Вам также может понравиться
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (895)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (345)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (121)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- 6th Central Pay Commission Salary CalculatorДокумент15 страниц6th Central Pay Commission Salary Calculatorrakhonde100% (436)
- Conf Schedule SmallДокумент46 страницConf Schedule SmallforizslОценок пока нет
- Cauchy Problem As A Two-Surface Based Geometrodynamics 1409.4914Документ35 страницCauchy Problem As A Two-Surface Based Geometrodynamics 1409.4914forizslОценок пока нет
- Distinguishable Consequence of Classical Gravity On Quantum MatterДокумент12 страницDistinguishable Consequence of Classical Gravity On Quantum MatterforizslОценок пока нет
- On Participatory RealismДокумент23 страницыOn Participatory RealismforizslОценок пока нет
- Be Here Now - NatureДокумент1 страницаBe Here Now - NatureforizslОценок пока нет
- A Renormalizable Theory of Quantum Gravity - Renormalization Proof of The Gauge Theory of Volume Preserving Diffeomorphisms 1308.2384Документ37 страницA Renormalizable Theory of Quantum Gravity - Renormalization Proof of The Gauge Theory of Volume Preserving Diffeomorphisms 1308.2384forizslОценок пока нет
- Delirium Quantum - Or, Where I Will Take Quantum Mechanics If It Will Let MeДокумент16 страницDelirium Quantum - Or, Where I Will Take Quantum Mechanics If It Will Let MeforizslОценок пока нет
- A Critique of The Determination of The Energy-Momentum of A System From The Equations of Motion of Matter in The General Theory of RelativityДокумент4 страницыA Critique of The Determination of The Energy-Momentum of A System From The Equations of Motion of Matter in The General Theory of RelativityforizslОценок пока нет
- What Is A Thing - Topos Theory in The Foundations of PhysicsДокумент212 страницWhat Is A Thing - Topos Theory in The Foundations of Physicsforizsl100% (2)
- !mermin - Physics-QBism Puts The Scientist Back Into Science - Nature - 507421a PDFДокумент3 страницы!mermin - Physics-QBism Puts The Scientist Back Into Science - Nature - 507421a PDFforizslОценок пока нет
- Testing Loop Quantum CosmologyДокумент20 страницTesting Loop Quantum CosmologyforizslОценок пока нет
- The 1965 Penrose Singularity TheoremДокумент52 страницыThe 1965 Penrose Singularity TheoremforizslОценок пока нет
- Antimatter and The Second Law of Thermodynamics 1704.01750Документ4 страницыAntimatter and The Second Law of Thermodynamics 1704.01750forizslОценок пока нет
- 136 Keréknyomok 2013 / 7Документ14 страниц136 Keréknyomok 2013 / 7forizslОценок пока нет
- Dīrghatamas, An Application of The Generalization of Witzel's GridДокумент15 страницDīrghatamas, An Application of The Generalization of Witzel's GridforizslОценок пока нет
- Testing Loop Quantum CosmologyДокумент20 страницTesting Loop Quantum CosmologyforizslОценок пока нет
- Modi Ed Chaplygin Gas Cosmology With Bulk Viscosity 1401.8002Документ11 страницModi Ed Chaplygin Gas Cosmology With Bulk Viscosity 1401.8002forizslОценок пока нет
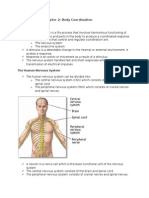
- Form 4 Science Chapter 2Документ13 страницForm 4 Science Chapter 2Cheah Foo Kit50% (2)
- Nervous System Endocrine System and Reproductive SystemДокумент5 страницNervous System Endocrine System and Reproductive SystemMcdaryl Inmenzo Lleno50% (2)
- Artificial Neural Networks - The Tutorial With MatlabДокумент23 страницыArtificial Neural Networks - The Tutorial With MatlabAndrea FieldsОценок пока нет
- NppcityДокумент63 страницыNppcityMohammad JoharОценок пока нет
- The Cambridge Declaration On ConsciousnessДокумент2 страницыThe Cambridge Declaration On ConsciousnessPatrik NazarioОценок пока нет
- Brain Computer Interface/Brain Machine InterfaceДокумент30 страницBrain Computer Interface/Brain Machine InterfaceSiddhant SinghОценок пока нет
- Learning Objectives - Nervous SystemДокумент3 страницыLearning Objectives - Nervous SystemBelle ChenОценок пока нет
- Sensory TortoraДокумент32 страницыSensory TortoraSharifah NadzirahОценок пока нет
- Chap 14Документ16 страницChap 14larryeОценок пока нет
- Proprioceptive Neuromuscular FacilitationДокумент38 страницProprioceptive Neuromuscular FacilitationNolanОценок пока нет
- Rhythm at A Molecular LevelДокумент4 страницыRhythm at A Molecular LevelCMОценок пока нет
- Guyton Somatic SensesДокумент2 страницыGuyton Somatic SensesJamie Ong100% (1)
- Referred PainДокумент9 страницReferred PainElsa Tamara SaragihОценок пока нет
- Neurobiology of The Neuron and The NeurogliaДокумент6 страницNeurobiology of The Neuron and The NeurogliaBarney CordovaОценок пока нет
- Stem Cell Treatment Alzheimer's Disease - ASCI - Asian Stem Cell InstituteДокумент39 страницStem Cell Treatment Alzheimer's Disease - ASCI - Asian Stem Cell Institutea73072Оценок пока нет
- Artificial Neural NetworksДокумент42 страницыArtificial Neural NetworksBhavik PrajapatiОценок пока нет
- Neurophysiology, Neurotransmitters and The Nervous SystemДокумент69 страницNeurophysiology, Neurotransmitters and The Nervous SystemKUNNAMPALLIL GEJO JOHN100% (2)
- ElectroneurogramДокумент10 страницElectroneurogramIrina Luciana DumitriuОценок пока нет
- B1 Nerves Hormones Homeostasis HigherДокумент15 страницB1 Nerves Hormones Homeostasis HigherdownendscienceОценок пока нет
- Perception and Perceptual ProcessДокумент17 страницPerception and Perceptual ProcessDhrub Arka DekaОценок пока нет
- Form 2 Science Mock Test (Sem 1 Term 1)Документ5 страницForm 2 Science Mock Test (Sem 1 Term 1)Spike ChingyenОценок пока нет
- Lesson PlansДокумент33 страницыLesson Plansjaclyn71171% (7)
- Jur Nyeri AlihДокумент11 страницJur Nyeri AlihAchoonk RamadhanОценок пока нет
- Mirror Therapy PosterДокумент1 страницаMirror Therapy Posterapi-335381896Оценок пока нет
- Practice Psych Test 1 QuestionsДокумент18 страницPractice Psych Test 1 Questions46bwilsonОценок пока нет
- Tech RGNL 1 Nerve Conduction Studies KatirjiДокумент52 страницыTech RGNL 1 Nerve Conduction Studies KatirjiSinduja Jayaraj100% (1)
- Lesson 3. Physiological Basis of BehaviorДокумент46 страницLesson 3. Physiological Basis of BehaviorMary Ann DimayugaОценок пока нет
- Physiological Psychology Lecture Notes: Nervous System DamageДокумент3 страницыPhysiological Psychology Lecture Notes: Nervous System DamageGeneric_PersonaОценок пока нет
- Amigdala ( PDFДокумент406 страницAmigdala ( PDFMauricio Escobar CastañedaОценок пока нет
- Embryology of The EarДокумент3 страницыEmbryology of The EarJem MontañaОценок пока нет