Академический Документы
Профессиональный Документы
Культура Документы
B C - 5a - Transporte Atraves de Membranas
Загружено:
Inês LapaАвторское право
Доступные форматы
Поделиться этим документом
Поделиться или встроить документ
Этот документ был вам полезен?
Это неприемлемый материал?
Пожаловаться на этот документАвторское право:
Доступные форматы
B C - 5a - Transporte Atraves de Membranas
Загружено:
Inês LapaАвторское право:
Доступные форматы
BIOLOGIA CELULAR 69 5 TRANSPORTE ATRAVS DE MEMBRANAS
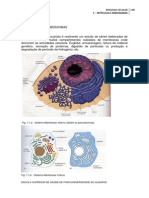
5 TRANSPORTE ATRAVS DE MEMBRANAS 5.1 - MOVIMENTO PARA DENTRO E PARA FORA DAS CLULAS O trfico para dentro e para fora das clulas muito intenso. As clulas trocam gases durante a respirao, absorvem nutrientes e vitaminas, absorvem e libertam gua. As clulas tambm excretam produtos, como por exemplo, certas enzimas e hormonas. As clulas vegetais excretam todas as substncias que vo construir a sua parede celular. Muitas clulas animais recebem impulsos nervosos na forma de substncias qumicas. As clulas animais e vegetais recebem hormonas e substncias de crescimento. A energia entra nas clulas quer na forma qumica quer na forma luminosa. Muita da energia sai da clula sob a forma de calor. Estas afirmaes esto resumidas na figura 5.1.
Fig. 5.1 Movimento da substncias para dentro e para fora das clulas ESCOLA SUPERIOR DE SADE DE FARO/UNIVERSIDADE DO ALGARVE
BIOLOGIA CELULAR 70 5 TRANSPORTE ATRAVS DE MEMBRANAS
5.2 - TRANSPORTE ATRAVS DAS MEMBRANAS CELULARES Do ponto de vista funcional, a discusso do transporte de materiais atravs das membranas celulares pode ser dividida em transporte passivo, em que os materiais se deslocam abaixo do seu gradiente de concentrao, e em transporte activo, que cria ou mantm esses gradientes. 5.2.1 DIFUSO A temperaturas superiores ao 0 absoluto (0K ou -273C) todos os componentes da matria esto em movimento, isto possuem uma certa quantidade de energia cintica. Este movimento ao acaso. As molculas e tomos dirigem-se em todas as direces colidindo umas com as outras. A difuso ocorre ao longo de um gradiente. Quanto maior a variao das concentraes, mais rpido o processo de difuso. Quanto mais concentrado o meio onde ocorre a difuso, mais lentamente se difundem as substncias. A difuso mais rpida em gases do que em lquidos e a temperaturas mais elevadas do que a baixas. A taxa de difuso nos gases inversamente proporcional raiz quadrada da sua densidade. As caractersticas essenciais da difuso so: 1. cada molcula move-se independentemente das outras 2. o movimento ao acaso Como resultado da difuso, a substncia a difundir, fica igualmente distribuda. Difuso simples Alguns materiais podem deslocar-se abaixo do seu gradiente de concentrao por difuso simples atravs da dupla camada lipdica. As pequenas molculas sem carga elctrica, como O2, CO2, NH3, NO, H2O, esterides e agentes lipoflicos, podem entrar ou sair das clulas por difuso simples. O fluxo efectivo desses compostos atravs da membrana proporcional diferena de sua concentrao em ambos os lados, ou, conforme expresso na seguinte equao:
Equao 5.1
Utilizando o sistema de unidades de centmetro-grama-segundo (CGS), J12 o nmero de moles que se move atravs de um centmetro quadrado de membrana do lado 1 para o lado 2 a cada segundo, e C1 e C2 so o nmero de moles do material por centmetro cbico de soluo em ambos os lados. P, a constante de proporcionalidade, denominada permeabilidade da membrana a esse material, em centmetros por segundo. A equao escrita com o sinal menos para lembrar que o fluxo ocorre abaixo do gradiente de concentrao. Essa relao est ilustrada graficamente na Fig. 5.2. A Eq. [5.1] a primeira lei de Fick. Pode ser utilizada para descrever o fluxo de substncias simples sem carga elctrica atravs de qualquer membrana. Por exemplo, mostra-se til
ESCOLA SUPERIOR DE SADE DE FARO/UNIVERSIDADE DO ALGARVE
BIOLOGIA CELULAR 71 5 TRANSPORTE ATRAVS DE MEMBRANAS
para descrever o movimento do oxignio do ar nos alvolos dos pulmes e no sangue, atravs das clulas do epitlio alveolar e endotlio capilar. Se a diferena de potencial atravs da membrana for zero, a lei de Fick tambm pode ser aplicada a substncias com carga elctrica. A permeabilidade descreve a propriedade de determinada membrana em relao a uma substncia especfica. A membrana considerada permevel enquanto as substncias so conhecidas como permeantes ou com capacidade de permear. A permeabilidade proporcional capacidade da substncia de se distribuir pela membrana e de se difundir no seu interior. A permeabilidade inversamente proporcional espessura da membrana. Algumas vezes, conveniente expressar a lei de Fick enunciando que o influxo efetivo de uma substncia igual ao influxo unidireccional (PC0) menos o efluxo unidireccional (PCi). A permeabilidade frequentemente medida com o uso de marcadores radioactivos e estabelecimento da experincia de modo a que a concentrao do marcador num dos lados seja mantida quase nula, medindo-se diretamente o fluxo unidireccional. A permeabilidade a medida da facilidade com que determinado soluto atravessa uma membrana. As duplas camadas lipdicas comuns so relativamente permeveis a pequenas molculas sem carga elctrica; a permeabilidade gua cerca 10-3 cm/s. Por conseguinte, a gua equilibra-se em poucos segundos atravs de uma membrana celular. A uria moderadamente permevel, P=10-6 cm/s e o seu tempo de equilbrio de alguns minutos. As molculas orgnicas pequenas e hidroflicas, como a glicose ou os aminocidos sem carga elctrica, so menos permeveis, com P=10 -7 cm/s e com um tempo de equilbrio de algumas horas; os es so essencialmente impermeveis, com P=10 -12 cm/s e tempos de equilbrio de muitos anos.
