Академический Документы
Профессиональный Документы
Культура Документы
Szabó Gábor - Sejtbiológia (2003, 73 Oldal)
Загружено:
Szasz Helen HenriettaОригинальное название
Авторское право
Доступные форматы
Поделиться этим документом
Поделиться или встроить документ
Этот документ был вам полезен?
Это неприемлемый материал?
Пожаловаться на этот документАвторское право:
Доступные форматы
Szabó Gábor - Sejtbiológia (2003, 73 Oldal)
Загружено:
Szasz Helen HenriettaАвторское право:
Доступные форматы
Forrs: http://www.doksi.
hu
SEJTBIOLGIA
Szerkesztette
SZAB GBOR
Forrs: http://www.doksi.hu
Forrs: http://www.doksi.hu
SEJTBIOLGIA
Szerkesztette
SZAB GBOR
MEDICINA KNYVKIAD RT. " BUDAPEST, 2004
Forrs: http://www.doksi.hu
A knyv az Oktatsi Minisztrium tmogatsval, a Felsoktatsi Plyzatok Irodja ltal lebonyoltott Felsoktatsi Tanknyv- s Szakknyvtmogatsi Plyzat keretben jelent meg. Technikai szerkeszt BACS ZSOLT
Dr. Szab Gbor, 2004
E knyv szvege, braanyaga s mindenfle tartozka szerzi jogi oltalom s a kizrlagos kiadi felhasznlsi jog vdelme alatt ll. Csak a szerzi jog tulajdonosnak s a knyv kiadjnak elzetes rsbeli engedlye alapjn jogszer a m egsznek vagy brmely rsznek felhasznlsa, illetve tbbszrzse akr mechanikai, akr fot-, akr elektronikus ton. Ezen engedlyek hinyban mind a msolatkszts, mind a sugrzs vagy a vezetk tjn a nyilvnossghoz val kzvetts, mind a digitalizlt formban val trols, mind a szmtgpes hlzaton tvitt m anyagi formban val megjelentse jogszertlen.
ISBN 963 242 773 4
MEDICINA A kiadsrt felel a Medicina Knyvkiad Rt. igazgatja Felels szerkeszt: dr. Bargr Ilona Mszaki szerkeszt: Bede Tamsn brkat rajzolta: Dr. Bodor Zoltn Bortt tervezte: Varsnyi Gyrgy Terjedelem: (A/5) v Azonost szm: 1754
Forrs: http://www.doksi.hu
A knyv szerzi
BACS ZSOLT
KOVCS JNOS
DAMJANOVICH SNDOR
KOZMA-BOGNR LSZL
DUDA ERN
KRASZNAI ZOLTN
FALUS ANDRS
MADARSZ EMLIA
GSPR REZS
MATK JNOS
GODA KATALIN
MTYUS LSZL
KERESZTES RON
MICZK ANDRS
KISS ANNA
MDIS LSZL
Forrs: http://www.doksi.hu
NAGY FERENC
SZAB GBOR
NAGY LSZL
SZATHMRY ERS
NAGY PTER
SZEBERENYI JZSEF
PAKU SNDOR
SZEKANECZ ZOLTN
PLCZI KATALIN
SZLLSI JNOS
PANYI GYRGY
TIMAR JZSEF
PRDUCZ RPD
UDVARDY ANDOR
RHLICH PL
VEREB GYRGY
SASS MIKLS
VITLIS SNDOR
SIPICZKI MTYS
Forrs: http://www.doksi.hu
Elsz
A knyv sokszerzs volta. Az olvas az els sokszerzs, hazai sejtbiolgia-trgy knyvet tartja a kezben. Tbb, mint harminc klnbz htter, tudomnyos rdeklds szakember, egyetemi oktat vett rszt a knyv megrsban. A sokszerzs jelleg remlt kamatai kztt az egyik legfontosabb az, hogy a knyv ezltal orszgos vllalkozss, a szakma kzs gyv lett. Ennek eredmnyeknt - egy rasztal (szmtgp) helyett - egy virtulis mhelyben szletett ez a knyv, az egyttgondolkods pozitv lmnyben rszestve a szerzket. A munkamegoszts cskkentette az egy szerzre jut anyag terheit, s a munka rmteli szellemi erfeszts maradhatott. Kinek szl a knyv? A gradulis kpzsben rsztvev egyetemi hallgatknak, orvosi s tudomnyegyetemeken egyarnt, posztgradulis kurzusok rsztvevinek s a biolgiai tudomnyok terletn dolgoz kutatknak, kutat orvosoknak, szakterletk ismeretanyagnak tgabb httereknt. Felttelezett elismeretek. Els ves egyetemi hallgatk szmra - ms trgyak elzetes ismerete nlkl is tanulhat a knyv. A sejtbiolgia legfontosabb fejezeteit ttekint bevezets elolvassa utn a tovbbi fejezetek kzpiskols biolgia, fizika, kmia tuds birtokban megrthetek. A nagy, angol nyelv sejtbiolgia knyvekben szoksos rszletes molekulris biolgiai bevezet helyett mi egy rendkvl tmr ttekintsre vllalkoztunk, mely tbb, mint egy sz- ill. fogalom-magyarzat (Glossary), mert sszefggen rja le a legfontosabb s a ksbbiekben visszakszn molekulris fogalmakat, de meghagyja a molekulris biolgia rszletes trgyalst annak a trgynak. Hogyan hasznlhat a gradulis kpzsben? Knyvnk bvebb s mlyebb, rszletesebb, mint ami egy-egy sejtbiolgia-trgy gradulis kurzus cljaira elegend lehet. A Debreceni Egyetem ltalnos Orvosi Karn pl. a flves sejtbiolgia trgy keretben a knyvnek kb. a fele tekinthet trzsanyagnak, kb. a harmada olvasmny, melyeket a hallgatk nll feldolgozsban szeminriumokon beszlnk meg - a fennmarad rszek ms felsoktatsi intzmnyek kurzusai szmra relevnsak. Tanulhatsg. A knyv nagyobb fejezetei szmos alfejezetet tartalmaznak, melyek kzl az els kompliklt anyagrsz esetn a tovbbiak rszletes anyagt rviden s knnyen rtheten ttekinti, a molekulris rszleteket elnagyolva. A tovbbi alfejezetekkel az tfedsek szndkosak, mert elsegtik a knnyebb tanulhatsgot. Br az egyes alfejezetek egymsra plnek, minden (al)fejezet nllan is tanulhat, olvashat. Prbakppen egy-egy fejezetet a bonyolultabbak kzl kiadtunk 10-10 hallgatnak a Debreceni Egyetemen, szeminriumi kiselads anyagaknt. Tanulhatnak, ttekinthetnek bizonyultak. A fejezetek tagolsa. A fejezetek tbbsge egysges tagols s a sejtbiolgia trgynak alapveten ksrletes belltdst, mentalitst tkrzi az ismerettads induktv jelleg. A legtbb fejezet egy biolgiai jelensg, ksrleti megfigyels lersval indul, melyet ahol ennek rtelme van a jelensg felmerl lehetsges magyarzatai kvetnek. Ezt a ma igaznak vlt magyarzat, interpretci kveti, majd az anyag tteles ismertetse (kapcsold ismeretek). A Kitekints c. rszben a mai kutatsi tren-
Forrs: http://www.doksi.hu
ELSZ
dekre ill. a kzeljvben vrhat fejlemnyekre trtnik rvid utals. A fejezeteket ajnlott olvasmnyok rvid listja, sszefoglals s ellenrz krdsek zrjk le. A knyv felptse. A sejt klseje fell haladunk a bels organellumok fel, a rszfunkcik fell a komplexebbek irnyban, az sszefgg ismeretek taglalsa megelzi a fggetlenthet anyagrszekt. Szmos olyan fejezet tallhat knyvnkben, mely az eddig megjelent nagy, angol nyelv kompendiumokbl mg hinyzik. (Ilyen pl. a cirkadin ritmusokrl, az ABC transzporterekrl, a sejten belli diffzis viszonyokrl, a nukleris organizci egyes fejlemnyeirl, az evolci-kutats alapelveirl szl fejezetek, az ionmilivel kapcsolatos tfog ismertets, stb.) Knyvnk szellemisge. Knyvnknek, tbb clkitzst, szmos remlt jellemzjt s fleg sokszerzs mivoltt tekintve, rsos hazai elzmnye nemigen lelhet fel. Termszetesen a korbbi kivl hazai sejtbiolgia knyvek s egyetemi jegyzetek tudsanyaga tovbbrleldve bennnk hozzaddtak knyvnk szellemisghez, melynek gykerei leginkbb abbl a talajbl tpllkoznak, amelyben ott vannak a klasszikus morfolgiai megkzelts soha feleslegess nem vl pedantrija, a molekulris biolgus s genetikus mra bsges nigazolst nyert szemllete s az ltalnos biolgus tgabb horizont szempontjai egyarnt: az egyik fejezetben egyik, a msikban egy msik szempont dominl. Pedaggiai alapelvek. - Vgtelen a tuds birodalma, hasznosabb irnytt adni s megtantani annak hasznlatt, mintsem leltrba venni minden fa, domb nevt s helyt. Nem a konkrt tnyek, akr sszefggsek maradandak, hanem az ismeretek elsajttsnak lmnye, metodikja, ignye s egy minl bvebb sztr (vocabulary), amely segt ebben a tjkozdsban. - Flpasszv tuds, mely elegend ahhoz, hogy a vlaszt egy krdsre a megismert anyagon bell megtalljuk, a szmonkrs jvje. - A tanulst nagyban megknnytik a mr megszerzett ismeretek: ezrt az sszefgg alfejezetek rviden megismtlik a korbbiak legfontosabb lltsait s ezekre ptik sajt mondandjukat. zenet a knyvet olvasknak, tanulknak. A knyvben megtallhatak az sszes szerz elrhetsgi paramterei. Az sszes hasznos javaslatra, ill. szakmai krdsre igyeksznk vlaszolni s az elkvetkez, javtott kiadsokban ezeket figyelembe venni. gy bvlhet virtulis mhelynk igaziv, melynek szellemi produktumban az anyagot elszr olvask s az egyes terleteken sajt tapasztalatokkal rendelkez szakmabeliek szerzk s a szerzk listjn egyelre nem szerepl kollgk gondolatai, keze nyoma mind fellelhetk lesznek. Ksznetnylvnts. A knyv szmos fejezetnek rsakor, brinak szerkesztsekor figyelembe vettk az albbi sejtbiolgiai tematikj kziknyvek, tanknyvek ismeretanyagt: Lodish, H., Matsudaria, P., Baltimore, D., Berk, A., Zipursky, S.L., Darnell, J., Molecular Cell Biology (Scientific American Books, New York, 1999); Alberts, B., Bray, D., Lewis, J., Raff, M., Roberts, K. and Watson, J.D., Molecular Biology of the Cell (Garland Publishing Inc. 1994); B. Alberts, D. Bray, A. Johnson, J. Lewis, M. Raff, K. Roberts, P. Walker, Essential Cell Biology (Garland Publishing, Inc., 1997). Mikroszkpos felvteleket bocstottak rendelkezsnkre Dean A. Jackson (Department of Biomolecular Sciences, UMIST, Manchester, Nagy-Britannia) s Pavel Hozak (Dept. of Cell Ultrastructure and Molecular Biology, Institute of Experimental Medicine, Prga, Csehorszg), melyeket a Company of Biologists LTD. engedlyvel a megfelel fejezetben felhasznltunk, a forrsok feltntetsvel segtsgket ezton is ksznjk. Vgl: ksznm a szerztrsaknak, hogy megtiszteltek egyttmkdskkel, s nevkben is kvnom, hogy sokan, haszonnal s kedvvel olvassk-tanuljk knyvnket. A knyvet desapm emlknek ajnlom. Szab Gbor
Forrs: http://www.doksi.hu
Rvid tartalom
1. 2.
