Вам также может понравиться
- 000504242-Microwave Ephedrine From LpacДокумент4 страницы000504242-Microwave Ephedrine From LpacPaul BrownОценок пока нет
- Interactions Among Soil MicroorganismsДокумент23 страницыInteractions Among Soil MicroorganismsHoD Biotechnology and Microbiology100% (1)
- Alkaloids and OthersДокумент24 страницыAlkaloids and OtherszakybaaОценок пока нет
- Immobilized Enzymes Availability For Glycerol - 1,3 Propanediol BioconversionДокумент6 страницImmobilized Enzymes Availability For Glycerol - 1,3 Propanediol BioconversionAly FirdausОценок пока нет
- The Remarkable Rhodococcus ErythropolisДокумент12 страницThe Remarkable Rhodococcus ErythropolisFrancis Xavier GitauОценок пока нет
- Bio CatalysisДокумент6 страницBio CatalysisNishi SainiОценок пока нет
- Preparatively UsefulДокумент10 страницPreparatively Usefulaniket1209915Оценок пока нет
- The Effect of Lignin Removal by Alkaline Peroxide Pretreatment On The Susceptibility of Corn Stover To Purified Cellulolytic and Xylanolytic EnzymesДокумент11 страницThe Effect of Lignin Removal by Alkaline Peroxide Pretreatment On The Susceptibility of Corn Stover To Purified Cellulolytic and Xylanolytic EnzymesAhmad Rama RamadhanОценок пока нет
- 10 1016@j Ijhydene 2018 09 066Документ8 страниц10 1016@j Ijhydene 2018 09 066Juan Luis Saldaña MéndezОценок пока нет
- Glycosidase Enzymes Stabilization Through Immobilization Onto Nanoparticulated Bimodal OrganosilicasДокумент1 страницаGlycosidase Enzymes Stabilization Through Immobilization Onto Nanoparticulated Bimodal Organosilicasdadang71Оценок пока нет
- Xanthomonas Campestris: Oxygen Supply Without Gas-Liquid Film Resistance To CultivationДокумент10 страницXanthomonas Campestris: Oxygen Supply Without Gas-Liquid Film Resistance To CultivationVipin KumarОценок пока нет
- Effect of Hydraulic Retention Time On Biohydrogen Production and Anaerobic Microbial CommunityДокумент6 страницEffect of Hydraulic Retention Time On Biohydrogen Production and Anaerobic Microbial CommunityAlejandro Duvan Lopez RojasОценок пока нет
- High Cell Density Cultivation and High Recombinant Protein Production of Escherichia Coli Strain Expressing Uricase PDFДокумент6 страницHigh Cell Density Cultivation and High Recombinant Protein Production of Escherichia Coli Strain Expressing Uricase PDFdivya_jspandianОценок пока нет
- Engineering The Third Wave of BiocatalysisДокумент10 страницEngineering The Third Wave of BiocatalysiszottisadОценок пока нет
- 13 67 Lipase DOtensionДокумент7 страниц13 67 Lipase DOtensionNena Och Exha Part IIОценок пока нет
- P.L. Rogers, H.S. Shin and B. Wang - Biotransformation For L-Ephedrine ProductionДокумент27 страницP.L. Rogers, H.S. Shin and B. Wang - Biotransformation For L-Ephedrine ProductionRoundSTICОценок пока нет
- Sustainable Fermentative Hydrogen Production: Challenges For Process OptimisationДокумент9 страницSustainable Fermentative Hydrogen Production: Challenges For Process Optimisationdiky2601Оценок пока нет
- AlgaeДокумент13 страницAlgaearinabrebesОценок пока нет
- Hydrogen Production by The Hyperthermophilic Eubacterium, Using Cellulose Pretreated by Ionic LiquidДокумент15 страницHydrogen Production by The Hyperthermophilic Eubacterium, Using Cellulose Pretreated by Ionic LiquidkimlinhОценок пока нет
- 1 s2.0 S0920586120304600 MainДокумент8 страниц1 s2.0 S0920586120304600 Mainbruno barrosОценок пока нет
- Hydrogen Production From Food Waste in Anaerobic Mesophilic and Thermophilic AcidogenesisДокумент9 страницHydrogen Production From Food Waste in Anaerobic Mesophilic and Thermophilic AcidogenesisĐêmTrắngОценок пока нет
