Вам также может понравиться
- Epitelio GlandularДокумент28 страницEpitelio GlandularBladimir Mora100% (2)
- Estructura Del EspermatozoideДокумент7 страницEstructura Del EspermatozoideedispaОценок пока нет
- Tarea de Tejido ConjuntivoДокумент6 страницTarea de Tejido ConjuntivoLurdesita GcОценок пока нет
- Origen Embriológico de RiñonДокумент1 страницаOrigen Embriológico de RiñonGlenn GarcíaОценок пока нет
- Tejido EpitelialДокумент44 страницыTejido EpitelialHenry Petit RodriguezОценок пока нет
- Guia 05 Tejido Conectivo PDFДокумент8 страницGuia 05 Tejido Conectivo PDFJuan Carlos Robayo100% (4)
- Cuestionario de Tejido ConjuntivoДокумент4 страницыCuestionario de Tejido ConjuntivoAlondraОценок пока нет
- Celula Eucariota y ProcariotaДокумент19 страницCelula Eucariota y ProcariotaChocolatesDoñaMaruОценок пока нет
- Fibras Tejido Conjuntivo. Teórico 5 PDFДокумент14 страницFibras Tejido Conjuntivo. Teórico 5 PDFValentina GuerreroОценок пока нет
- Seminario de Tejido MuscularДокумент19 страницSeminario de Tejido Musculardanly castilloОценок пока нет
- T CartilaginosoДокумент8 страницT CartilaginosoBelén Ramos MoralesОценок пока нет
- EpitelioДокумент13 страницEpitelioLeidyYutteОценок пока нет
- Práctica 3. Tejido Conjuntivo OrdinarioДокумент5 страницPráctica 3. Tejido Conjuntivo OrdinarioElizette AlverОценок пока нет
- Tejido Epitelial y ConectivoДокумент20 страницTejido Epitelial y Conectivoapi-26818885100% (5)
- Fichas Tejido ConectivoДокумент23 страницыFichas Tejido ConectivoMicaela HerrОценок пока нет
- ESPERMATOGÉNESISДокумент26 страницESPERMATOGÉNESISEduardo Ocampo100% (1)
- Citoesqueleto y MovimientoДокумент14 страницCitoesqueleto y Movimientoandreaave100% (2)
- Practica 4. Epitelios de RevestimientoДокумент21 страницаPractica 4. Epitelios de RevestimientoNorma GLОценок пока нет
- Matriz ExtracelularДокумент5 страницMatriz ExtracelularJose Luis Burbano VОценок пока нет
- Sistema CirculatorioДокумент7 страницSistema CirculatorioRoger Orlando LunaОценок пока нет
- Materia Completa HistologíaДокумент134 страницыMateria Completa HistologíaSabri JaramilloОценок пока нет
- Tejido Epitelial Usmp HistologiaДокумент63 страницыTejido Epitelial Usmp HistologiaAdita Barrantes CastilloОценок пока нет
- Tejido EpitelialДокумент29 страницTejido EpitelialDaniela AlarcónОценок пока нет
- División CelularДокумент10 страницDivisión CelularAndrea Fernanda Calderon CuadrosОценок пока нет
- Orgánulos No MembranososДокумент5 страницOrgánulos No Membranososalberto_fernandez_92100% (3)
- Histologia TrabajoДокумент29 страницHistologia TrabajoLinda Stefania Perez Zavalla0% (1)
- Mesodermo y MesenquimaДокумент3 страницыMesodermo y MesenquimaAntoAlvaradoОценок пока нет
- Tejido ConectivoДокумент12 страницTejido ConectivoAnonymous 9enVOQB8fОценок пока нет
- Tejido EpitelialДокумент10 страницTejido EpitelialKeyla Vente VelascoОценок пока нет
- Tejido EpiterialДокумент11 страницTejido EpiterialGaby CordovaОценок пока нет
- Citotecnología CompletoДокумент78 страницCitotecnología CompletoIngrid Parrales100% (1)
- Desarrollo de Las Gonadas Masculinas y FemeninasДокумент3 страницыDesarrollo de Las Gonadas Masculinas y FemeninasAle AntonioОценок пока нет
