oil: An Ancient Medicine and Modern Functional FoodAnkit Goyal100% (1)
- Physiology of Growth: Arlan L. RosenbloomДокумент12 страницPhysiology of Growth: Arlan L. Rosenbloomانجمن بدنسازی ایرانОценок пока нет
- Dietary Glycemic Factors, Insulin Resistance, and Adiponectin Levels in Acne Vulgaris PDFДокумент8 страницDietary Glycemic Factors, Insulin Resistance, and Adiponectin Levels in Acne Vulgaris PDFCayadi Sidarta Antonius PocipОценок пока нет
- Marine Corps Institute: Semper Fit Advanced Fitness CourseДокумент222 страницыMarine Corps Institute: Semper Fit Advanced Fitness CoursesrollinsОценок пока нет
- Physiology of The Endocrine and Nervous Systems 2017Документ169 страницPhysiology of The Endocrine and Nervous Systems 2017Criss CristinaОценок пока нет
- Acanthosis Nigricans An Extensive ReviewДокумент16 страницAcanthosis Nigricans An Extensive ReviewFebri Dwi HaryonoОценок пока нет
- Dr. Ujjwala Verma 2 Yr JRДокумент60 страницDr. Ujjwala Verma 2 Yr JRUjjwala Verma100% (1)
- Chapter 60 - Insulin, Oral Hypoglycaemic Agents, and The Pharmacology of The Endocrine PancreasДокумент17 страницChapter 60 - Insulin, Oral Hypoglycaemic Agents, and The Pharmacology of The Endocrine PancreasGiliana P. García AcevedoОценок пока нет
- Siim Land - Metabolic Autophagy - Practice Intermittent Fasting and Resistance Training To Build Muscle and Promote Longevity-Independently Published (2019)Документ629 страницSiim Land - Metabolic Autophagy - Practice Intermittent Fasting and Resistance Training To Build Muscle and Promote Longevity-Independently Published (2019)Fernando Martinez Personal Trainer92% (13)
- Endocrine SystemДокумент27 страницEndocrine SystemAlen Vukosavljevic100% (2)
- Fertility in CowДокумент11 страницFertility in CowRicardo QuiñonezОценок пока нет
- Observed Impacts of Insulin Therapy On Callus Cell Transforming Growth Factor-Beta 1 Expression in Diabetic RatsДокумент9 страницObserved Impacts of Insulin Therapy On Callus Cell Transforming Growth Factor-Beta 1 Expression in Diabetic RatsFifih AlamwiyahОценок пока нет
- Insulin and Insulin ResistanceДокумент21 страницаInsulin and Insulin ResistanceFisiologiaUFCG100% (1)
- Targeting Ageing and Preventing Organ Degeneration With MetforminДокумент7 страницTargeting Ageing and Preventing Organ Degeneration With Metforminannisa habibullohОценок пока нет
- Managing Gastrointestinal Complications of Diabetes: Joseph Sellin EditorДокумент133 страницыManaging Gastrointestinal Complications of Diabetes: Joseph Sellin EditorRuth Desiyana GultomОценок пока нет
- Neamtu Marius Cristian RJME 58 3 2017Документ5 страницNeamtu Marius Cristian RJME 58 3 2017Boldeanu MVОценок пока нет
- IPT, Vitamin C and QOLДокумент122 страницыIPT, Vitamin C and QOLBradford S. Weeks100% (1)
- Defic I Enc I A MultipleДокумент16 страницDefic I Enc I A MultipleEnrique GonzalvusОценок пока нет
- Priming Estrogen PDFДокумент5 страницPriming Estrogen PDFgongutzОценок пока нет
- Insulin-Like Growth Factor-I (IGF-I) and Clinical Nutrition: Callum LIVINGSTONEДокумент16 страницInsulin-Like Growth Factor-I (IGF-I) and Clinical Nutrition: Callum LIVINGSTONEYafanita IzzatiОценок пока нет
- Brodsky 2004Документ14 страницBrodsky 2004Ervan SuryaОценок пока нет
- A Unifying Neuro-Fasciagenic Model of Somatic Dysfunction - Underlying Mechanisms and Treatment - PART IДокумент43 страницыA Unifying Neuro-Fasciagenic Model of Somatic Dysfunction - Underlying Mechanisms and Treatment - PART IMarcus Dos SantosОценок пока нет
- Tablas Variabilidad Biológica Actualizadas 95% Confianza 2019Документ10 страницTablas Variabilidad Biológica Actualizadas 95% Confianza 2019Sonrisa Posada RiañoОценок пока нет
- Effects of An Oral Mixture Containing Glycine, Glutamine and Niacin On Memory, GH and IGF-I SecretioДокумент8 страницEffects of An Oral Mixture Containing Glycine, Glutamine and Niacin On Memory, GH and IGF-I SecretioCalin VОценок пока нет
- Fine Test ELISA KitsДокумент376 страницFine Test ELISA KitsDavid1002Оценок пока нет
- Clear For LifeДокумент181 страницаClear For LifeAlejandro Hernandez100% (4)
- LongevityДокумент12 страницLongevitychai7750% (2)
- WWW Westgard Com Biodatabase1 Htm-2014Документ16 страницWWW Westgard Com Biodatabase1 Htm-2014yousrazeidan1979Оценок пока нет
- Endocrine NotesДокумент157 страницEndocrine NotesLaila Cristine CarvalhoОценок пока нет
- A. Biology of Aging PDFДокумент9 страницA. Biology of Aging PDFlolipopckpОценок пока нет