- Yi-Huang Chang, Ku-Shang Chang, Cheng-Wei Huang, Chuan-Liang Hsu, Hung-Der JangДокумент8 страницYi-Huang Chang, Ku-Shang Chang, Cheng-Wei Huang, Chuan-Liang Hsu, Hung-Der JangNoor SalamОценок пока нет
- Kumar 2015Документ8 страницKumar 2015sanjiv KumarОценок пока нет
- Biological Hydrogen Production of The GenusДокумент8 страницBiological Hydrogen Production of The GenusProfessor Douglas TorresОценок пока нет
- Value-Added Utilization of Crude Glycerol From BiodieselДокумент16 страницValue-Added Utilization of Crude Glycerol From BiodieselKaweepoj SiwanartnusornОценок пока нет
- Desain Reaktor IsomerisasiДокумент7 страницDesain Reaktor IsomerisasiIka Y. RachmawatiОценок пока нет
- Catalytic Thermochemical Conversion of Glycerol To Simple and Polyhydric Alcohols Using Raney Nickel CatalystДокумент6 страницCatalytic Thermochemical Conversion of Glycerol To Simple and Polyhydric Alcohols Using Raney Nickel CatalystanitacahyaОценок пока нет
- Paper 1Документ6 страницPaper 1bagherilОценок пока нет
- Production of Clean Fuel From Waste Biomass Using Combined Dark and PhotofermentationДокумент9 страницProduction of Clean Fuel From Waste Biomass Using Combined Dark and PhotofermentationInternational Organization of Scientific Research (IOSR)Оценок пока нет
- Tocatj 4 83Документ5 страницTocatj 4 83IsraelPala-RosasОценок пока нет
- Ethanol ProductionДокумент6 страницEthanol ProductionFAIZ AHMADОценок пока нет
- KrahmerДокумент11 страницKrahmermndrawОценок пока нет
- Art 7Документ6 страницArt 7Rodny NavarroОценок пока нет
- Fed-Batch Fermentation For Production of Nitrile Hydratase by Rhodococcus Rhodochrous M33Документ7 страницFed-Batch Fermentation For Production of Nitrile Hydratase by Rhodococcus Rhodochrous M33Nòng Nọc Cô ĐơnОценок пока нет
- Microbial Cell FactoriesДокумент36 страницMicrobial Cell FactoriesAjay Kumar AОценок пока нет
- Surucu1975 PDFДокумент24 страницыSurucu1975 PDFMohammad Rifan TaufiqurrahmanОценок пока нет
- Phellinus Linteus: Structural and Biological Study of Carboxymethylated PolysaccharidesДокумент5 страницPhellinus Linteus: Structural and Biological Study of Carboxymethylated PolysaccharidesMinh NguyễnОценок пока нет
- Biohydrogen Production by Vermihumus-Associated Microorganisms Using Agro Industrial Wastes As SubstrateДокумент10 страницBiohydrogen Production by Vermihumus-Associated Microorganisms Using Agro Industrial Wastes As SubstrateIvan Dario Alfonso BetancourtОценок пока нет
- THE STRUCTURE OF METOPON - J. Am. Chem. Soc., 1953, 75 (17), PP 4373-4374Документ2 страницыTHE STRUCTURE OF METOPON - J. Am. Chem. Soc., 1953, 75 (17), PP 4373-4374muopioidreceptorОценок пока нет
- Tran2009 Microalga PDFДокумент6 страницTran2009 Microalga PDFAdriana PerezОценок пока нет
- Enhanced Penicillin Production by Oligosaccharides From Batch Cultures of Penicillium Chrysogenum in Stirred-Tank ReactorsДокумент6 страницEnhanced Penicillin Production by Oligosaccharides From Batch Cultures of Penicillium Chrysogenum in Stirred-Tank Reactorsazzahra ghina fadillahОценок пока нет
- Production of Liquid Biofuels From Microalgae Chlorella Sp. Via Catalytic SlowДокумент10 страницProduction of Liquid Biofuels From Microalgae Chlorella Sp. Via Catalytic Slowbambang_teknikkimiaОценок пока нет
- Fermentation of Sun Ower Seed Hull Hydrolysate To Ethanol by Pichia StipitisДокумент8 страницFermentation of Sun Ower Seed Hull Hydrolysate To Ethanol by Pichia StipitisJulio MoralesОценок пока нет
- Manaenkov 2018Документ31 страницаManaenkov 2018Антонина СтепачеваОценок пока нет