Bevezets a sejtbiolgiba (RHLICH PL) A sejt legfontosabb anyagi sszetevi s alapvet molekulris mechanizmusai. A sejtbiolgia molekulris biolgiai eszkztra (SZAB GBOR s NAGY LSZL) Sejtmembrn s anyagtranszport A sejtmembrn felptse s transzportmechanizmusai (MATK JNOS) Hidrofb anyagok passzv s aktv transzportja. ABC-transzporterek (GODA KATALIN s SZAB GBOR) A membrnpotencil eredete s sejtbiolgiai szerepe. Ioncsatornk s farmakolgiai vonatkozsaik (GSPR REZS) Ionmili - intracellulris kalcium, ozmotikus viszonyok, pH szablyozsa (PANYI GYRGY) A membrn laterlis organizcija (DAMJANOVICH SNDOR) Citoplazmatikus membrnrendszerek s organellumok, intracellulris transzportfolyamatok Endocitotikus s exocitotikus folyamatok, vezikulris transzport: ttekints (KOVCS JNOS) Az endocitotikus/exocitotikus folyamatokat vezrl molekulris trtnsek (NAGY PTER) Kaveolk (KISS ANNA) Endoplazms retikulum (NAGY PTER) Golgi-appartus (NAGY PTER) Lizoszmk (NAGY PTER) Peroxiszmk (NAGY PTER) Fehrjk szelektv clba juttatsa. Fehrjeszorting. Vezikulaszortrozs (NAGY PTER) A sejtmkds energiaforrsa: mitokondrium, kloroplasztisz (SZLLSI JNOS) Diffzis viszonyok a sejtben (BACS ZSOLT) Citoszkeleton: struktrk s funkcik A mikrotubulus, a mikrofilamentum s az intermedier filamentum rendszer (MTYUS LSZL) Sejtkzpont (KOVCS JNOS) Csillk, ostorok (KOVCS JNOS) Molekulris motorok (KOVCS JNOS) Sejtmotilits, kemotaxis (PAKU SNDOR s TIMR JZSEF)
3. 3.1. 3.2. 3.3. 3.4. 3.5. 4. 4.1. 4.2. 4.3. 4.4. 4.5. 4.6. 4.7. 4.8. 4.9. 4.10. 5. 5.1. 5.2. 5.3. 5.4. 5.5.
Forrs: http://www.doksi.hu
10
RVID TARTALOM
6. 6.1. 6.2. 6.3. 6.4. 7. 7.1. 7.2. 7.3. 7.4. 7.5. 7.6.
Sejtmag A sejtmag felptse s funkcii (SZAB GBOR) A nukleocitoplazmatikus makromolekulatranszport (UDVARDY ANDOR) Kromatinszerkezet s a gnexpresszi szablyozsa: molekulris biolgiai mechanizmusok sejtbiolgiai kontextusban (UDVARDY ANDOR) A maghrtya permeabilitsi viszonyai (DAMJANOVICH SNDOR) Sejtosztds Sejtciklus (SZAB GBOR) A mitzis s citokinezis fzisai. A sejtorganellumok sorsa az osztds sorn. A centrilumciklus (SZEBERNYI JZSEF) A sejtosztds mechanikja (SZEBERNYI JZSEF) Meizis (SIPICZKI MTYS) Gametogenezis, fertilizci (KRASZNAI ZOLTN) A sejciklus szablyozsa: biokmiai mechanizmusok sejtbiolgiai kontextusban (UDVARDY ANDOR) A vltoz sejt A sejt tulajdonsgainak megvltozsa kls jelek hatsra: jeltviteli folyamatok (VEREB GYRGY) Jeltviteli jelensgek idegsejteken (PRDUCZ RPD) Jeltvitel fotoreceptorsejtekben (RHLICH PL) Napi ritmusok: az id mint jel (KOZMA-BOGNR LSZL s NAGY FERENC) Sejt s krnyezete Az extracellulris mtrix (MDIS LSZL) Multicellulris szervezds: sejtek kztti s sejt-mtrix kapcsolatok (SZLLSI JNOS) Sejt-vrus interakcik sejtbiolgia vonatkozsai (DUDA ERN) Sejt-baktrium interakcik sejtbiolgiai vonatkozsai (MICZK ANDRS) Sejt-sejt kommunikci s jeltvitel sszefggsei immunsejtekben (FALUS ANDRS) Sejtsorsok s sorsfordulk Sejtsorsok (SZAB GBOR) Sejtsorsok s differencilds (DUDA ERN) Differencilds s szveti regenerci: vrkpz sejtek (PLCZI KATALIN) Mikroorganizmusok differencildsi jelensgei: bacillusok sprzsa (VITLIS SNDOR) A multicellulris szervezds fejldsbiolgiai vonatkozsai (SASS MIKLS) Az egyedfejlds szablyozsnak legfontosabb mozzanatai. A programozott sejthall fejldsbiolgiai vonatkozsai. Sejtadhzis mechanizmusok szerepe az egyedfejlds szablyozsban.
8. 8.1. 8.2. 8.3. 8.4. 9. 9.1. 9.2. 9.3. 9.4. 9.5. 10. 10.1 10.2. 10.3. 10.4. 11. 11.1. 11.2. 11.3.
Forrs: http://www.doksi.hu
RVID TARTALOM
11
12. 13. 13.1 13.2. 13.3. 13.4. 14. 14.1 14.2. 15. 15.1. 15.2.
Az lvilg kialakulsa, evolci. Prokaritk s eukaritk (SZATHMRY ERS) A nvnyi sejt sajtossgai (KERESZTES RON) A nvnyi sejt litikus appartusa A nvnyi sejtvz A sejtfal A nvnyi sejtosztds Sejtbiolgia az orvostudomnyban Sejtbiolgiai ismeretek haszna az orvosi gyakorlatban (SZEKANECZ ZOLTN) Az relmeszeseds sejtbiolgiai mozzanatai (NAGY LSZL) A sejtbiolgia gyakorlata Sejt- s szvettenyszts: mdszertani alapismeretek (MADARSZ EMILIA) Sejtalkotk fluoreszcencis jellse s detektlsa (VEREB GYRGY)
Rvidtsjegyzk Trgymutat
Forrs: http://www.doksi.hu
Forrs: http://www.doksi.hu
Rszletes tartalom
1. 1.1. 1.2. 1.3. 1.3.1. 1.3.1.1. 1.3.1.1.1. 1.3.1.1.2. 1.3.1.1.3. 1.3.1.1.4. 1.3.1.2. 1.3.1.2.1. 1.3.1.2.2. 1.3.1.2.3. 1.3.1.2.4. 1.3.1.2.5. 1.3.1.2.6. 1.3.1.2.7. 1.3.1.2.8. 1.3.1.2.9. 1.3.1.3. 1.3.1.4. 1.3.2. 2.
Bevezets a sejtbiolgiba (RHLICH PL) A sejtbiolgia kialakulsa Nagysgrendek Az l szervezetek alapvet kategrii: pro- s eukarita sejtek A prokaritk s eukaritk legfontosabb tulajdonsgai A prokarita sejt szerkezete Sejtburok Sejtmembrn Citoplazma Nukleoid. A bakterilis DNS Az eukarita sejt szerkezete Sejtmembrn Sejtmag Riboszmk Endoplazmatikus retikulum Golgi-appartus Lizoszma Peroxiszma Mitokondrium Sejtvz (citoszkeleton) A nvnyi sejt specifikumai Az egysejt eukaritk specifikumai Az let peremn A sejt legfontosabb anyagi sszetevi s alapvet molekulris mechanizmusai. A sejtbiolgia molekulris biolgiai eszkztra (SZAB GBOR s NAGY LSZL) A sejt legfontosabb anyagi sszetevi s alapvet molekulris mechanizmusai A fehrjk A sejtek energiaforgalmnak kulcsszereplje: az ATP A DNS szerkezete DNS-replikci
2.1. 2.1.1. 2.1.2. 2.1.3. 2.1.4.
Forrs: http://www.doksi.hu
14
RSZLETES TARTALOM
2.1.5. 2.1.6. 2.1.7. 2.1.8. 2.1.9. 2.1.10. 2.1.11. 2.2. 2.2.1. 2.2.2. 3. 3.1. 3.1.1. 3.1.1.1. 3.1.1.2. 3.1.1.3. 3.1.1.4. 3.1.1.5. 3.1.2. 3.1.2.1. 3.1.2.2. 3.1.2.2.1. 3.1.2.2.2. 3.1.2.2.3. 3.1.2.2.4. 3.1.2.2.5. 3.1.2.2.6. 3.2.
DNS-repair DNS -> mRNS (transzkripci), mRNS -> fehrje (transzlci) A naszcens RNS mRNS-s rse A fehrjk posztranszlcis mdostsai s clba juttatsa A gnexpresszi szablyozsa ltalnos szablyozsi alapelvek A DNS heterogenitsa A sejtbiolgia molekulris biolgiai eszkztra Mdszerek Modellllnyek Sejtmembrn s anyagtranszport A sejtmembrn felptse s transzportmechanizmusai (MATK JNOS) A sejtmembrn szerkezete A lipid ketts rteg Membrnfehrjk A membrnfehrjk azonostsa, a szerkezet s funkci vizsglati lehetsgei A membrnfehrjk laterlis mobilitsa, a "folykony mozaik" membrnmodell Membrnmikrodomnek Anyagtranszport a sejtmembrnon keresztl Molekulris diffzi a lipid ketts rtegen keresztl A membrn-transzportfehrjk fbb tpusai Egy jellegzetes uniport: a glukztranszporter A Na+ /K+-ATP-z pumpafehrjk mkdse s funkcionlis jelentsge A Ca2+ -pumpa-fehrjk mkdsi mechanizmusa A H+ -ATP-zok mkdse s jelentsge eukarita sejtekben ABC-transzporterek Szimport s antiport tpus kotranszportrendszerek A hidrofb vegyletek transzportja. ABC-kazetts transzporterek (SZAB GBOR s GODA KATALIN) Hidrofb molekulk passzv transzportja Aktv transzport: ABC-transzporterek Prokaritk, alacsonyabb rend eukaritk s nvnyek ABC-transzporterei Emberi sejtek ABC-transzporterei Pumpamkds emberi ABC-fehrjk Csatorna- vagy csatornaszablyoz emberi ABC-fehrjk
3.2.1. 3.2.2. 3.2.2.1. 3.2.2.2. 3.2.2.2.1. 3.2.2.2.2.
Forrs: http://www.doksi.hu
RSZLETES TARTALOM
15
3.3.
