Вам также может понравиться
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5795)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (895)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (345)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (121)
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- Pediatric Nursing ReviewДокумент45 страницPediatric Nursing Reviewɹǝʍdןnos98% (87)
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- Unit 1 TestДокумент5 страницUnit 1 Testapi-485795043Оценок пока нет
- Essential Variables API 1104Документ4 страницыEssential Variables API 1104Madidj_2014Оценок пока нет
- General Chemistry 2: 2 Semester - Module 2 Types of Solids, Phase Changes and DiagramsДокумент20 страницGeneral Chemistry 2: 2 Semester - Module 2 Types of Solids, Phase Changes and DiagramsKrisha Mae Oao Calumpit100% (2)
- A Brief Introduction To MotivationДокумент16 страницA Brief Introduction To MotivationJoanna Mary LopezОценок пока нет
- Automata, Grammars and Languages: Example: We describe a finite state machine with two input symbols (σ, σ) ,Документ4 страницыAutomata, Grammars and Languages: Example: We describe a finite state machine with two input symbols (σ, σ) ,Joanna Mary LopezОценок пока нет
- Junior Scholastic Dec 13, 2002 105, 9 Proquest CentralДокумент3 страницыJunior Scholastic Dec 13, 2002 105, 9 Proquest CentralJoanna Mary LopezОценок пока нет
- Three Words, Eight Letters, Say It and I'm YoursДокумент387 страницThree Words, Eight Letters, Say It and I'm YoursJoanna Mary Lopez100% (1)
- Pure SegmentationДокумент15 страницPure SegmentationJoanna Mary LopezОценок пока нет
- 1 Placer - Seniors 2 Placer - Grade 7: ST NDДокумент2 страницы1 Placer - Seniors 2 Placer - Grade 7: ST NDJoanna Mary LopezОценок пока нет
- The Feasibility of Chili (Capsicum Frutescens) and KamiasДокумент2 страницыThe Feasibility of Chili (Capsicum Frutescens) and KamiasJoanna Mary LopezОценок пока нет
- CHORDSДокумент1 страницаCHORDSJoanna Mary LopezОценок пока нет
- Why So Serious? Romanized:: (With Individual Parts)Документ1 страницаWhy So Serious? Romanized:: (With Individual Parts)Joanna Mary LopezОценок пока нет
- Corazon AquinoДокумент5 страницCorazon AquinoJoanna Mary LopezОценок пока нет
- Oplan Sagip Gubat Kung Saan Ipinagbabawal Ang Pagtrotroso Sa Lahat NG Matatandang Gubat Sa PilipinasДокумент1 страницаOplan Sagip Gubat Kung Saan Ipinagbabawal Ang Pagtrotroso Sa Lahat NG Matatandang Gubat Sa PilipinasJoanna Mary LopezОценок пока нет
- 151 AssДокумент3 страницы151 AssJoanna Mary LopezОценок пока нет
- Jayne Nathaly R. Lopez Bscs IvДокумент1 страницаJayne Nathaly R. Lopez Bscs IvJoanna Mary LopezОценок пока нет
- The Philippines A Century Hence (Summary)Документ2 страницыThe Philippines A Century Hence (Summary)Joanna Mary LopezОценок пока нет
- Barangay Health WorkerДокумент1 страницаBarangay Health WorkerJoanna Mary Lopez100% (1)
- Aade 11 Ntce 23Документ10 страницAade 11 Ntce 23Kinni ShenoldОценок пока нет
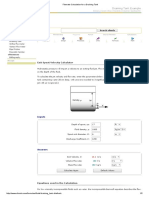
- Flowrate Calculation For A Draining TankДокумент2 страницыFlowrate Calculation For A Draining TankAnonymous bHh1L1Оценок пока нет