- Hyperthermostable Cellulolytic and Hemicellulolytic Enzymes and Their Biotechnological ApplicationsДокумент11 страницHyperthermostable Cellulolytic and Hemicellulolytic Enzymes and Their Biotechnological ApplicationsIngeniero QuimicoОценок пока нет
- A Novel Electroporation Procedure For Highly Efficient Transformation of Lipomyces StarkeyiДокумент7 страницA Novel Electroporation Procedure For Highly Efficient Transformation of Lipomyces Starkeyifilemon jaluОценок пока нет
- Hydrogen Production Characteristics of The Organic Fraction of Municipal Solid Wastes by Anaerobic Mixed Culture FermentationДокумент9 страницHydrogen Production Characteristics of The Organic Fraction of Municipal Solid Wastes by Anaerobic Mixed Culture FermentationAsh MaedaevaОценок пока нет
- Metabolic Engineering of Cyanobacteria For Ethanol ProductionДокумент8 страницMetabolic Engineering of Cyanobacteria For Ethanol ProductionPatrick FuОценок пока нет
- Biodiesel Production From Waste Cooking OilДокумент6 страницBiodiesel Production From Waste Cooking OilAnsam MohОценок пока нет
- PDFДокумент5 страницPDFtary_nuryanaОценок пока нет
- 4.3 BiotransformationДокумент52 страницы4.3 BiotransformationAnika SharmaОценок пока нет
- Process Performance Evaluation of Intermittent-Continuous Stirred Tank Reactor For Anaerobic Hydrogen Fermentation With Kitchen WasteДокумент10 страницProcess Performance Evaluation of Intermittent-Continuous Stirred Tank Reactor For Anaerobic Hydrogen Fermentation With Kitchen WastenabilОценок пока нет
- ScribdДокумент9 страницScribdSrinivas ReddyОценок пока нет
- Bioresources.: Characteristics And Kinetic Analysis Of Β-Glucosidase (Mabgl) From Mucor Ardhlaengiktus Rsc1Документ13 страницBioresources.: Characteristics And Kinetic Analysis Of Β-Glucosidase (Mabgl) From Mucor Ardhlaengiktus Rsc1el batoОценок пока нет
- CatalystsДокумент15 страницCatalystsmarianaortОценок пока нет
- BioRes 05-4-2404 Kurian MBK Bioconver Hemi Hdryolysate Ethanol 770Документ0 страницBioRes 05-4-2404 Kurian MBK Bioconver Hemi Hdryolysate Ethanol 770Ishwar ChandraОценок пока нет
- Cheng2017 e A Via NADHДокумент27 страницCheng2017 e A Via NADHjazorelОценок пока нет
- Radhika 2012Документ10 страницRadhika 2012Pendi Adi MertaОценок пока нет
- Fungi and Lignocellulosic BiomassОт EverandFungi and Lignocellulosic BiomassChristian P KubicekОценок пока нет
- Sustainable synthesis of ciclopentene derivatives through multicomponent reactions in continuous flow regimeОт EverandSustainable synthesis of ciclopentene derivatives through multicomponent reactions in continuous flow regimeОценок пока нет
- Transition Metal Catalyzed Furans Synthesis: Transition Metal Catalyzed Heterocycle Synthesis SeriesОт EverandTransition Metal Catalyzed Furans Synthesis: Transition Metal Catalyzed Heterocycle Synthesis SeriesОценок пока нет
- Heat TransferДокумент16 страницHeat TransferbanerjeetusharОценок пока нет
- Sanskarvidhi Antyeshthi SanskarДокумент9 страницSanskarvidhi Antyeshthi SanskarbanerjeetusharОценок пока нет
- Sanskarvidhi - Vivah SanskarДокумент90 страницSanskarvidhi - Vivah SanskarRakesh TandonОценок пока нет
- XXXIII INCA Full Paper Submission GuidelinesДокумент1 страницаXXXIII INCA Full Paper Submission GuidelinesbanerjeetusharОценок пока нет
- Enzymes Involved in Steroid Nucleas ModificationДокумент7 страницEnzymes Involved in Steroid Nucleas ModificationbanerjeetusharОценок пока нет
- What Skills Does A GIS Analyst RequireДокумент6 страницWhat Skills Does A GIS Analyst RequirebanerjeetusharОценок пока нет
- Radionic Amplifier Energy CirclesДокумент49 страницRadionic Amplifier Energy CirclesTito ZambranoОценок пока нет