A membrnpotencil eredete s sejtbiolgiai szerepe. Ioncsatornk s farmakolgiai vonatkozsaik (GSPR REZS) A membrnpotencil eredete Egyenslyi potencil: Nernst-egyenlet Diffzis potencil: Goldman-Hodgkin-Katz-egyenlet A membrnpotencil kialakulsa l sejt esetben Az ioncsatornk sokflesge A membrnpotencil s az ioncsatornk szerepe Az ioncsatornk farmakolgiai vonatkozsai Ioncsatorna-aktivits farmakolgiai szeparlsa csatornablokkolkkal A gygyszerek, toxinok s az ioncsatornk klcsnhatsnak mechanizmusa Ionmili - intracellulris kalcium, ozmotikus viszonyok, pH szablyozsa (PANYI GYRGY) Intracellulris Ca2+-homeosztzis Sejtmembrn Intracellulris Ca2+-raktrak: gyorsan reagl kalciumraktrak Rianodinreceptor IP3-receptor Intracellulris Ca2+-raktrak: mitokondrium Intracellulris raktrak: lassan reagl kalciumraktrak A citoszol tulajdonsgai Ca2+-oszcillci A kalciumoszcillcik jelentsge A kalciumjel dekdolsa: kalmodulin A sejtek ozmo- s volumenszablyozsa Sejtek reakcii nem izotnis kzegben Szablyz trfogat cskkens (RVD) Szablyz trfogat nvekeds (RVI) Intracellulris pH-homeosztzis A citoszol savasodst elidz fontosabb mechanizmusok sszefoglalsa A citoszol lgosodst elidz fontosabb mechanizmusok sszefoglalsa A membrn laterlis organizcija (DAMJANOVICH SNDOR) A Fluid Mozaik Membrn Modell Sapkakpzds Receptor-szuperstruktrk
3.3.1. 3.3.1.1. 3.3.1.2. 3.3.1.3. 3.3.2. 3.3.3. 3.3.4. 3.3.4.1. 3.3.4.2. 3.4.
3.4.1. 3.4.1.1. 3.4.1.2. 3.4.1.3. 3.4.1.4. 3.4.1.5. 3.4.1.6. 3.4.1.7. 3.4.1.8. 3.4.1.9. 3.4.1.10. 3.4.2. 3.4.2.1. 3.4.2.2. 3.4.2.3. 3.4.3. 3.4.3.1. 3.4.3.2. 3.5. 3.5.1. 3.5.2. 3.5.3.
Forrs: http://www.doksi.hu
16
RSZLETES TARTALOM
4.
Citoplazmatikus membrnrendszerek s organellumok, intracellulris transzportfolyamatok Endocitotikus s exocitotikus folyamatok, vezikulris transzport: ttekints (KOVCS JNOS) Az exo- s endocitotikus folyamatok kzs jellemzi A vlogats ltalnos molekulris alapjai A vezikulris transzport A vezikulakpzds s membrnfzi A vezikulk vlogatsa s irnytott transzportja Az egyensly s a kompartimentumok kztti klnbsgek Az endocitzis Fagocitzis Makropinocitzis Klatrinfggetlen endocitzis Az endocitotikus/exocitotikus/vezikulris transzportfolyamatokat vezrl molekulris trtnsek (NAGY PTER) Burkolt vezikulumok kpzdse a receptormedilt endocitzis sorn A vezikulumok kpzdsnek nhny ltalnos jellemzje Burkolt gdr (coated pit) szerepe a receptormedilt endocitzisban A klatrinburk vezikulum lefzdse a donormembrnrl A klatrinburok kapcsoldsa a sejtmembrnhoz s a citoszkeletonhoz A klatrinburok levlsa a vezikulum felsznrl Az alacsony denzits lipoprotein receptormedilt endocitzisa A transzferrin receptormedilt endicitzisa Az epidermlis nvekedsi faktor receptor medilt endocitzisa Nem klatrinburk vezikulumok A nem klatrinbl felpl burok szerepe A COP-burok polimerizcija A COP-burok levlsa a vezikulum felsznrl A szlltand fehrjk kivlasztsnak molekulris mechanizmusa A vezikulumok transzportja: a citoszkeleton s a molekulris motorok szerepe Egyalegysges G-fehrjk szerepe a vezikulris transzportban Vezikulumok fzija a clorganellum membrnjval Kaveolk (KISS ANNA) Morfolgiai jellemzs Biokmiai jellemzs
4.1.
4.1.1. 4.1.2. 4.1.3. 4.1.3.1. 4.1.3.2. 4.1.3.3. 4.1.4. 4.1.5. 4.1.6. 4.1.7. 4.2.
4.2.1. 4.2.1.1. 4.2.1.2. 4.2.1.3. 4.2.1.4. 4.2.1.5. 4.2.1.6. 4.2.1.7. 4.2.1.8. 4.2.2. 4.2.2.1. 4.2.2.2. 4.2.2.3. 4.2.3. 4.2.4. 4.2.5. 4.2.6. 4.3. 4.3.1. 4.3.2.
Forrs: http://www.doksi.hu
RSZLETES TARTALOM
17
4.3.3. 4.3.4. 4.3.5. 4.3.5.1. 4.3.5.1.1. 4.3.5.1.2. 4.3.5.1.3. 4.3.5.1.4. 4.3.5.2. 4.4. 4.4.1. 4.4.2. 4.4.3. 4.4.3.1. 4.4.3.2. 4.4.3.3. 4.4.3.4. 4.4.3.5.
A kaveolk biogenezise A kaveolk lefzdse, internalizcija A kaveolk funkcija A kaveolk szerepe a transzportfolyamatokban Potocitzis Transzcitzis Endocitzis Transzcellulris transzport, lipidszortrozds A kaveolk szerepe a jeltvitelben, szignltranszdukciban Az endoplazmatikus retikulum (NAGY PTER) Az eukarita sejt legkiterjedtebb intracellulris membrnrendszere az endoplazmatikus retikulum A sima felszn endoplazmatikus retikulum A durva felszn endoplazmatikus retikulum szerepe a fehrjeszintzisben A riboszmk a szignlszekvencia segtsgvel ktdnek az endoplazmatikus retikulum felsznhez A fehrjk poszttranszlcis modifikcijnak fontos lpse a glikozilci A glikozilci jelentsge A fehrjk az endoplazmatikus retikulumban veszik fel hromdimenzis szerkezetket A membrnfehrjk egy csoportja glikozil-foszfatidil-inozitol-csoporttal ktdik a membrnhoz A Golgi-appartus (NAGY PTER) A Golgi-appartus felptse A Golgi-appartusban a fehrjk glikozilldnak Fehrjk tovbbtsa a Golgi-appartuson bell A Golgi-appartus cisz- s transz-hlzatnak szerepe a vezikulris transzportfolyamatokban A dinamikus Golgi-appartus Lizoszmk (NAGY PTER) A lizoszmk bontenzimeket tartalmaz savas pH-j intracellulris vezikulumok Hogyan jutnak a lebontand anyagok a lizoszmkba? Hogyan jutnak a lebontst vgz savas hidrolzok a lizoszmkba? Trolsi betegsgek Peroxiszmk (NAGY PTER)
4.5. 4.5.1. 4.5.2. 4.5.3. 4.5.4. 4.5.5. 4.6. 4.6.1. 4.6.2. 4.6.3. 4.6.4. 4.7.
Forrs: http://www.doksi.hu
18
RSZLETES TARTALOM
4.7.1. 4.7.2. 4.7.3. 4.8. 4.8.1. 4.8.2. 4.8.3.
A peroxiszmk oxidatv bontfolyamatok helysznei A peroxiszmk eredete A peroxiszomlis fehrjk szintzise Fehrjk szelektv clba juttatsa. Vezikulaszortrozs (NAGY PTER) A nukleris DNS ltal kdolt fehrjk szintzist kvet transzportja A szignlszekvencik ltalnos jelentsge Bizonyos sejtek plazmamembrnja eltr lipid- s fehrje-sszettel domneket tartalmaz A sejtmkds energiaforrsa: mitokondrium, kloroplasztisz (SZLLSI JNOS) Mitokondrium Az ATP ellltsa aerob sejtekben A mitokondrium szerkezete Kemiozmzis ATP-szintzis A mitokondrium biogenzise Kloroplasztisz s fotoszintzis A kloroplasztisz szerkezete A fotoszintzis reakcii Diffzis viszonyok a sejtben (BACS ZSOLT) Az intracellulris diffzi jelentsge A citoplazmban zajl, diffzifgg esemnyek A citoplazmatikus szabad diffzit befolysol tnyezk Ionok, kis mret molekulk diffzija a citoplazmban Makromolekulk diffzija a citoplazmban Sejtorganellumokon belli szabad diffzi Mitokondriumok Endoplazmatikus retikulum Sejtmag Intracellulris diffzi mrsi lehetsgei: FRAP, FCS Citoszkeleton: struktrk s funkcik A mikrotubulus, a mikrofilament s az intermedier filament rendszer (MTYUS LSZL) Mikrotubulusok
4.9. 4.9.1. 4.9.1.1. 4.9.1.2. 4.9.1.3. 4.9.1.4. 4.9.1.5. 4.9.2. 4.9.2.1. 4.9.2.2. 4.10. 4.10.1. 4.10.1.1. 4.10.1.2. 4.10.2. 4.10.3. 4.10.4. 4.10.4.1. 4.10.4.2. 4.10.4.3. 4.10.5. 5. 5.1.
5.1.1.
Forrs: http://www.doksi.hu
RSZLETES TARTALOM
19
5.1.2. 5.1.2.1. 5.1.3. 5.1.3.1. 5.2. 5.2.1. 5.2.2. 5.3. 5.3.1. 5.4. 5.4.1. 5.4.2. 5.4.2.1. 5.4.2.2. 5.5. 5.5.1. 5.5.1.1. 5.5.1.1.1. 5.5.1.1.2. 5.5.1.2.1.1. 5.5.1.2.1.2. 5.5.1.2. 5.5.2. 5.5.3. 5.5.4. 5.5.4.1. 5.5.4.2. 5.5.4.3. 5.5.4.4. 5.5.4.5.
Mikrofilamentumok Mikrofilament-asszocilt/aktinkt fehrjk Intermedier filamentumok Intermedier filamentum alosztlyok A sejtkzpont (KOVCS JNOS) A sejtkzpont szerkezete s funkcii A centroszmk viselkedse az ivarsejtek kialakulsa s a fertilizci sorn Csillk, ostorok (KOVCS JNOS) A csillk s ostorok szerkezete, funkcii s keletkezse Molekulris motorok - a sejtvz motorfehrji (KOVCS JNOS) Az aktinvzhoz kapcsold motorfehrjk: a miozinok A mikrotubulusokhoz kapcsold motorfehrjk A kinezinek A dineinek Sejtmotilits, kemotaxis (TIMR JZSEF s PAKU SNDOR) A sejtek vndorlsnak szablyozsa Parakrin mechanizmusok Aspecifikus parakrin mechanizmusok Specifikus parakrin mechanizmusok Kemokinek s kemokin receptorok. Motogn citokinek s receptoraik Autokrin szablyozs: az autokrin citokinek s receptoraik A sejtek mozgsnak mechanikja s biokmiai mechanizmusa A sejtek mozgsa s az extracellulris matrix: integrinek, invadopdia s protezok Az egyes sejttpusok mozgsnak sajtossgai Leukocitk Limfoid sejtek Fibroblasztok s hmsejtek: regenerci Endotlsejtek, angiogenezis Daganatsejtek, tumoros progresszi
Forrs: http://www.doksi.hu
20
RSZLETES TARTALOM
6. 6.1. 6.1.1. 6.1.2. 6.1.2.1. 6.1.2.2. 6.1.2.3. 6.1.2.4. 6.1.2.5. 6.1.2.6. 6.1.2.7. 6.1.2.8. 6.1.3. 6.1.3.1. 6.1.3.2. 6.1.4. 6.2. 6.2.1. 6.2.2. 6.2.3. 6.2.4. 6.3. 6.3.1. 6.3.2. 6.3.3. 6.3.4. 6.3.4.1. 6.3.4.2. 6.3.5. 6.3.6. 6.3.6.1. 6.4. 6.4.1.
