Вам также может понравиться
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (895)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (344)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (121)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- Pathophysiology of InflammationДокумент50 страницPathophysiology of InflammationElla Jones0% (1)
- Minimally Invasive Cardiac SurgeryДокумент468 страницMinimally Invasive Cardiac Surgeryréda50% (2)
- Guide To Human CD AntigensДокумент22 страницыGuide To Human CD AntigensPablo Rodrigo Acuña MardonesОценок пока нет
- Chapter 2 (Questions 2008-2009 Compiled)Документ89 страницChapter 2 (Questions 2008-2009 Compiled)vetpathforum100% (1)
- Islamic MonthsДокумент2 страницыIslamic MonthssheikTCSОценок пока нет
- Vascularisation of The Lower LimbДокумент8 страницVascularisation of The Lower LimbIdrissou FmsbОценок пока нет
- Muslim, Blessed, Months, Days, Nights, FestivalsДокумент9 страницMuslim, Blessed, Months, Days, Nights, FestivalsabrahammahОценок пока нет
- Citation Guidelines: MLA (Modern Language Association) Citation FormatДокумент7 страницCitation Guidelines: MLA (Modern Language Association) Citation Formatdmh1976Оценок пока нет
- Summary OfLower Limb AnatoДокумент6 страницSummary OfLower Limb AnatoIdrissou FmsbОценок пока нет
- Topographical Anatomy of Lower LimbДокумент15 страницTopographical Anatomy of Lower LimbIdrissou Fmsb100% (1)
- Plantain PileДокумент1 страницаPlantain PileIdrissou FmsbОценок пока нет
- Folong PistДокумент1 страницаFolong PistIdrissou FmsbОценок пока нет
- Carbapenems and Monobactams - 090ct2012Документ6 страницCarbapenems and Monobactams - 090ct2012Idrissou FmsbОценок пока нет
- Alggeo 20130213eДокумент3 страницыAlggeo 20130213eIdrissou FmsbОценок пока нет
- Alggeo 20130802eДокумент3 страницыAlggeo 20130802eIdrissou FmsbОценок пока нет
- Renovascular Hypertension (M2) : G. Ashuntantang-SoméДокумент23 страницыRenovascular Hypertension (M2) : G. Ashuntantang-SoméIdrissou FmsbОценок пока нет
- 16 Anti Protozoal Drugs Lecture9 (0ct10)Документ18 страниц16 Anti Protozoal Drugs Lecture9 (0ct10)Idrissou FmsbОценок пока нет
- PK-PD of Antimicrobial Therapy-Lecture12 Oct11Документ37 страницPK-PD of Antimicrobial Therapy-Lecture12 Oct11Idrissou FmsbОценок пока нет
- 3 Penicillines Oct2012Документ19 страниц3 Penicillines Oct2012Idrissou FmsbОценок пока нет
- 1-Introduction To Pharmacology of Cardiovascular Drugs 0ct10Документ12 страниц1-Introduction To Pharmacology of Cardiovascular Drugs 0ct10Idrissou FmsbОценок пока нет
- 5-Carbapenems and Monobactams - 090ct2012Документ5 страниц5-Carbapenems and Monobactams - 090ct2012Idrissou FmsbОценок пока нет
- 11-Protein Synthesis Inhibitors - Lincomycin & OthersДокумент21 страница11-Protein Synthesis Inhibitors - Lincomycin & OthersIdrissou FmsbОценок пока нет
- 10 Antibiotics Protein Synthesis Inhibitors PhenicolsДокумент11 страниц10 Antibiotics Protein Synthesis Inhibitors PhenicolsIdrissou FmsbОценок пока нет
- 6-LACTAMASE Inhibitors - Oct2013Документ4 страницы6-LACTAMASE Inhibitors - Oct2013Idrissou FmsbОценок пока нет
- Opioid AnalgesicsДокумент13 страницOpioid AnalgesicsIdrissou FmsbОценок пока нет
- 5 Anti Angina Agents Nov2010Документ12 страниц5 Anti Angina Agents Nov2010Idrissou FmsbОценок пока нет
- 4.7-Vasodilators Nov10Документ14 страниц4.7-Vasodilators Nov10Idrissou FmsbОценок пока нет
- Antacids and Anti-Ulcer Agents: Gastrointestinal PharmacologyДокумент56 страницAntacids and Anti-Ulcer Agents: Gastrointestinal PharmacologyIdrissou FmsbОценок пока нет
- Franco-Cameroon Congress 3Документ9 страницFranco-Cameroon Congress 3Idrissou FmsbОценок пока нет
- Local Anesthetic May2010Документ26 страницLocal Anesthetic May2010Idrissou FmsbОценок пока нет
- Evaluation (Hemopoietic Pharm) Jan 2010lДокумент2 страницыEvaluation (Hemopoietic Pharm) Jan 2010lIdrissou FmsbОценок пока нет
- Opioid AnalgesicsДокумент13 страницOpioid AnalgesicsIdrissou FmsbОценок пока нет