- Infografía Sobre MeiosisДокумент5 страницInfografía Sobre Meiosismateo velezОценок пока нет
- Tejido Conectivo - HistologiaДокумент22 страницыTejido Conectivo - HistologiaWilson AugustoОценок пока нет
- Desarrollo Del Aparato RespiratorioДокумент2 страницыDesarrollo Del Aparato RespiratorioAngel Stevens Garcia CallaoОценок пока нет
- Resumen Capitulo 3 GuytonДокумент4 страницыResumen Capitulo 3 GuytonconyОценок пока нет
- Histologia Normal de La PlacentaДокумент21 страницаHistologia Normal de La PlacentaVanessa Ricalde Roldán75% (4)
- La Formación de Mórula, Blastocisto y GastrulaciónДокумент21 страницаLa Formación de Mórula, Blastocisto y GastrulaciónMarlene ToledanoОценок пока нет
- Histología - Aparato DigestivoДокумент31 страницаHistología - Aparato Digestivoanthony solorzano100% (2)
- Histología Del OídoДокумент11 страницHistología Del OídoluisОценок пока нет
- EmbriologiaДокумент5 страницEmbriologiaJhey Kenider Benites IzquierdoОценок пока нет
- Colageno y ElastinaДокумент9 страницColageno y ElastinaJosé NaranjoОценок пока нет
- Resumen Histologia 2Документ58 страницResumen Histologia 2Dimi Mattos Dos SantosОценок пока нет
- Uniones CelularesДокумент3 страницыUniones CelularesAndreaОценок пока нет
- Organelos No Membranosos - CLASE 3Документ21 страницаOrganelos No Membranosos - CLASE 3Víctor Manuel MoralesОценок пока нет
- Tejido Conectivo Adiposo-Cartilaginoso-OseoДокумент2 страницыTejido Conectivo Adiposo-Cartilaginoso-OseoAlexandra AramayoОценок пока нет
- CitoesqueletoДокумент50 страницCitoesqueletoBrian Fariña100% (1)
- Tejido NerviosoДокумент21 страницаTejido NerviosoDilan TorresОценок пока нет
- EstomagoДокумент44 страницыEstomagoBeberly Reyes EspinozaОценок пока нет
- Informe 3 Embriología SeminarioДокумент8 страницInforme 3 Embriología SeminarioEmery Sofía C. D. BustamanteОценок пока нет
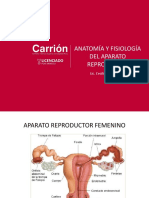
- Anatomia Reproductor Femenino-2021Документ23 страницыAnatomia Reproductor Femenino-2021Oldy YanQui100% (1)
- Uniones IntercelularesДокумент27 страницUniones IntercelularesSusana Salas NuñezОценок пока нет
- CUELLOДокумент48 страницCUELLOAndrés Rengifo OrjuelaОценок пока нет
- Informe Anatomía Humana II Sesión 3Документ7 страницInforme Anatomía Humana II Sesión 3Angie Flores MamaniОценок пока нет
- Tejido LinfoideДокумент4 страницыTejido Linfoided0ugy100% (2)
- Citopatologia. Pap 1.Документ168 страницCitopatologia. Pap 1.Julio Ramiro De La Cruz SernaqueОценок пока нет
- Uniones IntercelularesДокумент6 страницUniones IntercelularesAgustina NuñezОценок пока нет
- Guia Adhesion Celular - 4toELECTIVOДокумент4 страницыGuia Adhesion Celular - 4toELECTIVOMaria Alicia Bravo ArriagadaОценок пока нет
- Complejos de UniónДокумент7 страницComplejos de UniónAnalia Moraes100% (1)
- Adhesión y Unión CelularДокумент5 страницAdhesión y Unión CelularFrancisco AriasОценок пока нет
- Mapa Mental de Efectos de Las DrogasДокумент4 страницыMapa Mental de Efectos de Las DrogasAnonymous yWM3Za88% (17)
- Cuestionario #6. ListoДокумент13 страницCuestionario #6. ListoINDIANA MEJIAОценок пока нет