Sejtmag A sejtmag felptse s funkcii (SZAB GBOR) Magmtrix A kromatin A kromatin szerkezeti hierarchija Eukromatin, heterokromatin Az interfzisos kromoszmk trbeli elhelyezkedse A rendfenntarts eszkzei A metafzisos kromoszma szerkezete Az interfzisos kromatin s a metafzis-kromoszma szerkezete kztti sszefggs Kromatinszerkezet s gnkifejezds Kromatinszerkezet s replikci Intranukleris szuborganellumok Csavarulatos test, PML-test, speckle A sejtmagvacska Maghrtya A nukleocitoplazmatikus makromolekula transzport (UDVARDY ANDOR) A nukleocitoplazmatikus transzport jelentsge A citoplazma s a sejtmag kztti fehrjetranszport mechanizmusa RNS-ek exportja A nukleocitoplazmatikus transzport szablyozsa Kromatinszerkezet s a gnexpresszi szablyozsa: molekulris biolgiai mechanizmusok sejtbiolgiai kontextusban (UDVARDY ANDOR) A kromatin elemi szerkezeti egysge a nukleoszma A nukleoszmk elrendezdse a DNS mentn Magasabb rend kromatinstruktrk A transzkripcionlisan aktv kromatin szerkezete DNz I hiperszenzitv kromatinstruktrk Hisztonacetilci szerepe a transzkripcionlisan aktv kromatinszerkezet kialaktsban Represszv kromatinkomplexek A kromatinszerkezet szerepe a gnexpresszi szablyozsban A magasabb rend kromatinstruktra szerepe a gnexpresszi szablyozsban A maghrtya permeabilitsi viszonyai (DAMJANOVICH SNDOR) A maghrtya tjrhatsga makromolekulk s ionok szmra: passzv transzportfolyamatok
Forrs: http://www.doksi.hu
RSZLETES TARTALOM
21
6.4.2. 6.4.3. 7. 7.1. 7.1.1. 7.1.2. 7.1.2.1. 7.1.2.2. 7.1.2.3. 7.1.3. 7.1.3.1. 7.1.3.2. 7.1.3.3. 7.1.3.4. 7.2. 7.2.1. 7.2.2. 7.2.3. 7.2.3.1. 7.2.3.2. 7.2.3.3. 7.2.3.4. 7.2.3.5. 7.2.3.6. 7.3. 7.3.1. 7.3.2. 7.3.2.1. 7.3.2.2. 7.3.3. 7.3.4. 7.3.5. 7.3.6. 7.4.
Aktv transzportfolyamatok: energetikai megfontolsok Szablyozott ionramls a magmembrnon keresztl: a kalcium-puffok Sejtosztds Sejtciklus (SZAB GBOR) Alapfogalmak A sejtciklus vizsglati lehetsgei ramlsi citometris analzis Sejtek szinkronizlsa Radioaktv nukleozidanalg inkorporci A sejtciklus-szablyozs alapvet vonsai A szablyozs fszerepli Belps a ciklusba A ciklus elhagysa A szablyozs ellenrei A mitzis fzisai (SZEBERNYI JZSEF) A sejtciklus: interfzis s M-fzis A centroszma ciklus Az M-fzis esemnyei Profzis Prometafzis Metafzis Anafzis Telofzis Citokinzis A sejtosztds mechanikja (SZEBERNYI JZSEF) A mitotikus appartusban fellp mozgsok tpusai A centroszma megkettzdse Az utd centroszmk felkszlse a sztvlsra A centroszmk sztvlsa A metafzikus lemez kialakulsa Kromatidvndorls az anafzis A-ban Az osztdsi ors megnylsa az anafzis B-ben Az osztdsi ors eltnse A meizis (SIPICZKI MTYS)
Forrs: http://www.doksi.hu
22
RSZLETES TARTALOM
7.4.1. 7.4.2. 7.4.2.1. 7.4.2.2. 7.4.3. 7.4.4. 7.4.5. 7.4.6. 7.5. 7.5.1. 7.5.1.1. 7.5.1.2. 7.5.1.2.1. 7.5.1.2.2. 7.5.2. 7.5.2.1. 7.5.2.2. 7.5.2.3. 7.5.3. 7.6.
Az ivaros szaporods haploid s diploid nemzedkek vltakozsra pl A meizis a sejtosztds specilis formja A meizis I A meizis II A ni szervezetben a meizis szakaszokban megy vgbe A citoplazma aszimmetrikusan osztdik a petesejt kpzse sorn Az MPF kulcsszerepet jtszik a meizis szablyozsban Az anyai s apai eredet kromoszmk megoszlsa a meizis sorn Gametogenezis, fertilizci (KRASZNAI ZOLTN) Gametogenezis Spermatogenezis Oogenezis Az oogenezis fzisai A zona pellucida kialakulsa s szerkezete Szaporods, fertilizci Az ovulci A spermiumok tja a hvelybl a petevezetig A termkenyts folyamata A meiozis II befejezdse A sejtciklus szablyozsa: biokmiai mechanizmusok sejtbiolgiai kontextusban (UDVARDY ANDOR) A sejtciklus szablyozsnak ltalnos elve A ciklinfgg kinzok ltalnos tulajdonsgai Szablyozott intracellulris fehrjebonts A sejtciklus szablyozsa Saccharomyces cerevisiae-ben G1-fzis S-fzis G2-fzis M-fzis A sejtciklus ellenrzsi pontjai Sejtciklus-szablyozs magasabb rend eukaritkban A vltoz sejt A sejt tulajdonsgainak megvltozsa kls jelek hatsra: jeltviteli folyamatok (VEREB GYRGY) A sejtek kztti jeltvitel szakaszai
7.6.1. 7.6.2. 7.6.3. 7.6.4. 7.6.4.1. 7.6.4.2. 7.6.4.3. 7.6.4.4. 7.6.4.5. 7.6.5. 8. 8.1.
8.1.1.
Forrs: http://www.doksi.hu
RSZLETES TARTALOM
23
8.1.2. 8.1.3. 8.1.4. 8.1.5. 8.1.5.1. 8.1.5.1.1. 8.1.5.1.2. 8.1.5.1.3. 8.1.5.2. 8.1.5.3. 8.1.6. 8.1.6.1. 8.1.6.2. 8.1.6.3. 8.1.6.4. 8.1.7. 8.1.8. 8.1.9. 8.2. 8.2.1. 8.2.2. 8.2.3. 8.3. 8.3.1. 8.3.2. 8.3.3. 8.3.4. 8.3.5. 8.3.6. 8.3.7. 8.3.8. 8.4. 8.4.1. 8.4.2.
A jelmolekulk tja vltoz hosszsg lehet A jeltvitel specificitsnak alapja. A receptor-ligand klcsnhats. Az intracellulris receptorok A sejtfelszni receptorok f kategrii G-proteinhez kapcsolt receptorok 7 transzmembrn domnnel A cAMP mint msodlagos hrviv Gtl s serkent G-proteinek G-proteinek serkentse s gtlsa A Gs- s a Ras fehrje az intracellulris kapcsol fehrjk GTP-z szupercsaldjba tartoznak Tirozinkinz aktivitssal br receptorok A sejtmagba vezet jeltvteli utak Az aktivlt Ras jele egy kinz kaszkdon t a sejtmagba jut A citokinreceptorokhoz kapcsolt JAK kinzok kzvetlenl aktivljk a transzkripcis faktorokat A G-fehrjk is induklhatnak gntrst let s hall jelei A kalcium s a foszfatidil-inozitol rendszer, mint msodlagos hrviv A membrnreceptorok szablyozsa Onkognek s a jeltviteli folyamatok Jeltviteli jelensgek idegsejteken (PRDUCZ RPD) Elektromos szinapszis Kmiai szinapszis Egy nem hagyomnyos neurotranszmitter: a nitrogn-oxid (NO). Jeltvitel fotoreceptorsejtekben (fototranszdukci) (RHLICH PL) A gerincesek retinlis fotoreceptorsejtjnek szerkezete Plcikk s csapok A fotoreceptor-molekula A sttram s a cGMP-fgg kationcsatorna Jeltvitel a fotopigmenttl a kationcsatornig A fototranszdukcis lnc visszatrse a nyugalmi llapotba (relaxci) Jelersts s adaptci Nagy fnyerhz val alkalmazkods, a kalcium szerepe Napi ritmusok: az id mint jel (KOZMA-BOGNR LSZL s NAGY FERENC) A biolgiai ritmusokrl A cirkadin ritmusok jellemzi, kritriumai
Forrs: http://www.doksi.hu
24
RSZLETES TARTALOM
8.4.2.1. 8.4.2.2. 8.4.3. 8.4.4. 8.4.4.1. 8.4.4.2. 8.4.4.3. 9. 9.1. 9.1.1. 9.1.2. 9.1.2.1. 9.1.2.2. 9.1.2.3. 9.1.2.4. 9.1.3. 9.2. 9.2.1. 9.2.2. 9.2.3. 9.2.3.1. 9.2.3.2. 9.3. 9.3.1. 9.3.2. 9.3.3. 9.3.4. 9.4. 9.4.1. 9.4.2. 9.4.3. 9.4.4. 9.5.
A cirkadin ritmusok fenntarti a cirkadin rk A cirkadin rk ltalnos felptse, az ra elemei, defincik A cirkadin ra szerkezete s mkdse prokaritkban (Synechococcus fajok) A cirkadin ra szerkezete s mkdse eukarita modellszervezetekben Gombk: a Neurospora crassa FRQ/WC-1,2 rendszere Ecetmuslica: a TIM/PER rendszer Egr: hasonlsgok s klnbsgek a muslica TIM/PER rendszerhez kpest Sejt s krnyezete Az extracellulris mtrix (MDIS LSZL) Az extracellulris mtrix defincija, szerepe, jelentsge Az ECM fbb komponensei Kollagnek Elasztin Glukzaminogliknok s proteogliknok Glikoproteidek ECM-sejt klcsnhatsok Multicellulris szervezds: sejtek kztti s sejt-mtrix kapcsolatok (SZLLSI JNOS) A sejtek kztti s a sejt-extracellulris mtrix kapcsolatok szerepe, jelentsge Sejt-mtrix kapcsolatok. Multiadhzis fehrjk s receptoraik Sejt-sejt adhzi Adhzis fehrjk Sejtkapcsolatok (junkcik) Sejt-vrus interakcik sejtbiolgia vonatkozsai. (DUDA ERN) A vrusok mint a sejtbiolgiai kutats eszkzei A vrusok bejutsa a sejtbe Sejtpusztt s sejtkml vrusok A vrusok hatsa a sejtek differencildsi folyamataira Sejt-baktrium interakcik (MICZK ANDRS) A baktriumok bejutsa A baktriumok tllsi stratgii A gazdasejtek apoptzisra gyakorolt hats Egyb klcsnhatsok Sejt-sejt kommunikci s jeltvitel sszefggsei immunsejtekben (FALUS ANDRS)
Forrs: http://www.doksi.hu
RSZLETES TARTALOM
25
9.5.1. 9.5.2. 9.5.3. 9.5.3.1. 9.5.3.2. 9.5.4. 9.5.4.1. 9.5.4.1.1. 9.5.4.1.2. 9.5.4.1.3. 9.5.4.1.4. 9.5.4.2. 9.5.4.3. 9.5.4.3.1. 9.5.4.4. 10. 10.1. 10.1.1. 10.1.2. 10.1.3. 10.1.4. 10.1.5. 10.2. 10.2.1. 10.2.2. 10.2.2.1. 10.2.2.2. 10.2.2.3. 10.2.3. 10.2.4. 10.2.5. 10.2.6. 10.3. 10.3.1. 10.3.2. 10.3.3. 10.3.4.