Fig. 5.2 A dependncia quanto concentraco, da difuso simples ( esquerda) e da difuso facilitada ( direita).
Exemplo:
Fig. 5.3 - Difuso do oxignio activada pela formao de oxihemoglobina:a combinao do oxignio com a hemoglobina (Hb), nos eritrocitos, para dar a oxihemoglobina (HbO), reduz a concentrao de oxignio no plasma, mantendo sempre um grande gradiente que promove a difuso.
livre
ESCOLA SUPERIOR DE SADE DE FARO/UNIVERSIDADE DO ALGARVE
BIOLOGIA CELULAR 72 5 TRANSPORTE ATRAVS DE MEMBRANAS
5.2.2 DIFUSO FACILITADA Muitas substncias, como a glicose ou a uria, penetram facilmente nas clulas, apesar de terem baixos coeficientes de partio leo-gua; por conseguinte, a dupla camada lipdica relativamente impermevel a essas substncias, O fluxo desses materiais descrito pela lei de Fick apenas para baixas concentraes. Na presena de concentraes mais altas, o fluxo torna-se saturado quando se atinge um valor mximo (ver Fig. 5.2). Esse comportamento pode ser descrito pela equao de Michaelis-Menten, que tambm utilizada para descrever a cintica das enzimas. O fluxo unidireccional fornecido pela seguinte equao:
Equao 5.2
Jmx. o fluxo mximo, e Km a afinidade ou concentrao em que o fluxo metade do seu valor mximo. Essa propriedade saturvel do fluxo sugere que existe um nmero fixo de locais onde ele pode ocorrer. Alm disso, como no caso das enzimas, possvel demonstrar a competio de diferentes substncias pelo mesmo local ou a inibio nocompetitiva dos locais de transporte. Os locais so selectivos para determinada substncia ou grupo de substncias que iro transportar ou que permitem uma competio para o transporte. A selectividade, a afinidade e o Jmx. so trs qualidades independentes dos locais, sendo encontradas com diferentes valores em sistemas distintos. Na actualidade, a difuso facilitada foi elucidada em termos de canais ou transportadores. Os canais exibem, na sua maioria, baixa afinidade ou altos valores de Km; no so saturados em condies fisiolgicas normais. So encontrados trs uniportadores (unitransportadores) de glicose, GLUT1, GLUT3 e GLUT4, em quase todos os tecidos; esses unitransportadores exibem alta afinidade para a glicose e esto saturados em todas as concentraes fisiolgicas. O GLUT2, que encontrado em tecidos caracterizados por grandes fluxos de glicose (como o intestino, o rim e o fgado), possui baixa afinidade para a glicose, e o influxo atravs de transportadores GLUT2 aumenta quando o nvel de glicemia aumenta. Transporte da gua A vida est intimamente associada ao movimento de gua. O corpo constitudo, em sua maior parte, de gua e depende de seu fornecimeneto contnuo para se manter vivo. A gua uma molcula pequena, porm abundante. No muito maior do que um tomo de oxignio, com cerca de 0,2 nm - pequena o suficiente para se intercalar entre outras molculas. Um mol de gua tem 18 ml; por conseguinte, a gua pura tem 55 moI/l. Como as molculas so pequenas, elas movimentam-se com facilidade. Em virtude de sua abundncia, esses movimentos so importantes para o bem-estar do organismo. A morte de lactentes por desidratao em consequncia de diarreia constitui um problema de sade em muitas partes do mundo. Pode ser controlada com gua limpa e uma pequena quantidade de acar e de sal. O edema, frequentemente observado em leses causadas por desportos, mas que tambm ocorre em infeces mais graves, devido ao movimento da gua.
