Вам также может понравиться
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (895)
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (400)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2259)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (345)
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (121)
- Bifurcation Analysis of An Ecological Food-Chain Model With Switching PredatorДокумент12 страницBifurcation Analysis of An Ecological Food-Chain Model With Switching PredatorAnindita BhattacharyyaОценок пока нет
- Science of Pain PDFДокумент1 089 страницScience of Pain PDFKomang Tri Adi Suparwati100% (1)
- LipidДокумент36 страницLipidSina AgataОценок пока нет
- Non MedelianДокумент29 страницNon MedelianKlaudette Collin PaynorОценок пока нет
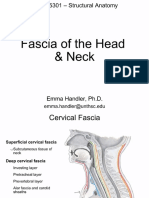
- Fascia of The Head & NeckДокумент8 страницFascia of The Head & NeckMahesh Jung KhadkaОценок пока нет
- 2794-Article Text-8720-1-10-20191103Документ9 страниц2794-Article Text-8720-1-10-20191103blowyour mindОценок пока нет
- Menstruation and FertilizationДокумент54 страницыMenstruation and FertilizationJanaica JuanОценок пока нет
- Unlocking G-Quadruplexes As Antiviral TargetsДокумент27 страницUnlocking G-Quadruplexes As Antiviral TargetsPavanОценок пока нет
- Comparative Analysis of Agrobiological Traits of Durum (T. Durum Desf.) and Bread Wheat (T. Aestivum L.) Varieties in The Karabakh RegionДокумент9 страницComparative Analysis of Agrobiological Traits of Durum (T. Durum Desf.) and Bread Wheat (T. Aestivum L.) Varieties in The Karabakh RegionPublisher NasirОценок пока нет
- CELL CYCLE PresentationДокумент22 страницыCELL CYCLE PresentationCalvin YusopОценок пока нет
- RHS Masterclass On Rose Pruning PDFДокумент3 страницыRHS Masterclass On Rose Pruning PDFCebotari Tatiana50% (2)
- Yearbook 2016Документ308 страницYearbook 2016achintОценок пока нет
- Remediating Motor ControlДокумент66 страницRemediating Motor Controlhis.thunder122100% (3)
- Origin of Eukaryotic Cell PDFДокумент8 страницOrigin of Eukaryotic Cell PDFaco_golarОценок пока нет
- (Brawer) Prostate Specific AntigenДокумент342 страницы(Brawer) Prostate Specific Antigenseeranga_raghuОценок пока нет
- Final IRA - Mangoes From IndiaДокумент174 страницыFinal IRA - Mangoes From Indiasakthi_dtsОценок пока нет
- The Involvement of Hippocampal Neuroinflammation and Oxidative Stress in The Pathogenesis of ASDДокумент13 страницThe Involvement of Hippocampal Neuroinflammation and Oxidative Stress in The Pathogenesis of ASDZeljko LekovicОценок пока нет
- Biology The Unity and Diversity of Life 13th Edition Starr Solutions Manual DownloadДокумент9 страницBiology The Unity and Diversity of Life 13th Edition Starr Solutions Manual DownloadGraham Hissem100% (24)
- QualitativetestforspirinДокумент7 страницQualitativetestforspirinYen BumОценок пока нет
- 1.wall of ThoraxДокумент14 страниц1.wall of ThoraxChandru AОценок пока нет
- Kel LabДокумент92 страницыKel LabAzmi Alvaro PEОценок пока нет
- Report of The 7th International Meeting of The Asian Pacific Society of PeriodontologyДокумент2 страницыReport of The 7th International Meeting of The Asian Pacific Society of PeriodontologyTahir AliОценок пока нет
- Biochemistry of GlycoproteinДокумент6 страницBiochemistry of GlycoproteinMahathir Mohmed100% (7)
- Q4 Week 3 4 GenBio2 EditedДокумент12 страницQ4 Week 3 4 GenBio2 EditedXyreen GalicinaoОценок пока нет
- Ethnic Minorities and Mental HealthДокумент33 страницыEthnic Minorities and Mental HealthAdrianoMascalzoneОценок пока нет
- Mob Ot 31753002739420Документ276 страницMob Ot 31753002739420fabriziozaraОценок пока нет
- Pener1991, Locust Phase Polymorphism and Its EndocrineДокумент79 страницPener1991, Locust Phase Polymorphism and Its EndocrineAdriano Medeiros de SiqueiraОценок пока нет
- Koi On Rearing Pond As ImmunostimulanДокумент18 страницKoi On Rearing Pond As ImmunostimulanDodi SaputraОценок пока нет
- PDFДокумент10 страницPDFmedgadget100% (1)
- Bye Bye Bison PART I Ed 16Документ4 страницыBye Bye Bison PART I Ed 16dquythinh6211Оценок пока нет