Mire j az immunvlasz? Milyennek kell lennie a jl mkd immunrendszernek? Kihvsok s vlaszok Mit ismer fel az immunrendszer? Antignek Antignfelismers A limfocitaaktivci smja Antignreceptorok felptse s felismersi mintzata B-sejt-receptor- (BCR-) komplex T-sejt-receptor-komplex Az immunolgiai szinapszis Az intracellulris jeltvitel smja BCR s TCR komplexeken t Kostimulcis klcsnhatsok Citokinek s citokinreceptorok klcsnhatsa Citokinek ltal kivltott jeltviteli folyamatok A kimen jel Sejtsorsok s sorsfordulk Sejtsorsok (SZAB GBOR) Sejtsorsok in vivo s in vitro Differencici Proliferci Sejtregeds s immortalits Sejthall Sejtsorsok s differencilds (DUDA ERN) Sorsdnt lpsek A sejtdifferencilds molekulris fszerepli Nvekedsi s differencildsi faktorok s receptoraik Transzkripcis faktorok A sejt krnyezetnek molekulris tnyezi Sejtdifferencilds mestersges krlmnyek kztt A differencildsi folyamat egyirnysga A differencilds ritkn a genom vltozsaival jr egytt. A differencilds zskutci s a daganatok keletkezse Differencilds s szveti regenerci: A vrkpz sejtek (PLCZI KATALIN) A vrkpz rendszer kialakulsa - vrkpzs az embrionlis s felntt szervezetben A vrsvrtest-regenerci A fehrvrsejtrendszer regenercija A thrombocitaregenerci
Forrs: http://www.doksi.hu
26
RSZLETES TARTALOM
10.3.5. 10.4.
A hemopoetikus regenerci modellje: a csontvel-tltets Mikroorganizmusok differencildsi jelensgei: bacillusok sprzsa (VITLIS SNDOR) A sporulci folyamata Csrzs A sporulci szablyozsa Sporulcis -faktorok A transzkripci modulcija Az idbeli szablyozs legfontosabb jellemzi Az alakzatok trbeli formldsnak szablyozsa A multicellulris szervezds fejldsbiolgiai vonatkozsai (SASS MIKLS) Az egyedfejlds szablyozsnak legfontosabb mozzanatai A petesejtek fejldsnek elmletei A mozaikos petesejtek fejldse A regulatv petesejtek fejldse A Xenopus laevis fejldsnek szablyozsa A fejldst meghatroz zenetek a petesejt felptsben A test hossztengelynek determincija A mezoderma kialakulsa ktltembriban Az embrionlis indukci mechanizmusa Specifikus mRNS-ek a vegetatv trflben A mezoderma trbeli mintzatt kialakt indukcis hatsok A morfognek s hatsmechanizmusuk A gasztrulci folyamata A szrke flhold szerepe a gasztrulci sorn Sejttrendezdsek a gasztrulci ideje alatt A gasztrulci szablyozsban szerepet jtsz gnek A kzponti idegrendszer fejldse A neurulci szablyozsnak klasszikus modellje A velcs rz- s mozgatterleteinek kialakulsa A sejtrklds fogalma Az embrionlis determinci A Drosophila melanogaster mutnsok tanulmnyozsnak lehetsge s jelentsge az embriolgiban Az ugrl gnek (mozg genetikai elemek) s ksrleti felhasznlsuk A petesejt ells plusnak (feji rsznek) determincija A htuls plus meghatrozottsga
10.4.1. 10.4.2. 10.4.3. 10.4.3.1. 10.4.3.2. 10.4.3.3. 10.4.3.4. 11. 11.1. 11.1.1. 11.1.1.1. 11.1.1.2. 11.1.2. 11.1.2.1. 11.1.2.2. 11.1.2.3. 11.1.2.3.1. 11.1.2.3.2. 11.1.2.3.3. 11.1.2.3.4. 11.1.2.4. 11.1.2.4.1. 11.1.2.4.2. 11.1.2.4.3. 11.1.2.5. 11.1.2.5.1. 11.1.2.5.2. 11.1.3. 11.1.3.1. 11.1.4. 11.1.4.1. 11.1.4.2. 11.1.4.3.
Forrs: http://www.doksi.hu
RSZLETES TARTALOM
27
11.1.4.4. 11.1.4.5.
A dorzoventrlis testtengely kialakulsa A dorzoventrlis testtengely mentn aktivld gnek hatsa a csralemezek kialakulsra 11.1.4.6. Az anyai hats gnek s a zigotikus gnek fogalma 11.1.4.6.1. A homonom szelvnyezettsg kialakulsnak szablyozsa, a fejldst irnyt gnek hierarchija 11.1.4.6.2. A heteronom szelvnyezettsg ltrejttnek molekulris mechanizmusa 11.1.4.6.2.1. A HOM-komplex tagjainak elfordulsa s szerepe az emlsk s az ember egyedfejldsben 11.2. 11.2.1. 11.2.1.1. 11.2.1.2. 11.2.1.2.1. 11.2.2. 11.2.2.1. 11.2.2.2. 11.2.2.3. 11.2.2.3.1. 11.2.2.3.2. 11.3. 11.3.1. 11.3.1.1. 11.3.1.2. 11.3.2. 11.3.2.1. 11.3.2.2. 11.3.2.3. 11.3.2.4. 11.3.3. 11.3.4. 11.3.4.1. 11.3.4.2. 11.3.4.3. 11.3.4.4. 11.3.4.5. 11.3.4.6. A programozott sejthall fejlodsbiolgiai vonatkozsai A sejtpusztuls kt f, jl megklnbztethet formja Nekrzis Programozott sejthall A programozott sejthallnak kt f sejttani mechanizmust ismerjk A programozott sejtpusztuls szerepe az ontogenezis sorn Gerinctelen llatok A programozott sejthall s szablyozsa rovarokban Sejtpusztuls a gerincesek fejldse sorn A vgtagfejlds Sejtpusztuls az idegrendszer kialakulsa sorn Sejtadhzis mechanizmusok szerepe az egyedfejlds szablyozsban. A soksejtsg kialakulsnak felttelei A differencilis gnaktivlds A soksejtsg kialakulsnak egyb felttelei A sejtadhzis molekulk szerepe a multicellularits kialakulsban A barzdlds kezdetn sejtadhzi nem figyelhet meg Az els sejtkapcsol fehrjk megjelense a barazdlds idejn A tight junction s a gap junction megjelense s funkcija a blasztula llapotban A sejtadhzis molekulk funkcija a gasztrulci sorn A sejttrendezds mechanizmusai A sejtadhzis molekulk szerepe a csralemezek kialakulsban s sejttrendezdsekben Ksrleti eredmnyek in vitro rendszerekben A sejtadhzis molekulk szvetspecifikus megoszlsa az embrionlis szervezetben A kalciumtl fggetlen sejtadhzis molekulk szerepe az egyedfejldsben Az N-CAM hatsa in vitro ksrleti rendszerekben Az egyedfejlds zavarai CAM hinyos mutnsokban Az integrinek funkcija az embriogenesisben
Forrs: http://www.doksi.hu
28
RSZLETES TARTALOM
11.3.5. 11.3.5.1. 11.3.5.2. 11.3.5.3. 11.3.5.4. 12. 12.1. 12.1.1. 12.1.2. 12.1.3. 12.1.4. 12.1.5. 12.1.6. 12.2. 12.2.1. 12.2.2. 12.2.3. 12.2.4. 13. 13.1. 13.1.1. 13.1.2. 13.1.2.1. 13.1.2.2. 13.1.2.3. 13.1.2.4. 13.2. 13.2.1. 13.2.2. 13.3. 13.3.1. 13.3.2. 13.3.3. 13.3.4.
A sejtvndorlsok s szablyozsuk az egyedfejldsi folyamatokban A sejtvndorls nyomon kvetse a vgtagbimb fejldse alatt A velsnc sejtjeinek tja s megtapadsa a szervezet klnbz helyein A sejtvndorls nhny megoldatlan krdse A sejtvndorls mechanizmust magyarz hipotzisek Az lvilg kialakulsa, evolci. Prokaritk s eukaritk (SZATHMRY ERS) A nagylptk evolcis kp talakulsa A kis rRNS klasszikus trzsfja A molekulris trzsfk szerkesztse A trzsfa gykereztetse paralg gnekkel A laterlis gntranszfer (LGT) zavar hatsa Inszercik s delcik a filogenetika szolglatban Az utols kzs s Az eukarita sejt eredete A sejtmag s a mitokondriumok eredete A fagocitzis fontossga Genetikai membrnok s az organellris protein import Organellris eredet gnek a magban: hasznosuls a citoszolban s az organellumban A nvnyi sejt sajtossgai (KERESZTES RON) A nvnyi sejt litikus appartusa A vakulum keletkezse A vakulum funkcii Litikus funkci Raktrozs Ozmoregulci A fehrjetest s a lipidtest keletkezse s lebomlsa A nvnyi sejtvz A mikrotubulusok elrendezdse A mikrofilamentlis rendszer s a sejten belli mozgsok A sejtfal A sejtfal kialakulsa a sejtosztds sorn A sejtfal f komponensei A sejtfal nvekedse A sejtfal, mint kmiai szignlok forrsa
Forrs: http://www.doksi.hu
RSZLETES TARTALOM
29
13.3.5. 13.3.6. 13.3.6.1. 13.3.6.2. 13.3.6.3. 13.4. 14. 14.1. 14.1.1. 14.1.2. 14.1.3. 14.1.4. 14.1.5. 14.2. 14.2.1. 14.2.2. 14.2.3. 14.2.4. 14.2.5. 15. 15.1. 15.1.1. 15.1.2. 15.1.2.1. 15.1.2.2. 15.1.2.3. 15.1.2.3.1. 15.1.2.3.2. 15.1.3. 15.2. 15.2.1. 15.2.1.1.
Adhzis molekulk Transzport-szablyoz sejtfal-sejtmembrn komplexek Hidrofb sejtfal-sejtmembrn komplex Sejtfal-protuberancik Plazmodezmk A nvnyi sejtosztds Sejtbiolgia az orvostudomnyban Sejtbiolgiai ismeretek haszna az orvosi gyakorlatban (SZEKANECZ ZOLTN) Rheumatoid arthritis: a gyullads sejtjei s meditorai Sejtadhzi, adhzis molekulk A gyullads meditorai Angiogenezis Sejtes s molekulris klcsnhatsok s ezek lehetsges terpis befolysolsa Az relmeszeseds sejtbiolgiai mozzanatai (NAGY LSZL) A korai relmeszesedses elvltozs (lzi) kialakulsa A gyullads szerepe Zsrral teli habos sejtek kialalkulsa Fibrotikus plakk kialakulsa Komplex elvltozs (lzi) s trombzis A sejtbiolgia gyakorlata. Sejt- s szvettenyszts: mdszertani alapismeretek (MADARSZ EMILIA) A sejt- s szvettenyszetek tpusai Sejttenyszetek ksztse Primer sejttenyszetek Sejtvonaltenyszetek indtsa Felszni sejttenyszetek A sejtek folyadkkrnyezete A sejtek szilrd krnyezete Sejtszmvltozs a sejttenyszetekben Sejtalkotk fluoreszcencis jellse s detektlsa (VEREB GYRGY) Fehrjk s nukleinsavak jellse specifikusan ktd molekulk segtsgvel Jells antitestek segtsgvel
Forrs: http://www.doksi.hu
30
RSZLETES TARTALOM
15.2.1.2. 15.2.1.3. 15.2.1.4. 15.2.1.5. 15.2.1.6. 15.2.1.7. 15.2.2. 15.2.3. 15.2.4. 15.2.5.