ESCOLA SUPERIOR DE SADE DE FARO/UNIVERSIDADE DO ALGARVE
BIOLOGIA CELULAR 73 5 TRANSPORTE ATRAVS DE MEMBRANAS
Nmero de Avogadro (em homenagem a Amedeo Avogadro) uma constante numrica empregada em Fsica e Qumica. Pode ser definido como o nmero de tomos existentes quando a massa atmica de um elemento expressa em gramas e igual ao peso molecular ( mol). Este nmero aproximadamente igual a 6,02 10. O nmero de Avogadro formalmente definido como o nmero de tomos de carbono-12 em 12 gramas (0,012 kg) de carbono-12, o que aproximadamente igual a 6,02 10. Historicamente, o carbono-12 foi escolhido como substncia de referncia porque sua massa atmica podia ser medida de maneira bastante precisa. O nmero de Avogadro ainda pode ser definido como o nmero de elementos em um mol. Na ausncia de outros factores que influenciem o potencial hdrico ( como por exemplo, a presso), o movimento da gua na osmose ser de uma regio de menor concentrao de soluto (e consequentemente de grande concentrao de gua) para uma regio de maior concentrao de soluto (baixa concentrao de gua). A presena do soluto diminui o potencial hdrico e por isso faz com que a gua v de uma regio de maior para uma de menor potencial hdrico. A presso osmtica pode ser medida por um osmmetro. O principio est esquematizado na figura 5.3. O recipiente contem gua e dentro do tubo est gua com um soluto (sal ou acar). Na entrada do tubo colocada uma membrana semipermevel (permevel gua, mas no ao soluto). A gua que se desloca do recipiente para o tubo, eleva o nvel da soluo dentro do tubo at um equilbrio, ou seja at que o potencial hdrico de ambos os lados da membrana seja igual. A quantidade de presso hidrosttica - neste caso, a fora da gravidade - exercida na coluna de lquido do tubo, iguala a presso osmtica exercida pelo decrscimo de concentrao da soluo no tubo. O movimento da gua no afectado pelo tipo de substncia que est dissolvida na gua, mas pela quantidade de soluto (quantas molculas ou ies).
Fig. 5.4 - a) O tubo contm uma soluo e o recipiente gua destilada
Fig. 5.4 - b) A ua atravessa a mambrana para igualar a concentrao de solutos de ambos os lados
Fig. 5.4 - c) A fora que deve ser aplicada ao pisto para se opor subida no tubo da soluo, a medida da presso osmtica
ISOTNICO- igual concentrao de partculas de ambos os lados da membrana HIPOTNICO- soluo com menos soluto, menor presso osmtica HIPERTNICO- soluo com mais soluto, maior presso osmtica Uma vez que os solutos diminuem o potencial hdrico, uma soluo hipotnica tem um potencial hdrico maior do que uma soluo hipertnica.
ESCOLA SUPERIOR DE SADE DE FARO/UNIVERSIDADE DO ALGARVE
BIOLOGIA CELULAR 74 5 TRANSPORTE ATRAVS DE MEMBRANAS
A clula funciona como um osmmetro. Na presena de um soluto no penetrante como a sacarose, a gua difunde-se de uma regio de maior para uma de menor potencial hdrico. Existem trs mecanismos distintos para o movimento da gua: fluxo de massa, difuso molecular e bombeamento molecular. Quando retiramos a tampa de uma banheira, ou quando seu corao bate, ocorre um fluxo de massa de gua em resposta a uma fora mecnica externa - uma compresso ou uma traco capaz de distender uma mola. A fora propulsora para o fluxo de massa a presso mecnica comumente produzida pela compresso ou pela gravidade. A difuso molecular ou osmose um processo passivo pelo qual a gua se difunde de reas de alta concentrao para reas de baixa concentrao de gua. Existe uma alta concentrao de gua nos locais onde h uma baixa concentrao de solutos, e vice-versa. A gua pode sofrer difuso atravs da maioria das membranas celulares directamente pela dupla camada lipdica ou atravessando canais especficos de gua ou aquaporinas. Muitas clulas produzem aquaporinas, visto que a difuso simples no permite um fluxo adequado de gua. Algumas clulas renais produzem aquaporinas em resposta hormona antidiurtica (ADH), de modo a aumentar o fluxo de gua da urina em formao de volta para o sangue, com consequente conservao da gua. Esse tipo passivo de movimento de gua denominado fluxo osmtico, e a fora propulsora associada o gradiente de concentrao da gua. A gua tambm pode ser transportada atravs das membranas custa de energia pelo cotransportador de Na-glicose (SGLT1). Ao transporte transmembranar de dois ies de Na e uma molcula de acar est associado, dentro da prpria protena, com o influxo de 210 molculas de gua, independentemente do gradiente osmtico. A energia provm do movimento de Na abaixo do seu gradiente de concentrao. Essa bomba molecular proporciona um mecanismo de transporte activo secundrio, que pode responder por quase metade da captao diria de gua do intestino delgado. A presso osmtica refere-se presso mecnica que produz um fluxo de gua igual e oposto ao fluxo osmtico produzido por um determinado gradiente de concentrao. A concentrao refere-se ao somatrio de concentrao molar de todas as partculas, criada quando o soluto dissolvido em gua. medida como osmolaridade, isto , a soma dos moles de cada componente da soluo. Uma soluo 2 mM de MgCl2 contm 6 miliosmoles (mosm) por litro, 2 para o Mg2+ e dois para cada Cl-. A osmolaridade dessa soluo de 6 miliosmolares. Uma soluo 3 mM de NaCI e uma soluo 6 mM de uria possuem a mesma osmolaridade e so designadas como isosmticas. A osmolalidade de uma soluo pode ser medida pela mudana que produz no ponto de congelamento ou presso de vapor. A osmolalidade refere-se aos moles de soluto por quilograma de solvente, enquanto a osmolaridade refere-se aos moles de soluto por litro da soluo. Tonicidade um conceito relacionado com a osmolalidade; entretanto, constitui um caso especial para as clulas. Uma soluo considerada isotnica quando no produz contraco nem dilatao das clulas. Uma soluo 150 mM de NaCI (9 g/l ou 0,9%) isotnica para as clulas de mamferos e tambm isosmtica para o contedo celular. Uma soluo 300 mM de uria tambm isosmtica para o contedo celular; entretanto, uma clula colocada nessa soluo ir aumentar de volume, e, por fim, ir sofrer lise ou ruptura (Fig. 5.4). A soluo de uria hipotnica; possui tonicidade insuficiente para impedir o aumento de volume da clula. Difere da soluo de NaCl, porque a uria pode atravessar a
ESCOLA SUPERIOR DE SADE DE FARO/UNIVERSIDADE DO ALGARVE
BIOLOGIA CELULAR 75 5 TRANSPORTE ATRAVS DE MEMBRANAS
membrana celular. A adio de uma substncia permevel a uma soluo aumenta sua osmolaridade, mas no a sua tonicidade. A adio de solutos mais impermeveis torna uma soluo hipertnica; uma soluo de NaCI de 300 mM hipertnica, causando contrao das clulas. Se for adicionado um soluto moderadamente permevel a uma soluo isotnica (p. ex., 300 mM de uria + 150 mM de NaCl), as clulas sofrem contraco transitria e, a seguir, readquirem o seu volume original (Fig. 5.5). A velocidade com que as clulas se contraem proporcional permeabilidade da membrana gua; a velocidade de recuperao do volume proporcional permeabilidade uria.