- Franco-Cameroon Congress 3Документ9 страницFranco-Cameroon Congress 3Idrissou FmsbОценок пока нет
- Drug List For Health CampaignДокумент1 страницаDrug List For Health CampaignIdrissou FmsbОценок пока нет
- Fixed Drug EruptionsДокумент5 страницFixed Drug EruptionsShalu RjОценок пока нет
- Stafilokokus Aures SeminarskiДокумент81 страницаStafilokokus Aures SeminarskiMersed HuremovicОценок пока нет
- Malaria 2018Документ11 страницMalaria 2018gerges8723Оценок пока нет
- Skin Wounds Vitality Markers in Forensic Pathology - An Updated ReviewДокумент10 страницSkin Wounds Vitality Markers in Forensic Pathology - An Updated Reviewrodrigo100% (1)
- cc11046 PDFДокумент189 страницcc11046 PDFNurfadhilah YusufОценок пока нет
- Art:10.1186/1939 4551 1 S3 A1 PDFДокумент335 страницArt:10.1186/1939 4551 1 S3 A1 PDFYunus wanendaОценок пока нет
- The Role of Endothelial Glycocalyx in Sepsis (DR Budi)Документ6 страницThe Role of Endothelial Glycocalyx in Sepsis (DR Budi)Ovie HoshieОценок пока нет
- Junctional Epithelium: Dr. Ishita Singhal Mds First YearДокумент54 страницыJunctional Epithelium: Dr. Ishita Singhal Mds First YearDR. ISHITA SINGHALОценок пока нет
- ATEROESCLEROSISДокумент228 страницATEROESCLEROSISTenazОценок пока нет
- Medicinal Chemistry & Drug Discovery: Dr. Peter Wipf Department of Chemistry University of PittsburghДокумент69 страницMedicinal Chemistry & Drug Discovery: Dr. Peter Wipf Department of Chemistry University of PittsburghKhushi KhanОценок пока нет
- Diapedesis LeucocitariaДокумент3 страницыDiapedesis LeucocitariaJudhers IsarioОценок пока нет
- Acute and Chronic InflammationДокумент52 страницыAcute and Chronic Inflammationjames20123100% (1)
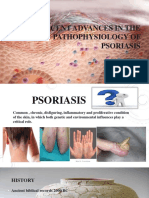
- Recent Advances in The Pathophysiology of Psoriasis: Dr. Mikhin George ThomasДокумент72 страницыRecent Advances in The Pathophysiology of Psoriasis: Dr. Mikhin George ThomasYudi FadilahОценок пока нет
- Etiology and Pathogenesis of Preeclampsia: Current Concepts: Ajog ReviewsДокумент17 страницEtiology and Pathogenesis of Preeclampsia: Current Concepts: Ajog ReviewstapayanaОценок пока нет
- Markers of Inflammation: Dori R. Germolec, Kelly A. Shipkowski, Rachel P. Frawley, and Ellen EvansДокумент23 страницыMarkers of Inflammation: Dori R. Germolec, Kelly A. Shipkowski, Rachel P. Frawley, and Ellen EvansTiến Đạt NguyễnОценок пока нет
- Pathology Book - Chapter 2 - Inflammation - HKДокумент12 страницPathology Book - Chapter 2 - Inflammation - HKagar agarОценок пока нет
- 2006 FOCAL POINTS Dry - Eye PDFДокумент15 страниц2006 FOCAL POINTS Dry - Eye PDFKyros1972Оценок пока нет
- Damage Control OrthopedicsДокумент17 страницDamage Control OrthopedicsterencedszaОценок пока нет
- Intercellular Adhesion Molecule 1 Functions As An Efferocytosis Receptor in in Ammatory MacrophagesДокумент21 страницаIntercellular Adhesion Molecule 1 Functions As An Efferocytosis Receptor in in Ammatory MacrophagesOlfisusan TumbolОценок пока нет
- JCI RG 1 - Ms - 248748Документ56 страницJCI RG 1 - Ms - 248748Alex GoodingОценок пока нет
- Progress in The Mechanism and Targeted Drug Therapy For CopdДокумент20 страницProgress in The Mechanism and Targeted Drug Therapy For CopdliadiafaridaОценок пока нет
- Damage Control Orthopaedics, EvolvingДокумент17 страницDamage Control Orthopaedics, EvolvingAzmi Farhadi100% (1)
- Mechanism of AntipyreticsДокумент12 страницMechanism of AntipyreticsAnamaria Blaga PetreanОценок пока нет
- Reviews: Polymeric Particle-Based Therapies For Acute Inflammatory DiseasesДокумент18 страницReviews: Polymeric Particle-Based Therapies For Acute Inflammatory DiseasesJ. Antonio Ram. M.Оценок пока нет
- Erythrodermic Psoriasis Pathophysiology and CurrenДокумент12 страницErythrodermic Psoriasis Pathophysiology and CurrenDiah Rana HafizhahОценок пока нет
- Acute Viral RhinitisДокумент4 страницыAcute Viral Rhinitisdosen paОценок пока нет