Jells toxinokkal s ms bioaktv vegyletekkel Jells szubsztrttal s liganddal DNS-festkek Szekvenciaspecifikus nukleinsav-prbk Indirekt jellsi mdszerek Egyb, ex vivo jellsi mdszerek A prbk eljuttatsa a clmolekulkhoz Stratgia sejtalkotk jellsre: PDGF-receptorok, F-aktin s a sejtmag egyidej megfigyelse A fluoreszcencis mikroszkpia nhny alkalmazsa A feloldkpessg hatrai s azok tlpse
Rvidtsjegyzk Trgymutat
Forrs: http://www.doksi.hu
1.
1.1. 1.2. 1.3. 1.3.1. 1.3.1.1. 1.3.1.1.1. 1.3.1.1.2. 1.3.1.1.3. 1.3.1.1.4. 1.3.1.2.
Bevezets a sejtbiolgiba
RHLICH PL
A sejtbiolgia kialakulsa Nagysgrendek Az l szervezetek alapvet kategrii: pro- s eukarita sejtek A prokaritk s eukaritk legfontosabb tulajdonsgai A prokarita sejt szerkezete Sejtburok Sejtmembrn Citoplazma Nukleoid. A bakterilis DNS Az eukarita sejt szerkezete
1.3.1.2.1. 1.3.1.2.2. 1.3.1.2.3. 1.3.1.2.4. 1.3.1.2.5. 1.3.1.2.6. 1.3.1.2.7. 1.3.1.2.8. 1.3.1.2.9. 1.3.1.3. 1.3.1.4. 1.3.2.
Sejtmembrn Sejtmag Riboszmk Endoplazmatikus retikulum Golgi-appartus Lizoszma Peroxiszma Mitokondrium Sejtvz (citoszkeleton) A nvnyi sejt specifikumai Az egysejt eukaritk specifikumai Az let peremn
1.1. A sejtbiolgia kialakulsa
A sejt mretnl fogva szabad szemmel nem lthat, ezrt felfedezse, valamint a sejtelmlet kialakulsa szoros kapcsolatban volt az optikai lencsk s azok kombincijbl kialaktott mikroszkpok fejldsvel. A sejt elnevezs a XVII. sz.-ra nylik vissza, amikor Robert Hooke elhalt nvnyi szvetet (paraft) vizsglva abban kis kamrcskkat figyelt meg. Ma mr tudjuk, hogy ezek a kis regek (latinul cella, kicsinytve cellula, grgl kytos vagy cyta) a nvnyi sejteket krlvev cellulzfal ltal kpzett kamrcskk, teht nem a valdi sejteket, hanem azok helyt ltta. Mg abban az vszzadban Antony van Leewenhoek (ejtsd: Lvenhuk) az ltala szerkesztett egyszer mikroszkp segtsgvel valdi, szabad sejteket (pl. magvas vrsvrtesteket, egysejteket, spermiumokat) is meg tudott figyelni. Ezek a megfigyelsek abban az idben kurizumok vol-
tak, nhny kivncsi ember kedvtelsnek szmtottak, s az id mg nem rett meg arra, hogy az let mibenltrl dnt krdseket tegyenek fel. Az a felismers, hogy mind a nvnyi, mind az llati szervezetek sejtekbl s azok termkeibl plnek fel, a XIX. sz. els felig vratott magra. A sejtelmlet vgleges formjban a botanikus Matthias Schleiden s a zoolgus Theodor Schwann nevhez fzdik (1838 s 1839). Br kezdetben mg sokan gondoltk gy, hogy sejtek spontn mdon, pocsolykban, szerves anyagok bomlsbl is kialakulhatnak, a XIX. sz. kzepre nyilvnvalv vlt, hogy j sejtek csak mr meglv sejt osztdsbl keletkeznek (omnis cellula e cellula, Rudolf Virchow, 1855). lesszem kutatk az egyre tkletesed mikroszkpokkal s preparcis eljrsokkal a XIX. sz. vgig a sejtek egyre tbb komponenst fedeztk fel, a sejtmagot, a magvacskt, a mitzis sorn kialakul kromoszmkat, a centriolumot, a mitokondriumo-
Forrs: http://www.doksi.hu
32
BEVEZETS A SEJTBIOLGIBA
kat s a Golgi-appartust. Louis Pasteur s Robert Koch munkssga nyomn fny derlt arra is, hogy szmos fertz betegsg, elhalt szervezetek lebomlsnak htterben csrk, a mikroszkpban mg ppen lthat baktriumok llnak, s ezzel a prokarita sejtet is felfedeztk. Kiderlt, hogy a petesejt s a spermium egyeslsbl keletkez j egyed sejtek osztdsbl s specializldsbl fejldik ki, tovbb, hogy a szervezet kros mkdse is sejtek hibs mkdsre vezethet vissza. Egyre nyilvnvalbb vlt, hogy a legtbb biolgiai problma htterben sejtes folyamatok llnak, s az let alapvet jelensgeit magnak a sejtnek a megismersvel rthetjk meg. Az gy ltrejv tudomny, a sejttan vagy citolgia a XX. sz. elejn mg nagyrszt ellt s megfestett sejtek, szvetek morfolgiai megfigyelseire alapozdott. Fokozatosan ntt azonban az rdeklds az l sejtek megfigyelse irnt, ami nemcsak szabadon l egysejtek tanulmnyozsval vlt lehetv, hanem azltal is, hogy soksejt szervezetek szveteibl a sejteket sikerlt kiszabadtani s az letfelttelek biztostsval vegben (in vitro) akr hetekig, hnapokig letben tartani (sejttenyszts). l sejtek megfigyelse vezetett a klnbz mozgstpusok, gy a nvnyi sejtekben tallhat krkrs citoplazmaramls (ciklzis), az amboid mozgs, a csills s ostoros mozgs s az izom-sszehzds alapos megismersre. Lnyeges metodikai elrelpst jelentett, amikor a fziskontraszt-mikroszkp bevezetsvel az l sejtek krvonalai, bels szerkezete a nagyobb optikai kontraszt kvetkeztben jobban eltntek. Ez egyben igazolta, hogy az addig lert sejtkomponensek tbbsge nem preparcis mtermk, hanem az l sejtben valsgosan ltez struktrknak felel meg. Az l sejtekbe ksrletesen, sebszileg is sikerlt beavatkozni (egyes sejtkomponenseket eltvoltani vagy tltetni) igen fnomra hzott vegtk segtsgvel, amelyeket preczis mszerekkel (mikromanipultorokkal) a mikroszkp ellenrzse mellett mozgattak. A sejttudomnynak a mig tart forradalmi fejldse a XX. sz. tvenes veiben kezddtt meg az elektronmikroszkpinak s a biokminak a
citolgiba val bevonulsval. Azltal, hogy az elektronmikroszkp feloldkpessge 2-3 nagysgrenddel jobb, mint a legjobb fnymikroszkp, 100-1000-szeresen kisebb struktrk vltak lthatv, s gy feltrult a sejtek bels szerkezetnek bonyolult vilga (szubmikroszkpos citolgia, ultrastruktra-kutats). Minthogy az elektronmikroszkpban az elektronoknak az thatolkpessge csak igen vkony (30-100 nm) prepartumot tesz lehetv, ki kellett dolgozni azokat a mdszereket, melyekkel ilyen vkony mintk voltak kszthetk. Az esetek tbbsgben kmiailag fixlt, mgyantba gyazott sejtekbl ksztenek igen vkony (ultravkony) metszeteket klnleges mikrotomok (ultramikrotomok) segtsgvel, de gyakran hasznltak feltrt sejtekbl izollt sejtkomponenseket is. Teljesen j dimenzik nyltak meg: feltrult a mr ismert sejtorganellumok (mitokondriumok, centriolum, Golgi-appartus, csillk, maghrtya stb.) bels, sokszor a makromolekulris nagysgrendig terjed szerkezete, de ezen tlmenen egy sor egszen j sejtkomponenst fedeztek fel. Ilyenek: a riboszmk, az endoplazmatikus retikulum, a sejtvz egyes komponensei (mikrofilamentumok, intermedier filamentumok, mikrotubulusok), szllt vezikulk, szinaptikus vezikulk, mikrobolyhok, peroxiszmk, lizoszmk stb. De lthatkk vltak egyes makromolekulris komplexek is, mint a DNS, ribonukleoprotein-rszecskk, fehrje- s egyb makromolekulk, melyek mr tvezetnek a biokmia, molekulris biolgia vilgba. Mg az elektronmikroszkp a sejt szerkezett az egyre kisebb dimenzik, makromolekulk vilgba bontotta le, az egyre gyorsabban fejld biokmia viszont az egyedi szerves molekulk fell ptkezett az egyre komplexebb makromolekulris dimenzik, klcsnhatsok fel. A biokmia s a sejttan kapcsolata akkor vlt szorosabb, mikor az egyes sejtkomponensek molekulris szerkezett, az azokban zajl kmiai folyamatokat kvntk megvizsglni. Ehhez arra volt szksg, hogy egyes sejtkomponenseket minl nagyobb tisztasgban s mennyisgben sikerljn l sejtek tmegbl kinyerni. Ez a cl a sejtek mechanikai feltrsa, majd az egyes komponenseknek az
Forrs: http://www.doksi.hu
NAGYSGRENDEK
33
ultracentrifugban val lepedsi sebessge, ill. anyagsrsge rvn val elvlasztsval vlt lehetv. Jellegzetes pldja ennek az irnyzatnak az a felismers, hogy a sejtes lgzs a mitokondriumokhoz ktdik, s hogy a mitokondriumnak a bels membrnja ebben kulcsszerepet jtszik. Ugyancsak tt eredmnyt hoztak a membrnok molekulris szerkezetre s funkciira vonatkoz vizsglatok. Az igazi forradalmi fejldst azonban a molekulris biolgia kialakulsa s ennek a sejttannal val tvzdse jelentette. Br a XX. sz. huszas-harmincas veiben mr ismert volt, hogy a sejtmag DNS-t tartalmaz, a DNS ketts hlix szerkezete s a nukleotid-triplet kdol szerepe a XX. sz. kzepn derlt ki. Innen rohamosan vezetett az t a gnkifejezds sejttani vonatkozsainak, a kromatin molekulris szerkezetnek, a gnek szablyozsnak, az egyes gneknek a sejt mkdsben betlttt szerepnek a megismershez. Mr az tvenes vek vgn nyilvnvalv vlt, hogy a sejtekkel foglalkoz tudomny nem maradhat a klasszikus sejttan keretei kztt, hanem integrlnia kell mindazokat az alapvet ismereteket, amelyeket egyb alaptudomnyok (biokmia, molekulris biolgia, biofizika, genetika, mikrobiolgia, fejldstan, lettan stb.) a sejtre vonatkozlag szolgltatnak. Az gy ltrejtt integratv, szintetizl tudomny sokkal tfogbb s sokoldalbb a sejttannl, s ezrt megklnbztetsl a sejtbiolgia elnevezst kapta.
szabad szem 1 cm FM EM 1 mm 100 mm 10 mm 1 mm 100 nm 10 nm 1 nm 1 szveti struktrk tlagos sejt (1020 mm) vrsvrtest (7,5 mm) kromoszmk (15 mm) baktriumok (0,11 mm) vrusok (10100 nm) riboszmk (2325 nm) globulris fehrjk (215 nm aminosavak (0,33 nm) proton (H )
+
1/1. bra. Nagysgrendi skla. A fggleges nyilak a szabad szemmel, fnymikroszkppal (FM) s az elektronmikroszkppal (EM) lthat struktrk nagysgterjedelmt mutatjk.