Fig.5.5 contrao em solues hipertnicas e aumentam de volume em solues hipotnicas.
As
clulas
sofrem
Fig. 5.6 - A adio de uria provoca contrao transitria, porm no modifica a tonicidade no estado de equilbrio dinmico.
Em alguns casos, conveniente considerar um coeficiente de reflexo como descrio da permeabilidade de solutos. O movimento de gua atravs das paredes capilares depende da diferena de presso mecnica ou hidrosttica e da diferena de presso coloidosmtica, devido a diferenas na concentrao de protenas no plasma e no lquido intersticial. Se a parede capilar for totalmente impermevel s protenas, o seu coeficiente de reflexo de 1,0. Se a parede se tornar permevel, o coeficiente de reflexo ir diminuir, com consequente entrada de protenas no espao intersticial, acompanhadas de gua. O movimento de gua no organismo como um todo est relacionado com dois compartimentos: intracelular e extracelular. O compartimento extracelular constitudo de dois subcompartimentos: o lquido plasmrico nos vasos sanguneos e o lquido intersticial, que banha o restante das clulas. O plasma e o lquido intersticial so separados pelas paredes capilares, que so livremente permeveis a todas as pequenas molculas e ies,
ESCOLA SUPERIOR DE SADE DE FARO/UNIVERSIDADE DO ALGARVE
BIOLOGIA CELULAR 76 5 TRANSPORTE ATRAVS DE MEMBRANAS
mas que normalmente impedem a passagem das prorenas plasmticas para o lquido intersticial. As protenas possuem uma carga negativa efectiva global no pH sanguneo. O equilbrio que surge com uma protena impermevel e ies livremente permeveis denominado equilbrio de Gibbs-Donnan. Esse efeito produz pequenos gradientes de concentrao (<3%) e um pequeno potencial (de alguns mV, negativo no lmen) atravs das paredes dos capilares. Para a maioria dos propsitos clnicos, isso pode ser ignorado e as concentraes inicas no plasma, que so facilmente determinadas, podem ser consideradas como representativas do lquido extracelular em geral. As protenas plasmticas so osmoticamente importantes; tendem a manter a gua dentro dos vasos sanguneos. O equilbrio entre a presso hidrosttica e essa presso osmtica colide, denominado efeito de Starling, regula o fluxo de gua atravs dos endotlios capilares. A perda desse equilbrio resulta na formao de edema. Os volumes e a osmolaridade dos compartimentos corporais podem ser calculados com base nos quatro princpios seguintes: 1. Em todo compartimento, o volume multiplicado pela osmolaridade fornece o nmero de osmoles. 2. A gua movimenta-se entre os compartimentos de modo a tornar a osmolaridade igual em todos eles. 3. A quantidade total de gua e o nmero total de osmoles so obtidos pela soma das quantidades existentes nos compartimentos. 4. Qualquer osmolito adicionado ir permanecer no compartimento extracelular. A adio de gua distribui-se entre os compartimentos, de acordo com os primeiros trs princpios. Por exemplo, considere um estudante de medicina de 70 kg com 16 l de lquido extracelular e 24 l de lquido intracelular, para um total de 40 l. Se a osmolaridade desses compartimentos for de 300 mosM, o compartimento extracelular deve conter 16 x 0,3 = 4,8 osm, e o compartimento intracelular, 7,2 osm, para um total de 12 osm. Caso esse estudante venha a beber 1 l de gua do mar contendo 1 osm de sais, na sua maior parte NaCI, a gua total ir aumentar para 41 l, e o nmero total de osmoles, para 13, de modo que a nova osmolaridade ser de 13/41 = 317 mosM. O compartimento extracelular ter 5,8 osm, de modo que o seu novo volume ser de 5,8/0,317 = 18,3 l. O novo volume intracelular ir atingir 7,2/0,317 = 22,7 l. Observe que a gua saiu das clulas para diluir a gua do mar. O corpo sadio possui mecanismos osmticos destinados a restabelecer o equilbrio original. O rim ir excretar uma urina mais concentrada, e o estudante sentir sede e, provavelmente, ir ingerir gua sem osmolitos. A hiponatremia, ou a presena de baixo nvel sanguneo de sdio, pode levar a srios problemas nos corredores de maratona, que perdem gua e sal em consequncia da transpirao, mas que tendem a repor apenas a gua. Essa situao resulta em sangue hipotnico, de modo que a gua deixa os vasos sanguneos e penetra nas clulas, aumentando o seu volume. Se essa situao ocorrer no crebro, que uma cavidade fechada, essa condio pode levar perda da conscincia e morte.
