Вам также может понравиться
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (895)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (399)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (266)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2259)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (121)
- Nursing Management of Pressure Ulcers in Adults PDFДокумент33 страницыNursing Management of Pressure Ulcers in Adults PDFAnindi Vivia MuktitamaОценок пока нет
- Anatomy, Head and Neck, Temporomandibular Joint - StatPearls - NCBI BookshelfДокумент8 страницAnatomy, Head and Neck, Temporomandibular Joint - StatPearls - NCBI BookshelfDrishti ShahОценок пока нет
- 1st SEMESTER MIDTERM REVIEWER in MEDICAL SURGICAL NURSINGДокумент4 страницы1st SEMESTER MIDTERM REVIEWER in MEDICAL SURGICAL NURSINGskoolrkiveОценок пока нет
- Maslows Hierarchy of NeedsДокумент10 страницMaslows Hierarchy of NeedsOne-rl Isa-pn100% (1)
- BOS Correspondences On Chakras and AurasДокумент9 страницBOS Correspondences On Chakras and AurasDaniel EnglishОценок пока нет
- Delfi Spec LimbProtectionSleeveДокумент2 страницыDelfi Spec LimbProtectionSleeveabhishekbmcОценок пока нет
- Acupuntura OcularДокумент4 страницыAcupuntura OcularratamanoОценок пока нет
- Lalpathlab PDFДокумент3 страницыLalpathlab PDFTushar Kar0% (1)
- WEEK 1 - Introduction To EnzymologyДокумент6 страницWEEK 1 - Introduction To EnzymologyToff GoyenecheaОценок пока нет
- Introduction History and Development of MicrobiologyДокумент17 страницIntroduction History and Development of Microbiologymoses samuelОценок пока нет
- Botany Laboratory Specimens (Bryophytes)Документ40 страницBotany Laboratory Specimens (Bryophytes)lorrainebarandonОценок пока нет
- Biosep Short OverviewДокумент9 страницBiosep Short OverviewMeyakorberОценок пока нет
- Kombinasi Pengaruh Media Tanam Akar Pakis Dan Arang Sekam Terhadap Perkecambahan Dan Pertumbuhan Bibit EucalyptusДокумент9 страницKombinasi Pengaruh Media Tanam Akar Pakis Dan Arang Sekam Terhadap Perkecambahan Dan Pertumbuhan Bibit EucalyptusNeta AdzkiyaОценок пока нет
- Nooij18 Frontiers OverviewVirusMetagenomicClassificationMethodsДокумент21 страницаNooij18 Frontiers OverviewVirusMetagenomicClassificationMethodsAnh H NguyenОценок пока нет
- 1 - UK Lamuno Plus BrochureДокумент2 страницы1 - UK Lamuno Plus BrochurePhyo WaiОценок пока нет
- Chemistry FactsДокумент2 страницыChemistry FactsabcSDОценок пока нет
- Smejkalova - Nikola - 513281 - Bachelor ThesisДокумент86 страницSmejkalova - Nikola - 513281 - Bachelor ThesisShayn J. BenignoОценок пока нет
- Digestive-System-2 Grade 8Документ39 страницDigestive-System-2 Grade 8Joshua Muego100% (2)
- LFSC Class Test GRD 10 2023Документ4 страницыLFSC Class Test GRD 10 2023gugumatlanatoОценок пока нет
- Acid Hydrolysis and Chemical Characterization of DNAДокумент28 страницAcid Hydrolysis and Chemical Characterization of DNAai_ferminОценок пока нет
- Sja 15 435Документ6 страницSja 15 435Nilamsari KurniasihОценок пока нет
- Research Paper On 3D Printed Prosthesis As A Better AlternativeДокумент16 страницResearch Paper On 3D Printed Prosthesis As A Better AlternativeTanbin Rafee100% (1)
- TC18Документ3 страницыTC18Robert BarnesОценок пока нет
- Predicting The Response of Molluscs To The Impact of Ocean AcidificationДокумент43 страницыPredicting The Response of Molluscs To The Impact of Ocean Acidificationmji makОценок пока нет
- Klasmann Peat Substrates Product Information - ForemostCoДокумент32 страницыKlasmann Peat Substrates Product Information - ForemostCoMuhamad FajarОценок пока нет
- Plant Cell Physiology: Osmosis, Turgidity, PlasmolysisДокумент9 страницPlant Cell Physiology: Osmosis, Turgidity, PlasmolysisAli SulaimanОценок пока нет
- 1.pfizer Documents Reveal at Least 800 People Never Finished The COVID Vaccine Trial Due To Death, Injury or Withdrawn Consent - The ExposeДокумент15 страниц1.pfizer Documents Reveal at Least 800 People Never Finished The COVID Vaccine Trial Due To Death, Injury or Withdrawn Consent - The ExposeElvaОценок пока нет
- GenBio 1 Lesson 1 ATP and ADP CycleДокумент2 страницыGenBio 1 Lesson 1 ATP and ADP CycleLlaban MunicaОценок пока нет
- Endocrine System: QuestionsДокумент15 страницEndocrine System: QuestionsJeane Irish Paller EgotОценок пока нет
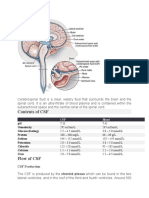
- Cerebrospinal FluidДокумент3 страницыCerebrospinal FluidlecturioОценок пока нет