1.2. Nagysgrendek
A sejtek s azok bels szerkezete a fny- s elektronmikroszkpok segtsgvel tehetk lthatv. A laikus szmra szokatlan dimenzikban val eligazods nem knny, s nmi tapasztalatot ignyel, ezt kvnja az 1/1. brn lthat nagysgrendi skla megknnyteni. A leggyakrabban hasznlt mrtkegysgek a mikrometer (m) s a nanometer (nm), ahol a m a millimeter ezredrsze (10-3 mm) s a nm a m ezredrsze (10-3 m, ill. 10-6 mm). Idnknt hasznljk mg
az elektronmikroszkpiban korbban bevezetett mrtkegysget, az angstrmt (), mely a nm tizedrsze (10-1 nm). Az atomris s molekulris dimenziktl a sejtes nagysgrendig haladva a kvetkez fbb irnyrtkeket lehet megadni. A H-atom tmrje 0,1 nm, az aminosavak, cukormolekulk 0,3 nm, a nukleotidok 0,5 nm, a legtbb fehrje nagysga 2 s 15 nm kztt mozog, a DNSmolekula vastagsga 2 nm krl van, a vrusok tmrje 10 s 100 nm kztt van, a riboszmk nagysga 25 nm, a baktriumok, mitokondriumok 0,1-1 m, a sejtmag tlagos tmrje 5 m krl van, a legtbb eukarita sejt nagysga 6 s 50 m kz esik. Az emberi vrsvrtest tlagos tmrje 7,5 m, az emberi spermiumfej 6 m, a legnagyobb emberi sejt a petesejt (150 m), de a madarak (madrtojs) 50 mm vagy annl nagyobb is lehet. Ugyancsak nagy sejtek tallhatk az idegsejtek kztt, ahol pl. a gerincveli mozgatneuron hossza az 1 mtert is elrheti.
Forrs: http://www.doksi.hu
34
BEVEZETS A SEJTBIOLGIBA
Tekintve, hogy a szabad szem feloldkpessge 0,3 mm, a fnymikroszkp 0,2 m, mg a legjobb elektronmikroszkp 0,3 nm, a sejteket s a nagyobb sejtorganellumokat fnymikroszkppal tehetjk lthatv, mg a kisebb organellumok, valamint a nagyobbak finomabb szerkezete s a makromolekulris komplexek vizsglatnak megfelel vizsgleszkze az elektronmikroszkp. Mind a fny-, mind az elektronmikroszkp klnbz vltozatait fejlesztettk ki (pl. fluoreszcencis, polarizcis, fziskontraszt, interferenciakontraszt, lzer psztz konfoklis stb. fnymikroszkpok, valamint a transzmisszis s psztz elektronmikroszkp, ezeket l. a biofizika- s a szvettanknyvekben). A feloldkpessg nem felttlenl jelenti, hogy minden ilyen nagysgrend struktra lthat is a mikroszkpokban, a lthatv ttelhez legtbbszr hozztartoznak a vizsgland sejtet vagy sejtkomponenseket megrz, kontrasztjukat fokoz vagy ket szelektve kimutat specilis preparatv mdszerek is.
baktrium
1/2. bra. A prokariota s eukariota sejt mretnek sszehasonltsa. Az elektronmikroszkpos felvtelen baktriumokat fagocitlt makrofg sejtet ltunk (3200x, Rhlich P. felv.).
1.3. Az l szervezetek alapvet kategrii: pro- s eukaritasejtek
1.3.1. A prokaritk s eukaritk legfontosabb tulajdonsgai
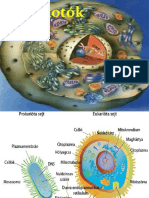
A sejtek szervezdsi fokuk, bonyolultsguk szerint kt alapvet csoportra oszthatk (1/2-3. bra). Az egyik ezek kzl a prokarita sejt, amely arnylag egyszer felpts, kismret sejt (tbbnyire 0,1-1 m), mgis rendelkezik mindazokkal az alapvet struktrkkal s mechanizmusokkal, amelyek az lethez nlklzhetetlenek. Elnevezse onnan szrmazik, hogy nem tallhat benne igazi, maghrtyval rendelkez, a citoplazmtl elklnl sejtmag, hanem csak a mag elfutra (pro = el, karyon = mag, gr.) egy DNS-t tartalmaz terlet formjban (nukleoid). A maghr-
tya hinya mellett szmos ms intracitoplazmatikus membrn, sejtorganellum is hinyzik az ilyen sejtbl, ennek megfelelen felptse igen egyszer. Az alapvet molekulris folyamatok a DNShez, a citoplazmhoz vagy a sejtmembrnhoz kttten zajlanak le. A prokarita sejtek legfontosabb kpviselje a baktriumsejt. A sejtek msik nagy csoportjt alkotjk az eukaritk. Ezek a sejtek jval nagyobbak (6-50 m) s sszetettebbek (1/4-5. bra) a prokaritknl, funkciik is sokkal szertegazbbak, teljestkpessgk jval magasabb szint. Ez jval tbb gnt s ennek megfelelen jval nagyobb DNSllomnyt, genomot ignyel, melynek elhelyezse s a sejtosztds alkalmval val sztosztsa igen nagy feladat el lltja a sejtet. A megolds: az igen hossz DNS egyedi darabokra (kromoszmkra) val tagolsa, a DNS sszecsomagolsa hisztonfehrjkhez val kts ltal (kromatin) s mindennek a citoplazmtl val elhatrolsa maghrtyval. Az eukarita sejt teht valdi sejtmagot tartalmaz (eu = valdi, karyon = mag, gr.). Az eukarita sejt msik fontos jellegzetessge a kompartmentalizci, ami membrnokkal krlhatrolt bels terek (kompartmentek), rekeszek megjelenst jelenti. A terek bels tartalma k-
Forrs: http://www.doksi.hu
AZ L SZERVEZETEK ALAPVET KATEGRII: PRO- S EUKARITASEJTEK
magvacska maghrtya mitokondrium
35
sejtmembrn peptidoglikn rteg komplex membrn kls membrn mezoszma
DNS
plus
nykburok
stt szemcse
muraminburok (szakkulusz)
flagellum
1/4. bra. Eukarita sejt fnymikroszkpos kpe. A fziskontraszt-mikroszkpos felvtelen vegfelsznhez letapadt l makrofg sejtet ltunk, amelyben a sejtmag, maghrtya, magvacska, mitokondriumok s klnbz nagysg stt szemcsk klnbztethetk meg (950x, Rhlich P. felv.).
1/3. bra. Idealizlt baktriumsejt vzlata a fontosabb sejtalkotk bemutatsra. Az bra bal feln Gram-pozitv baktrium burkt, jobb feln pedig Gram-negatv baktrium komplex membrnjt ltjuk.
lnbzik a citoplazmtl, sszettelt az ket hatrol membrn fehrjekszlete hatrozza meg, aminek kvetkeztben a sejten bell bizonyos funkcik kis helyre koncentrldva sokkal hatkonyabbak. Az intracellulris membrnok arnylag nagy fellete szmos enzim s egyb fontos fehrje elhelyezsre ad lehetsget, amelyek a sejt fontos anyagcsere-folyamataiban jtszanak szerepet. Klnbz feladatokra klnbz felpts kompartmentek jttek ltre, ezeket membrnnal
hatrolt sejtorganellumoknak nevezzk (mitokondrium, kloroplasztisz, endoplazmatikus retikulum, Golgi-appartus, lizoszma, peroxiszma, vakulum stb.). A sejtnek a prokarithoz viszonytott lnyegesen nagyobb tmege miatt szksges az alaktarts, ill. annak vltoztatsa, amihez az eukarita sejt egy fonalas elemekbl ll bels vzat fejlesztett ki (sejtvz vagy citoszkeleton). Eukarita sejt minden olyan sejt, amely nem prokarita, teht idetartoznak a protozoonok, az algk, a gombk, a nvnyi s az llati sejtek. A pro- s eukarita sejtek kztt az lvilgban nem ismertek tmenetek, a ktfle sejttpus teht az evolci kt elklnl fejldsi irnyt jelenti.
Forrs: http://www.doksi.hu
36
BEVEZETS A SEJTBIOLGIBA
1/5. bra. Idealizlt eukarita (llati) sejt vzlata a fontosabb sejtkomponensek bemutatsra.
1.3.1.1. A prokarita sejt szerkezete
A legtbb baktrium kismret sejt (0,1-1 m), felptse lnyegben egy membrnnal krlvett citoplazmatmegnek felel meg. Az utbbiban klnll struktrkknt ismerjk fel az elektronmikroszkpban a fehrjeszintzisben fontos szerepet jtsz riboszmkat s a sejt genomjt (rkt llomnyt) kpez DNS-t, tbbnyire a sejt kzponti rszben (nukleoid). A baktriumsejtet hatrol sejtmembrn felsznt klnbz szerkezet burok bortja be, amely a sejt mechanikai stabilitst, ozmotikus erkkel szembeni ellenllkpessgt biztostja. A baktriumsejt ltalnos felptst az 1/3. bra mutatja be. Az egyes komponenseket kvlrl befel haladva trgyaljuk.
1.3.1.1.1. Sejtburok
A sejt felsznt fed burok az egyes baktriumfajtknl eltr szerkezet lehet. Aszerint, hogy egy hagyomnyos festsi mdszerrel (Gram-fests) festdnek-e vagy sem, a baktriumok kt nagy csoportjt szoks megklnbztetni, a Gram-pozitv, ill. -negatv sejteket. A festds minden bizonnyal a baktrium burknak a szerkezeti sajtossgaitl fgg.
A Gram-fests lnyege, hogy a vizsgland ksztmnyt elszr egy bzikus festkkel (gentiana-ibolyval) festik meg, majd jd-kliumjodid oldattal kezelik, s alkohollal differenciljk. A kezels utn egyes baktriumfajtk elszntelednek (Gram-negatv), mg msok megtartjk kkesfekete festdsket (Gram-pozitv).