ESCOLA SUPERIOR DE SADE DE FARO/UNIVERSIDADE DO ALGARVE
BIOLOGIA CELULAR 77 5 TRANSPORTE ATRAVS DE MEMBRANAS
Canais de gua Algumas clulas exigem uma maior permeabilidade a gua do que a proporcionada pela dupla camada lipdica. Os eritrcitos, que precisam modificar rapidamente a sua forma para atravessar os capilares estreitos, e algumas clulas epiteliais, principalmente no rim, possuem canais de gua especializados ou aquaporinas (AQP), que permitem a passagem de gua, porm excluem a de ies. As AQP so encontradas como tetrameros com quatro poros funcionais, um em cada subunidade. As subunidades apresentam seis segmentos transmembranares e duas regies semelhantes ala P dos canais K. Uma das alas est voltada para a superfcie extracelular, enquanto a outra est voltada para o citoplasma, encontrando-se no meio da membrana.
Fig. 5.7 a) Comportamento de glbulos vermelhos em solues hipertnicas Fig. 5.7 b) Comportamento de glbulos vermelhos em solues hipotnicas
5.2.3 - CANAIS Os canais podem ser diferenciados pelo seu mtodo de abertura em canais mecanossensveis envolvidos em processos sensoriais, canais sensveis voltagem envolvidos na propagao do potencial de aco, e canais quimiossensveis envolvidos na transmisso sinptica. Existem tambm canais que esto habitualmente abertos, como os canais que mantm o potencial em repouso, os canais de gua e os canais intercelulares especializados, que ligam o citoplasma de uma clula com o de outra clula.
ESCOLA SUPERIOR DE SADE DE FARO/UNIVERSIDADE DO ALGARVE
BIOLOGIA CELULAR 78 5 TRANSPORTE ATRAVS DE MEMBRANAS
Canais mecanossensveis (mecanorreceptores) Os canais mecanossensveis (mecanorreceptores) formam uma classe distinta de canais estruturalmente no-relacionados, que desempenham numerosas funes diferentes em diferentes clulas. A mecanossensibilidade (mecanorreceptividade) importante para o tacto e a audio, bem como para a propiocepo, fornecendo informaes sobre a posio, a orientao, a velocidade e a acelerao do corpo e suas partes. Os canais esto associados a molculas e estruturas celulares acessrias que amplificam as suas funes especficas. As clulas no-sensoriais somticas tambm respondem ao stress mecnico sem levar a informao ao sistema nervoso - por exemplo, para compensar o intumescimento osmtico ou para modular a secreo ou a contrao. Os organismos unicelulates que nadam, quando se deparam com uma barreira, podem responder invertendo a sua direco. As bactrias possuem canais mecanossensveis (mecanorreceptores) capazes de responder a mudanas sbitas no ambiente osmtico, actuando como valvula de segurana. Muitos canais mecanossensveis (mecanorreceptores) consistem em canais catianicos relativamente no-seletivos. Alguns desses canais so muito grandes e permitem a passagem de electrlitos e pequenos metaboltos, mas no de protenas, atravs da membrana. Canais sensveis voltagem Os canais de K sensveis voltagem (KV) so responsveis pelo retorno ao estado de repouso, levando ao trmino de um potencial de aco. O KV possui um cerne semelhante ao do Kir (canal de potssio rectificador) e quatro hlices trans-membranares (TM) adicionais em cada subunidade (Fig. 5.8). O quarto segmento TM (S4) destaca-se pela presena de quatro a oito cadeias laterais de carga positiva (Arg ou Lys). O S4 uma caracterstica dos canais sensveis voltagem. Acredita-se que constitui o sensor de voltagem que se desloca em direco superfcie extracelular quando o potencial de membrana se modifica, produzindo as alteraes conformacionais que levam abertura do canal. Existem nove subfamlias de canais K e vrias subfamlias de canais 6-TM, incluindo os canais K activados pelo Ca, os canais activados por hiperpolarizao, que so importantes na actividade do corao, e os canais regulados por nucleotdios cclicos. As ltimas duas famlias consistem em canais catinicos no-selectivos.
Fig. 5.8 - Topologia de um monmero de canais de K dependentes de voltagem (KV)
ESCOLA SUPERIOR DE SADE DE FARO/UNIVERSIDADE DO ALGARVE
BIOLOGIA CELULAR 79 5 TRANSPORTE ATRAVS DE MEMBRANAS
Os canais de Na sensveis voltagem (NaV) so responsveis pela fase ascendente do potencial de aco e sustentam a sua propagao. Os canais de Ca sensveis voltagem (CaV) acumulam as alteraes do potencial de membrana com um aumento na concentrao intracelular de Ca, que actua como segundo mensageiro no controle de uma variedade de processos intracelulares. Os canais Na V e CaV possuem uma estrutura semelhante aos canais KV, excepto que consistem em molculas simples maiores que incorporam quatro domnios, tendo, cada um deles, segmentos 6-TM ligeiramente diferentes (Fig. 5.9). Os filtros de selectividade possuem quatro paredes diferentes. O canal de CaV apresenta quatro glutamatos caractersticos (EEEE) no revestimento do poro, um em cada domnio. O canal de Na V exibe um padro DEKA (ver anexo final) nas quatro paredes de seu poro. Essas cadeias laterais precisam estar expostas ao lmen do poro. A selectividade do canal determinada pelas cargas eltricas expostas ao lmen e pelo tamanho do poro.
