Вам также может понравиться
- PH of Common FoodsДокумент2 страницыPH of Common Foodsfisayobabs11Оценок пока нет
- AH FAQ v2.0 Rules & ErrataДокумент15 страницAH FAQ v2.0 Rules & ErrataShum Kah HayОценок пока нет
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (894)
- Blackberry Juice Inbibits Foodborne GrowthДокумент9 страницBlackberry Juice Inbibits Foodborne GrowthzksaraОценок пока нет
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- Effect of Ascorbic Acid Intake On Nonheme-Iron Absorption From A Complete DietДокумент6 страницEffect of Ascorbic Acid Intake On Nonheme-Iron Absorption From A Complete DietsuperyoopyОценок пока нет
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (399)
- Elder Sign FAQ v1.0: Last Updated 8/31/2012Документ5 страницElder Sign FAQ v1.0: Last Updated 8/31/2012zksaraОценок пока нет
- Pathfinder ChecklistДокумент9 страницPathfinder Checklistjuniusgallio100% (5)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- Guidance Notes On Nutrition Labelling: PurposeДокумент20 страницGuidance Notes On Nutrition Labelling: PurposezksaraОценок пока нет
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- AIJNMarketReport2012 PDFДокумент23 страницыAIJNMarketReport2012 PDFzksaraОценок пока нет
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- 2011 June Euro OjДокумент3 страницы2011 June Euro OjzksaraОценок пока нет
- Emerald Neil GaimanДокумент9 страницEmerald Neil GaimanKausik KskОценок пока нет
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- Pathfinder Chronicles Extracted ArtworkДокумент54 страницыPathfinder Chronicles Extracted Artworkzksara100% (5)
- Organic Products and Prices GuideДокумент76 страницOrganic Products and Prices GuidezksaraОценок пока нет
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- VA61 - Gearworld-Rule Book Low ResДокумент24 страницыVA61 - Gearworld-Rule Book Low ReszksaraОценок пока нет
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- SchedaДокумент1 страницаSchedazksaraОценок пока нет
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2219)
- 2011 June Euro OjДокумент3 страницы2011 June Euro OjzksaraОценок пока нет
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- SchedaДокумент1 страницаSchedazksaraОценок пока нет
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (344)
- Amorphous Cellulose - Structure and CharacterizationДокумент9 страницAmorphous Cellulose - Structure and CharacterizationzksaraОценок пока нет
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (265)
- Colorimetric Study of The Post-Processing Effect Due To Pulsed Laser Cleaning of PaperДокумент0 страницColorimetric Study of The Post-Processing Effect Due To Pulsed Laser Cleaning of PaperzksaraОценок пока нет
- Dissolution of Cellulose For Textile Fibre ApplicationsДокумент61 страницаDissolution of Cellulose For Textile Fibre ApplicationszksaraОценок пока нет
- A Transition From Cellulose Swelling To Cellulose Dissolution by O-Phosphoric Acid: Evidence From Enzymatic Hydrolysis and Supramolecular StructureДокумент5 страницA Transition From Cellulose Swelling To Cellulose Dissolution by O-Phosphoric Acid: Evidence From Enzymatic Hydrolysis and Supramolecular StructurezksaraОценок пока нет
- Cocoa Tart With Orange CreamДокумент1 страницаCocoa Tart With Orange CreamzksaraОценок пока нет
- Schroeder, Gentile, Atalla - Nondegradative Preparation of CelluloseДокумент15 страницSchroeder, Gentile, Atalla - Nondegradative Preparation of CellulosezksaraОценок пока нет
- Enemies of BeerДокумент24 страницыEnemies of Beerpana0048100% (1)
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- Posters Whofic 2020Документ107 страницPosters Whofic 2020Kristel HurtadoОценок пока нет