- What Are The Causes of Food SpoilageДокумент3 страницыWhat Are The Causes of Food SpoilageMuhammad Arfan JadoonОценок пока нет
- Advanced Microbiome Science To Shield Against Emf Damage UpdatedДокумент16 страницAdvanced Microbiome Science To Shield Against Emf Damage UpdatedSusanaОценок пока нет
- Republic of The PhilippinesДокумент14 страницRepublic of The PhilippinesTheryze Anne BaringОценок пока нет
- 2018 Cohort - Appendix II Course Descriptions 20181123 PDFДокумент88 страниц2018 Cohort - Appendix II Course Descriptions 20181123 PDF與終人.Оценок пока нет
- Kingdoms of LifeДокумент10 страницKingdoms of LifeGerwin Lazaro-Lapuz CortezОценок пока нет
- Medical Microbiology Anesthesia Lecture 2Документ39 страницMedical Microbiology Anesthesia Lecture 2Ubaid KhanОценок пока нет
- LinktreeДокумент31 страницаLinktreeadeebullah43Оценок пока нет
- Characterization of Pathogens: We Strive For Wisdom Chapter-2Документ29 страницCharacterization of Pathogens: We Strive For Wisdom Chapter-2AZ Amanii BossОценок пока нет
- IIT JAM Biotechnology SyllabusДокумент4 страницыIIT JAM Biotechnology Syllabuskumar HarshОценок пока нет
- Final GNM CurriculumДокумент15 страницFinal GNM CurriculumdonpimprikarОценок пока нет
- Science Guidebook For SSC CGL, CHSL, Railways & PSC Exams - Exampundit - in PDFДокумент55 страницScience Guidebook For SSC CGL, CHSL, Railways & PSC Exams - Exampundit - in PDFvilas bollabathiniОценок пока нет
- Microbial Metabolites As Postbiotics: A ReviewДокумент5 страницMicrobial Metabolites As Postbiotics: A ReviewInternational Journal of Innovative Science and Research TechnologyОценок пока нет
- Microbiology OPДокумент1 309 страницMicrobiology OPThư Phạm100% (2)
- Screening and Optimization of Pectinase Producing Bacterial Isolates From Fruit WastesДокумент8 страницScreening and Optimization of Pectinase Producing Bacterial Isolates From Fruit WastesIJRASETPublicationsОценок пока нет
- Canadian Biosafety Standard-EngДокумент188 страницCanadian Biosafety Standard-EngTeobaldo Mazango100% (1)
- Cosmetics MicrobialExamination Is-14648.2011Документ23 страницыCosmetics MicrobialExamination Is-14648.2011I. Murali KrishnaОценок пока нет
- Name:: Dr. Yogesh Sureshrao. BanginwarДокумент8 страницName:: Dr. Yogesh Sureshrao. BanginwarDR. YOGESHОценок пока нет
- Diversity of Soil Fauna and Ecosystem FunctionДокумент14 страницDiversity of Soil Fauna and Ecosystem Functiontariqulmasud12Оценок пока нет
- WJEC GCSE Biology - Cells & Cell MembranesДокумент4 страницыWJEC GCSE Biology - Cells & Cell MembranesMegan TaylorОценок пока нет
- Biochem IntroДокумент6 страницBiochem IntroAlliah CasingОценок пока нет
- Usp 38-1231Документ27 страницUsp 38-1231spiderman00Оценок пока нет
- Critical Reviews in Plant SciencesДокумент46 страницCritical Reviews in Plant ScienceslacisagОценок пока нет
- Reduction of Organic Load and Biodegradation of PaДокумент19 страницReduction of Organic Load and Biodegradation of PaArie Ikhwan SaputraОценок пока нет
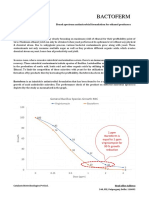
- BactofermДокумент3 страницыBactofermPankaj RajbharОценок пока нет
- Jintexeco Abz-200: Hygiene Protection For TextilesДокумент3 страницыJintexeco Abz-200: Hygiene Protection For Textilesdeepak kumarОценок пока нет
- Introduction To Asian Natural FarmingДокумент47 страницIntroduction To Asian Natural Farminggeen201Оценок пока нет
- Matching Skill Sheets To Topics Book 8Документ6 страницMatching Skill Sheets To Topics Book 8imranОценок пока нет
- A Frightening Interview With CIA Microbiologist Larry Hughes PDFДокумент6 страницA Frightening Interview With CIA Microbiologist Larry Hughes PDFDerrick KimaniОценок пока нет