Fig. 5.9 Topologia dos canais de Na dependentes de voltagem (NaV)
Canais quimiossensveis Existem muitos canais quimiossensveis ou regulados por ligantes diferentes. Esses canais controlam o fluxo de ies e, portanto, geram sinais elctricos em resposta a substncias qumicas especficas, como a acetilcolina (ACh), o glutamato ou ATP. Podem ser agrupados em trs superfamlias, de acordo com a estequiometria e a topologia de membrana das suas subunidades. Muitos foram inicialmente descobertos em farmacologia quando foi constatado que certos compostos, denominados agonistas, produziam correntes de membrana ou alteravam a actividade elctrica das clulas e de outros compostos, enquanto os antagonistas eram capazes de bloquear esses efeitos. Para algumas correntes induzidas por agonistas, o ligante liga-se mesma molcula que contm o poro. Trata-se dos canais regulados por ligantes, que algumas vezes so denominados receptores de ligantes ionotrpicos para distingui-los dos receptores de ligantes metabotrpicos, em que o ligante liga-se a um receptor acoplado protena G (GPCR) e desencadeia uma cascata bioqumica, que pode incluir a abertura de outros canais, como, por exemplo KACh. Os canais receptores de ACh (AChR) so denominados AChR nicotnicos ou nAChR. O termo nicotnico indica que esses receptores ligam-se nicotina, que tambm determina a abertura dos canais. Os nAChR so encontrados nas membranas ps-sinpticas nas junes neuromusculares esquelticas, bem como nos sistemas nervosos autnomo e central.
ESCOLA SUPERIOR DE SADE DE FARO/UNIVERSIDADE DO ALGARVE
BIOLOGIA CELULAR 80 5 TRANSPORTE ATRAVS DE MEMBRANAS
Os nAChR mais extensamente estudados so pentmeros heteromricos. Os monmeros apresentam, cada um, quatro segmentos TM e uma grande regio N-terminal extracelular. A ligao da ACh induz uma alterao conformacional que abre o poro formado no nvel da membrana lipdica e revestido pelo segundo segmento TM de cada uma das cinco subunidades monomticas. Os canais abertos so altamente permeveis ao Na e ao K, ligeiramente permeveis ao Ca e impermeveis aos anies.
Fig. 5.10 A Topologia de um monmero dos canais receptores nicotnicos da acetilcolina (nAChR), com vista superior mostrando a disposio dos cinco monmeros. . B. Vista lateral do canal.
Os receptores ps-sinpticos no SNC para a glicina (glyR), o cido gamaminobutrico (GABAAR) e a serotonina (5HT3R) exibem uma arquitectura pentamrica, embora alguns sejam homomricos, como uns nAChR. Os glyR e GABAAR so selectivamente permeveis a anies e produzem potenciais ps-sinpticos inibitrios (PPSI). Os 5HT3R so selectivos pata caties, semelhana dos nAChR, e produzem potenciais ps-sinpticos excitatrios (PPSE). Os canais de PPSE mais comuns no SNC so receptotes de glutamato (gluR), que apresentam uma arquitectura (Fig. 5.11) de tetrmeros hetetomricos, com trs TM por subunidade. Possuem uma grande regio extracelular com quatro locais de ligao de glutamato e uma ala P voltada para o citoplasma.
Fig. 5.11 - Topologia de um monmero dos canais do receptor de glutamato (gluR), com vista superior mostrando a disposio dos quatro monmeros.
ESCOLA SUPERIOR DE SADE DE FARO/UNIVERSIDADE DO ALGARVE
BIOLOGIA CELULAR 81 5 TRANSPORTE ATRAVS DE MEMBRANAS
Duas outras famlias de canais quimiossensveis so a famlia do canal de sdio epitelial (ENaC) e a famlia do receptor de IP3 (IP3R). Os ENaC so importantes na reabsoro de sdio da urina, em formao, nos tbulos do nefrnio. Acredita-se que os ENaC sejam tetrmeros heteromricos, apresentando, cada um, dois segmentos TM. Esses canais no so dependentes de voltagem. Sabe-se que so regulados por controle da sua insero e remoo da membrana, e alguns pesquisadores suspeitam da existncia de um ligante desconhecido para esse canal. Existem canais estruturalmente relacionados com invertebrados que possuem ligantes conhecidos. Os IP3R e os receptores de rianodina (RyR) relacionados so encontrados na membrana do retculo endoplasmtico. Quando abertos, permitem a liberao de Ca do retculo endoplasmrico. O trifosfato de inositol (IP3) um segundo mensageiro produzido pela aco da fosfolipase C (PLC) sobre o lpido da membrana, o fosfatidilinositol, que foi previamente fosforilado para constituir o PIP2. Os RyR tambm controlam a liberao de clcio, do retculo sarcoplasmtico do msculo,. A rianodina refere-se a uma toxina que abre parcialmente esses canais. Os RyR so abertos por uma interaco directa com um canal CaV modificado no musculo esqueltico e pelo Ca intracelular no msculo cardaco. Os RyR so homotetrmeros com uma enorme regio N-terminal citoplasmtica de 20 nm de dimetro. O peso molecular total do tetrmero cerca de 10 vezes maior que o dos canais NaV ou KV. Os canais dos IP3R tambm so homotetrmeros, cujo tamanho aproximadamente metade do tamanho dos RyR. Foi deduzido que os IP 3R possuem seis segmentos TM por monmero, enquanto os RyR apresentam quatro a doze.