- IWA Publishing - Anaerobic Reactors For Sewage Treatment - Design, Construction and Operation - 2020-01-10Документ1 страницаIWA Publishing - Anaerobic Reactors For Sewage Treatment - Design, Construction and Operation - 2020-01-10JOHNY ALEJANDRO GARCIA SEPULVEDAОценок пока нет
- MDR Guideline Medical Devices LabelingДокумент7 страницMDR Guideline Medical Devices Labelingarade43100% (1)
- Informed Consent and Release, Waiver, and Quitclaim: Know All Men by These PresentsДокумент2 страницыInformed Consent and Release, Waiver, and Quitclaim: Know All Men by These PresentsRobee Camille Desabelle-SumatraОценок пока нет
- Family Nursing Care PlanДокумент1 страницаFamily Nursing Care PlanDersly LaneОценок пока нет
- Hahnemann Advance MethodДокумент2 страницыHahnemann Advance MethodRehan AnisОценок пока нет
- Oferta Hexis - Multisign: Preturile Sunt in EURO Si Nu Includ TVAДокумент9 страницOferta Hexis - Multisign: Preturile Sunt in EURO Si Nu Includ TVAPoschina CiprianОценок пока нет
- Healy Professional DeviceДокумент1 страницаHealy Professional DeviceBramarish KadakuntlaОценок пока нет
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (119)
- Yoga Your Home Practice CompanionДокумент257 страницYoga Your Home Practice Companionjohncoltrane97% (33)
- MsdsДокумент6 страницMsdsJackyОценок пока нет
- Case Study - Genetic DisordersДокумент3 страницыCase Study - Genetic Disordersapi-340003532100% (1)
- 380 Final PaperДокумент46 страниц380 Final Paperapi-538048965Оценок пока нет
- Table : Number of Population, Hospitals and Beds in All Over JordanДокумент8 страницTable : Number of Population, Hospitals and Beds in All Over JordanjОценок пока нет
- Complete VaccinationДокумент2 страницыComplete VaccinationNgoo NwosuОценок пока нет
- HSE List of PublicationsДокумент12 страницHSE List of PublicationsDanijel PindrićОценок пока нет
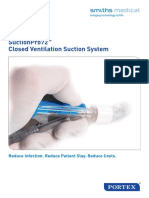
- Brosur Suction Pro 72Документ4 страницыBrosur Suction Pro 72Anonymous tbJ24554Оценок пока нет
- Human Sexual Response Physiology PhasesДокумент2 страницыHuman Sexual Response Physiology PhasesLovely HerreraОценок пока нет
- 2012 Bringing Our Dying HomeДокумент68 страниц2012 Bringing Our Dying HomeendofliferesearchОценок пока нет
- Psychoeducational and Family Therapy in Relapse PreventionДокумент4 страницыPsychoeducational and Family Therapy in Relapse PreventionEdson HilárioОценок пока нет
- The NBCP Referral Code SubДокумент4 страницыThe NBCP Referral Code SubArcon Solite BarbanidaОценок пока нет
- The Helping Art Clinical Nursing Who - Google SeaДокумент1 страницаThe Helping Art Clinical Nursing Who - Google Sea26sbn8d4p9Оценок пока нет
- Birads PosterДокумент1 страницаBirads PosterGopalarathnam BalachandranОценок пока нет
- Everett Association of School Administrators (EASA) Administrative HandbookДокумент46 страницEverett Association of School Administrators (EASA) Administrative HandbookJessica OlsonОценок пока нет
- Hazard Identification Risk Assessment Risk ControlДокумент3 страницыHazard Identification Risk Assessment Risk Controle cubeОценок пока нет
- Stefan White, Andrew Sinclair (Auth.), John M. Hutson, Garry L. Warne, Sonia R. Grover (Eds.) - Disorders of Sex Development_ an Integrated Approach to Management-Springer-Verlag Berlin Heidelberg (20Документ327 страницStefan White, Andrew Sinclair (Auth.), John M. Hutson, Garry L. Warne, Sonia R. Grover (Eds.) - Disorders of Sex Development_ an Integrated Approach to Management-Springer-Verlag Berlin Heidelberg (20Aakanksha MehtaОценок пока нет
- A Medical Outreach Elective CourseДокумент11 страницA Medical Outreach Elective CourseRobert SmithОценок пока нет
- Intro To Wastewater Collection and PumpingДокумент84 страницыIntro To Wastewater Collection and PumpingMoh'd KhadОценок пока нет
- (SEIU) Out Loud Sodexo Workers From Seven Countries Speak Out On Human RightsДокумент30 страниц(SEIU) Out Loud Sodexo Workers From Seven Countries Speak Out On Human RightsNewThorHinoОценок пока нет