- International Journal of Mining, Reclamation and EnvironmentДокумент17 страницInternational Journal of Mining, Reclamation and EnvironmentMargaritha Alexanderina FrancisОценок пока нет
- Is 248-1987 Ra 2021Документ14 страницIs 248-1987 Ra 2021Aadarsh Kumar ShitalОценок пока нет
- ThermodynamicsДокумент10 страницThermodynamicsJeric Nichol DaquizОценок пока нет
- Ultramet 2507: Stainless Steel Electrodes Product Data SheetДокумент1 страницаUltramet 2507: Stainless Steel Electrodes Product Data Sheetshaan1001gbОценок пока нет
- Lesson 2 Leaves - The Food Factory of PlantsДокумент4 страницыLesson 2 Leaves - The Food Factory of PlantsRama Nathan0% (2)
- UNIT VДокумент23 страницыUNIT VDhilsanth SLОценок пока нет
- Two-Phase Total Synthesis of Taxanes: Tactics and StrategiesДокумент28 страницTwo-Phase Total Synthesis of Taxanes: Tactics and StrategiesTạ Đình TrungОценок пока нет
- Flow Boiling and Pool BoilingДокумент9 страницFlow Boiling and Pool BoilingShivani SharmaОценок пока нет
- AssociationДокумент2 страницыAssociationRichmond EresmasОценок пока нет
- Pharmaceutical Industry BR 9108227 en Us 2102 1Документ12 страницPharmaceutical Industry BR 9108227 en Us 2102 1PhongОценок пока нет
- Lithium Ion Battery Overview: by Tony Green Email: Tony@tonygreen - NameДокумент24 страницыLithium Ion Battery Overview: by Tony Green Email: Tony@tonygreen - NameTony GreenОценок пока нет
- 2nd COTdetailedДокумент6 страниц2nd COTdetailedfe delgadoОценок пока нет
- A Brief Review On Optical Properties of Polymer Composites - Insights Into Light-Matter Interaction From Classical To Quantum Transport Point of ViewДокумент28 страницA Brief Review On Optical Properties of Polymer Composites - Insights Into Light-Matter Interaction From Classical To Quantum Transport Point of Viewomkar BembalgeОценок пока нет
- Lydia 1-S2.0-S0022391320301669-MainДокумент9 страницLydia 1-S2.0-S0022391320301669-MainCherifОценок пока нет
- Chapter 13.1 QuestionsДокумент6 страницChapter 13.1 QuestionsLeena BhaiОценок пока нет
- Sci8-Qtr3-Lesn2-Part1-Atomic StructureДокумент21 страницаSci8-Qtr3-Lesn2-Part1-Atomic StructureDaniel LorioОценок пока нет
- MCQ Chapter 2 ElectrochemistryДокумент4 страницыMCQ Chapter 2 ElectrochemistrySavien BrandanОценок пока нет
- Formation Evaluation and Petrophysics MR D. G. BowenДокумент225 страницFormation Evaluation and Petrophysics MR D. G. BowenAimiWani100% (2)
- Exp 10 - Exp 7 A Green Synthesis of Camphor UploadДокумент4 страницыExp 10 - Exp 7 A Green Synthesis of Camphor UploadUyen V. NguyenОценок пока нет
- Solvent Sbps 1425 - HPCLДокумент2 страницыSolvent Sbps 1425 - HPCLBharat ChatrathОценок пока нет
- Iso 9606 4 1999Документ11 страницIso 9606 4 1999Lino Alves InácioОценок пока нет
- TDS Sikasil SG-18 Structural SealantДокумент2 страницыTDS Sikasil SG-18 Structural SealantTuấn Anh NguyễnОценок пока нет
- ETPДокумент3 страницыETPSanaОценок пока нет
- Process Description For Effluent Treatment PlantДокумент2 страницыProcess Description For Effluent Treatment Plantyaxita talati50% (2)
- Aircraft Materials BrochureДокумент6 страницAircraft Materials BrochureAdnan ColoОценок пока нет