Transporte facilitado Este processo semelhante difuso passiva uma vez que nenhum do dois necessitam gastar energia e ambos so reversveis, dependendo da concentrao num e noutro lado da membrana. Contudo a difuso facilitada envolve transporte de uma substncia atravs de um transportador especfico da membrana. A dupla camada lipdica da membrana celular forma uma barreira a aucares, amino cidos, nucletidos e muitos outros nutrientes essenciais. Na difuso facilitada, existe uma espcie de sistema de vai e vem no qual, uma molcula proteica que se pode deslocar atravs da membrana, liga-se a uma outra molcula especfica que no se pode deslocar atravs da membrana presumivelmente do mesmo modo que uma enzima se liga ao seu substrato. As molculas combinadas, difundem-se atravs da membrana para o interior da clula onde a molcula "escoltada" libertada. Este transporte faz-se graas a enzimas (permeases). Existe uma complementaridade precisa do transportador (permease) com a substncia transportada cujo nome geral "ligando". O transportador especfico para um determinado substrato. Por exemplo, se uma membrana tiver s transportadores de glucose, outros carbohidratos no entram na clula.
ESCOLA SUPERIOR DE SADE DE FARO/UNIVERSIDADE DO ALGARVE
BIOLOGIA CELULAR 82 5 TRANSPORTE ATRAVS DE MEMBRANAS
Fig. 5.12 - Vrias fases do transporte facilitado. De acordo com este modelo, a difuso facilitada envolve uma protena transmembranar que cria um canal hidroflico para um soluto polar especfico e com capacidade para ter duas conformaes, uma aberta para o exterior e outra para o interior. Este exemplo representa o transporte de glucose atravs da membrana plasmtica do eritrcito, esquematizado em 3 passos.(1) a protena transportadora apresenta o local de ligao aberto para o exterior. A molcula de Dglucose colide e liga-se a esse local.(2) com a glucose ligada, a protena tem de tomar a outra conformao, abrindo-se para o interior. (3) com a libertao da molcula de glucose a protena volta posio inicial, abrindo novamente para o exterior.
BIBLIOGRAFIA Azevedo, C. (1999) . Biologia Celular e Molecular . Lidel, Edies Tcnicas, Lisboa. Becker, W. M., Reece, J. B., Poernie, M. F. (1996) . The World of the Cell. The Benjamin/Cummings Publishing Company, California. Junqueira, L.C., Carneiro, J. (2004) . Histologia Bsica . Ed. Guanabara Koogan S.A., Rio de Janeiro. Kierszenbaum, A.L. (2004) . Histologia e Biologia Celular . Elsevier, Rio de Janeiro. Landowne, D. (2007) . Fisiologia Celular . McGraw-Hill, So Paulo. Lodish, H., et al. (1995) . Molecular Cell Biology . Scientific American Books, New York.
ESCOLA SUPERIOR DE SADE DE FARO/UNIVERSIDADE DO ALGARVE
BIOLOGIA CELULAR 83 5 TRANSPORTE ATRAVS DE MEMBRANAS
ANEXO
Simbologia e Nomenclatura
Na nomenclatura dos aminocidos, a numerao dos carbonos da cadeia principal iniciada a partir do carbono do carboxilo.
Nome Glicina ou Glicocola Alanina Leucina Valina Isoleucina Prolina Fenilalanina Serina Treonina Cisteina Tirosina Asparagina Glutamina Aspartato ou cido asprtico Glutamato ou cido glutmico Arginina Lisina Histidina Triptofano Smbolo Gly, Gli Ala Leu Val Ile Pro Phe ou Fen Ser Thr, The Cys, Cis Tyr, Tir Asn Gln Asp Glu Arg Lys, Lis His Trp, Tri Abreviao Nomenclatura G A L V I P F S T C Y N Q D E R K H W cido 2-aminoactico ou cido 2-amino-etanico cido 2-aminopropinico ou cido 2-amino-propanico cido 2-aminoisocaprico ou cido 2-amino-4-metilpentanico cido 2-aminovalrico ou cido 2-amino-3-metil-butanico cido 2-amino-3-metil-n-valrico ou cido 2-amino-3-metilpentanico cido pirrolidino-2-carboxlco cido 2-amino-3-fenil-propinico ou cido 2-amino-3-fenilpropanico cido 2-amino-3-hidroxi-propinico ou cido 2-amino-3hidroxi-propanico cido 2-amino-3-hidroxi-n-butrico cido 2-bis-(2-amino-propinico)-3-dissulfeto ou cido 3tiol-2-amino-propanico cido 2-amino-3-(p-hidroxifenil)propinico ou paraidroxifenilalanina cido 2-aminossuccionmico cido 2-aminoglutarmico cido 2-aminossuccnico ou cido 2-amino-butanodiico cido 2-aminoglutrico cido 2-amino-4-guanidina-n-valrico cido 2,6-diaminocaprico ou cido 2, 6-diaminoexanico cido 2-amino-3-imidazolpropinico cido 2-amino-3-indolpropinico
ESCOLA SUPERIOR DE SADE DE FARO/UNIVERSIDADE DO ALGARVE