- Valoracion Geriatria222 PDFДокумент6 страницValoracion Geriatria222 PDFMonse Morales SantaellaОценок пока нет
- Seminario 1 Dra YUSLENNYДокумент15 страницSeminario 1 Dra YUSLENNYYuslenny AponteОценок пока нет
- Biologia 8 Semana 6 3PДокумент5 страницBiologia 8 Semana 6 3Pfelipe hernandezОценок пока нет
- Revista Farmaco Artículo AnticonbulcionantesДокумент6 страницRevista Farmaco Artículo AnticonbulcionantesdanielaОценок пока нет
- 2011 Pro Trauma Perm FrasДокумент0 страниц2011 Pro Trauma Perm FrasSaylin ArgüelloОценок пока нет
- Toxocara CanisДокумент56 страницToxocara CanisStephane LovonОценок пока нет
- Primer Parcial AnatoДокумент3 страницыPrimer Parcial AnatoMontse MartínezОценок пока нет
- Respuestas Subcelulares A La LesiónДокумент2 страницыRespuestas Subcelulares A La LesiónJheyris PintoОценок пока нет
- Que Es El IgssДокумент4 страницыQue Es El IgssRosalva AcОценок пока нет
- Volleyball AsignacionДокумент23 страницыVolleyball AsignacionKammla Miranda100% (3)
- NecropsiaДокумент13 страницNecropsiaSam SaharaОценок пока нет
- Prevención ATEL 3Документ3 страницыPrevención ATEL 3Angelica Mancipe SastoqueОценок пока нет
- Historia de La Medicina en El RenacimientoДокумент55 страницHistoria de La Medicina en El RenacimientoAngèlica QuielОценок пока нет
- AvesДокумент34 страницыAvesBryan CoelloОценок пока нет
- Aparato DigestivoДокумент13 страницAparato DigestivoMontse RodriguezОценок пока нет
- ODONTOLOGIA FORENSE - Cuando Los Dientes HáblanДокумент16 страницODONTOLOGIA FORENSE - Cuando Los Dientes HáblanHeber Especilista LaboralОценок пока нет
- 4 - Alteracionesdel CrecimientoДокумент51 страница4 - Alteracionesdel CrecimientoK - CastroОценок пока нет
- Tecnologia de La Carne 1234756650412903 1Документ24 страницыTecnologia de La Carne 1234756650412903 1humbertoОценок пока нет
- El Calentamiento y Estiramiento FisicoДокумент12 страницEl Calentamiento y Estiramiento FisicoSocrates Huanca PazОценок пока нет
- Informe Biologia CeluassДокумент10 страницInforme Biologia CeluassGabriela Vera MorenoОценок пока нет
- Apuntes de La Maestra Nektli RojasДокумент63 страницыApuntes de La Maestra Nektli RojasDiana Álvarez100% (1)
- Actividad Integrativa Fotosíntesis y Respiración CelularДокумент4 страницыActividad Integrativa Fotosíntesis y Respiración CelularMichel De JerónimoОценок пока нет
- La NeuronaДокумент2 страницыLa NeuronaLiliana MoscolОценок пока нет
- Propanolol Any FarmacoДокумент7 страницPropanolol Any FarmacoPedro Emilio LeonОценок пока нет
- DETERMINACION DE LAS CARACTERISTICAS SENSORIALES DE LA CARNE Y EVALUCION BASICA DE SU FRESCURA Lab ListoДокумент6 страницDETERMINACION DE LAS CARACTERISTICAS SENSORIALES DE LA CARNE Y EVALUCION BASICA DE SU FRESCURA Lab ListoKaroLina Patarroyo CardenasОценок пока нет
- AUTISMOДокумент40 страницAUTISMOyvmuhechОценок пока нет
- Capítulo 11 - Anemias & Hemograma - Serie RojaДокумент27 страницCapítulo 11 - Anemias & Hemograma - Serie RojaRichard RosalesОценок пока нет
- Reflejos Primitivos Del Recién NacidoДокумент4 страницыReflejos Primitivos Del Recién Nacidocepeda.sanchez.ivannaОценок пока нет