Вам также может понравиться
- Anatomia Do Sistema RenalДокумент33 страницыAnatomia Do Sistema RenalFernando Busanelli100% (1)
- Distúrbios Hidroeletrolíticos - CesmacДокумент59 страницDistúrbios Hidroeletrolíticos - CesmacMichell Barbosa Aquino100% (1)
- ESMO ACF Cancro Colorretal Um Guia para o Doente PDFДокумент53 страницыESMO ACF Cancro Colorretal Um Guia para o Doente PDFInês LapaОценок пока нет
- ExerciciosEstudosAnaliticos PDFДокумент9 страницExerciciosEstudosAnaliticos PDFInês Lapa0% (1)
- Miologia PDFДокумент15 страницMiologia PDFInês LapaОценок пока нет
- Sistema Tegumentar PDFДокумент71 страницаSistema Tegumentar PDFInês LapaОценок пока нет
- Sistema Endócrino PDFДокумент56 страницSistema Endócrino PDFInês Lapa100% (1)
- Sistema Urinário PDFДокумент30 страницSistema Urinário PDFInês Lapa100% (1)
- Sistema Digestivo PDFДокумент74 страницыSistema Digestivo PDFInês LapaОценок пока нет
- Pigmentos+de+Plantas+ +TLCДокумент11 страницPigmentos+de+Plantas+ +TLCInês LapaОценок пока нет
- Resposta InflamatóriaДокумент48 страницResposta InflamatóriaInês Lapa100% (1)
- B C - 7 - Reticulos e RibossomasДокумент18 страницB C - 7 - Reticulos e RibossomasInês LapaОценок пока нет
- Disturbios Hidro-EletrolíticoДокумент14 страницDisturbios Hidro-EletrolíticoJosé Bruno João PomboОценок пока нет
- Revisão EnemДокумент5 страницRevisão EnemAnita Burgan100% (1)
- Capítulo Choque PDFДокумент42 страницыCapítulo Choque PDFlendalindaОценок пока нет
- Aspectos Conceituas e Fisiologia Do EnvelhecimentoДокумент42 страницыAspectos Conceituas e Fisiologia Do EnvelhecimentoTássia CristineОценок пока нет
- Princípios Físicos Da Água 1Документ64 страницыPrincípios Físicos Da Água 1Anderson PauloОценок пока нет
- 1 Terapia Nutricional em Nefropatias e Nutrição Hospitalar - PDF Versão 1Документ28 страниц1 Terapia Nutricional em Nefropatias e Nutrição Hospitalar - PDF Versão 1Marcelle AbreuОценок пока нет
- Ficha Informativa - OsmorregulaçãoДокумент8 страницFicha Informativa - OsmorregulaçãoMaria Clara GregoОценок пока нет
- XXIV III Avaliacao Laboratorial Liquido AsciticoДокумент15 страницXXIV III Avaliacao Laboratorial Liquido AsciticoDaniberg Rimenis100% (1)
- Distúrbios Do SódioДокумент50 страницDistúrbios Do SódioLuiz Henrique AlvesОценок пока нет
- Disturbios HidroeletroliticosДокумент40 страницDisturbios HidroeletroliticosTaely GomesОценок пока нет
- Apostila de HemodiáliseДокумент70 страницApostila de HemodiáliseANGELAОценок пока нет
- Cap. 12 Sistema DigestórioДокумент20 страницCap. 12 Sistema DigestórioanaОценок пока нет
- Resumo Do Cap 28 Do Guyton - Passei DiretoДокумент6 страницResumo Do Cap 28 Do Guyton - Passei DiretoCarlaОценок пока нет
- Questões Balanço Hídrico 2019.03Документ4 страницыQuestões Balanço Hídrico 2019.03Diário de Ana e AnaОценок пока нет
- Biologia Sistema Endocrino ExerciciosДокумент18 страницBiologia Sistema Endocrino ExerciciosPaulCumbassОценок пока нет
- Biologia Exercicios Fisiologia Animal Sistema Endocrino Gabarito ResolucaoДокумент18 страницBiologia Exercicios Fisiologia Animal Sistema Endocrino Gabarito ResolucaoCaioSilvaОценок пока нет
- Fisiologia - Lista de Exercícios 02Документ20 страницFisiologia - Lista de Exercícios 02Anderson B. RollaОценок пока нет
- Regulação e ExcreçãoДокумент13 страницRegulação e ExcreçãoHASFTAGОценок пока нет
- Roteiro RenalДокумент3 страницыRoteiro Renalnaldo111100% (1)
- Patologias Do Sistema EndócrinoДокумент6 страницPatologias Do Sistema EndócrinoMiguel SousaОценок пока нет
- Uel 2012Документ12 страницUel 2012LaraCocheteОценок пока нет
- Ebook Extensivo Medicina - Semana 33Документ270 страницEbook Extensivo Medicina - Semana 33djeifnaОценок пока нет
- 1o Simulado para Fhemig Mentoria FVGДокумент29 страниц1o Simulado para Fhemig Mentoria FVGPedro HenriqueОценок пока нет
- Função Tubular e Controle Hormonal - 2011Документ27 страницFunção Tubular e Controle Hormonal - 2011Gabriela FariaОценок пока нет
- HORMÔNIOS Hipofisários 2 CicloДокумент3 страницыHORMÔNIOS Hipofisários 2 CicloBruna CapattiОценок пока нет
- HomeostaseДокумент9 страницHomeostaseEdgar MarcusОценок пока нет
- Resumo Do Sistema EndócrinoДокумент6 страницResumo Do Sistema EndócrinoAdna LimaОценок пока нет