Forrs: http://www.doksi.hu
AZ L SZERVEZETEK ALAPVET KATEGRII: PRO- S EUKARITASEJTEK
37
a) Muraminsacculus. ltalban a Gram-pozitv baktriumok sejtfelsznt arnylag vastag (2080 nm) fal bortja be, amely hossz sznhidrtlncokbl s rvid sszekt peptidszakaszokbl pl fel. Ezek egymsra fekv ktdimenzis rtegeket alkotnak, melyek mlysgben egymshoz is ktdnek, s gy a burok lnyegben egyetlen risi molekulbl ll, amely zskszeren veszi krbe a sejtet. A sznhidrtlncok vltakozva N-acetil-glukzaminbl s annak tejsavat is tartalmaz szrmazkbl, az N-acetil-muraminsavbl plnek fel. Ezeket a hossz lncokat L- s D-aminosavakbl ll tetrapeptidek s az azokhoz kapcsold glicin-oligopeptidek ktik keresztbe. b) Komplex membrn. A burok msik elterjedt vltozatban a sejtmembrnon kvl, azzal prhuzamosan egy msik membrnszer lipid ketts rteg helyezkedik el. Ennek a kls membrnnak a felsznt sokszor egy fehrjkbl ll szablyos szerkezet rteg (S-rteg) fedi, a membrnba flig beptve pedig lipidbl s sznhidrtbl ll n. lipopoliszacharid (LPS) molekulk tallhatk. A sejtmembrn s a kls membrn kztti igen vkony (2-3 nm) rst (periplazmatikus rs) a muraminburokhoz hasonl szerkezet, de annl lnyegesen vkonyabb peptidogliknrteg tlti ki. Az ebbe a csoportba tartoz baktriumok ltalban Gram-negatvak. Egyb felszni struktrk. a) Nykburok. Egyes baktriumokban a burkot mg egy tovbbi burok fedi, amely kls vgkn szabadon ll, fonlszer, sznhidrtot tartalmaz makromolekulkbl tevdik ssze. Ez a rteg a baktrium felsznnek nykos, viszkzus jelleget ad. b) Ostor, csill (flagellum). Bizonyos baktriumok aktv mozgsra is kpesek, a membrnba beptett ostor mozgatsval. Az ostor fehrjbl (flagellin) felpl, hossz, dughzszeren csavarod, fonlszer kpzdmny, melynek forgatsval a sejt elrehalad mozgst tud ltrehozni. Az ostort a tvnl, a membrnba beptett makromolekulris motor (rotorlemez) forgatja, ener-
gia felhasznlsval. Figyelem: a bakterilis ostor szerkezetileg s a mozgs mechanizmust illeten merben klnbzik az eukarita sejtek csillitl, ostortl (l. ott)! c) Pilusok. Fehrjbl (pilin) felpl, vkony (710 nm), tszer csvek, melyek a baktrium felsznrl nylnak ki. Segtsgkkel a baktrium eukarita sejtek sznhidrtburkt ismeri fel, s ahhoz kapcsoldhat. A pilusok alagtjn keresztl DNS is kicserldhet egyes baktriumok kztt (konjugci), ami egyfajta primitv szexulis eredet genetikai vltozkonysgot is eredmnyez.
1.3.1.1.2. Sejtmembrn
A citoplazmt kvlrl bort membrn, a sejtmembrn, a baktrium egyetlen membrnja (a Gram-negatvak kls membrnjt leszmtva). Vastagsga, molekulris felptsi elve hasonlt az eukarita sejt membrnjhoz (l. ott). Azonkvl, hogy anyagok felvtelben s leadsban, valamint a sejtnek a klvilg fel val elhatrolsban igen fontos szerepe van, a bakterilis membrnban vannak jelen azok a legfontosabb molekulris mechanizmusok is, amelyek az eukarita sejt intracellulris membrnjaira lokalizldnak. Intracellulris membrnok a baktriumokban nincsenek, kivtelt csupn a mezoszma s a cianobaktriumok fotoszintetikus membrnjai kpeznek. Mezoszma. A hosszks sejt kzepe tjn a sejtmembrn begyrdhet a sejt belsejbe, s gy membrnkettzetet alkot, amely nha kiss fel is csavarodik. Ez a mezoszmnak nevezett kettzet teht nem valdi intracellulris membrn, hanem lnyegben a sejtmembrn rsze. Jelentsgt abban ltjk, hogy ide rgzl a bakterilis DNS, s ennek replikcija utn a mezoszma a kt DNS-nek a kt utdsejtbe val sztosztsban jtszik szerepet. Egyes fotoszintetikus baktriumok intracellulris membrnjai. A filogenetikailag si cianobaktriumokban a fotoszintzis molekulris komponensei
Forrs: http://www.doksi.hu
38
BEVEZETS A SEJTBIOLGIBA
intracellulris membrnokban mkdnek. Ennek megfelelen ezen sejtek szli citoplazmjban prhuzamosan elrendezdtt membrnokat tallunk.
1.3.1.1.3. Citoplazma
Elektronmikroszkppal a bakterilis citoplazma tmtt szerkezet s finoman szemcszett. A nagyobb szemcsk a sejt riboszmi, melyek ribonukleinsavbl s fehrjbl llnak, s a fehrjeszintzisben jtszanak fontos szerepet. Valamivel kisebbek s egyszerbbek az eukarita sejt riboszminl. A kzttk lev teret az anyagcsere klnbz, nem membrnhoz kttt enzimei s termkei tltik ki. A citoplazmbl hinyzanak az eukarita sejt jellegzetes sejtorganellumai, gy a mitokondriumok, az endoplazmatikus retikulum, a Golgi-appartus, a centriolum, a sejtvz stb.
Tekintve, hogy a DNS-t a citoplazmtl maghrtya nem vlasztja el, tovbb, hogy a sejt kis mrete miatt minden kis helyre koncentrldik, a gnexpresszi kt lpse: a transzkripci s a transzlci sem klnl el a trben s idben, mint ahogyan az az eukarita sejtben ismert. Ennek megfelelen a gnexpresszi folyamn olyan komplex szerkezetek jnnek ltre, ahol a DNS mentn trd RNS-en mr kzvetlenl zajlik a fehrjeszintzis is. Ezek a sejtbl izollva elektronmikroszkppal lthatv is tehetk. A baktriumok, amilyen egyszerek, olyan gyorsan szaporodnak. Kedvez tpllkozsi s krnyezeti felttelek kztt 20 percenknt osztdnak, ez azt jelenti, hogy 12 ra alatt egyetlen egyedbl sok millird baktrium keletkezik. Ez a gyors szaporodsi kpessg a prokarita sejt legfontosabb letben maradsi stratgija. Amennyiben ez jelents genetikai vltozkonysggal prosul, a baktriumsejt mg a krnyezet drasztikus vltozsait is kpes kivdeni. Br a tenyszet legnagyobb rsze elpusztul, a genetikailag alkalmasabb, jobban adaptld nhny baktrium igen gyorsan elszaporodik, s j, tll populcit alkot. A genetikai vltozkonysgot nemcsak DNS-mutcik biztostjk, hanem egyes baktriumokban a prosods (konjugci) folyamn a plazmidok cserje a piluson keresztl, a fgfertzs folyamn a bakterilis DNS-nek idegen DNS-darabokkal val szennyezdse (transzdukci), tovbb a krnyezetbl szabad DNS-tredkeknek a sejtbe val felvtele (transzformci) is jelentsen hozzjrul a DNS vltozatossghoz. A sejt osztdsa meglehetsen egyszer. Elszr a cirkulris DNS-nek kell replikcival megkettzdnie, majd a sejt befzdssel kettosztdik. A kt cirkulris DNS-t a mezoszma osztja szt a kt klnvl utdsejtbe.
1.3.1.1.4. Nukleoid. A bakterilis DNS
A prokarita DNS olyan ktszl DNS, amelynek nincs kt szabad vge, hanem gyrszeren visszatr nmagba (cirkulris DNS). Tbbnyire a sejt kzepn tallhat meg, elektronmikroszkppal fonalas gombolyag (nukleoid) formjban. A sejtbl kiszabadtva a gyrszer alak elektronmikroszkpban lthatv tehet. A gyrszer DNS-t bakterilis kromoszmnak is szoks nevezni, br az eukarita kromoszmktl mind alakilag (nem lineris, hanem cirkulris), mind pedig szerkezetnek molekulris rszletei miatt elgg tvol ll. Ezen cirkulris kromoszma mellett a prokarita sejtben tovbbi kis cirkulris DNS-ek lehetnek (plazmidok), melyek beplhetnek a nagy kromoszmba, de abbl ki is vlhatnak. Kis tmegarnyuk (2% krl) ellenre fontos informcikat hordozhatnak (pl. Rfaktor, F-faktor stb.). Szemben az eukarita sejttel, a DNS-t maghrtya nem veszi krl, hiszton, egyb csomagol fehrjk s gy kromatin sincs, hinyzik a magvacska is.
Forrs: http://www.doksi.hu
AZ L SZERVEZETEK ALAPVET KATEGRII: PRO- S EUKARITASEJTEK
39
1.3.1.2. Az eukarita sejt szerkezete
Bevezetknt egy idealizlt llati sejten vzlatosan bemutatjuk az eukarita sejt felptst (l. 1/4. bra), majd sorra vesszk az egyes sejtkomponenseket, vgl azokat a klnlegessgeket emltjk meg, amelyek a nvnyi sejtet, ill. az egysejteket az llati sejttl megklnbztetik. A sejtet a klvilgtl vkony hrtya, a sejtmembrn (vagy plazmamembrn) hatrolja el. A sejt tartalmt a hagyomnyos citolgiai elnevezssel protoplazmnak nevezzk, ez a DNS-t tartalmaz s maghrtyval hatrolt sejtmagbl, valamint az azt krlvev citoplazmbl ll. Az utbbi komplex felpts, mert benne szmos, fnyvagy elektronmikroszkppal lthat sejtkomponens, n. sejtorganellum foglal helyet. Az organellumok nagy rsze membrnnal van krlhatrolva, ezek lehetnek egyszer membrnok (endoplazmatikus retikulum, Golgi-appartus, vakulum, lizoszma, peroxiszma, transzportvezikulk esetben) vagy ketts membrnok (mitokondriumokban, nvnyi sejtekben a kloroplasztiszokban). A citoplazma tovbbi organellumai mg a sejtvzat alkot mikrofilamentumok, intermedier filamentumok s mikrotubulusok, tovbb a nagyszm riboszma. Bizonyos sejtekben raktrozott anyagokat, pl. glikognt, zsrcseppeket is tallunk. Az elbb emltett organellumok s tartalkanyagok a citoplazma folykony llomnyba (alapplazma vagy citoszol) begyazva foglalnak helyet. A sejtek felszne klnleges ujjszer nylvnyokat mutathat, ezek a sejtmembrnbl s az ahhoz trsul sejtvzbl jnnek ltre, s merevek vagy mozgkonyak lehetnek (mikrobolyhok, ill. csillk).
1.3.1.2.1. Sejtmembrn
A sejtmembrn (ms nven sejthrtya, plazmamembrn, felszni membrn, (1/6-7. bra) a sejt felsznt bortja be, s ezzel elhatrolja a krnyezettl. Olyan vkony, hogy a rgi citolgusok a fnymikroszkppal kzvetlenl nem lthattk,
s csak egyb jelekbl kvetkeztethettek a jelenltre. Elszr az elektronmikroszkp tette lthatv, amikor is a sejtmembrn keresztmetszetben egy kb. 810 nm vastag vonalnak ltszott. Nagyobb elektronmikroszkpos nagyts az egyetlen vonalat hrom rtegre: kt szls stt s egy kzbls vilgos rtegre tudja felbontani. Ez a 8-10 nm vastag hromrteg (trilaminris) szerkezet jl megfelel annak az elkpzelsnek, hogy a membrn alapszerkezett lipid ketts rteg alkotja, amelyben a hidrofb zsrsavlncok egyms fel fordulnak. A kzps, vilgos rteg ennek megfelelen a zsrsavlncokat reprezentlja, mg a kt szls stt rteg a foszfolipidek fejcsoportjainak felel meg (az elektronmikroszkpos preparatv technika sorn alkalmazott nehzfm atomok, pl. ozmium, urn, lom itt rakdik le, ezrt ad elektronrnykot). A membrnfehrjk jelents rsze (karrierek, ioncsatornk, receptorok, adhzis molekulk, integrinek stb.) a lipid ketts rtegbe van beptve, tbbnyire gy, hogy teljesen tri a membrn vastagsgt. Ezeknek az n. integrns membrnfehrjknek a helyzete annyira stabil, hogy a membrnbl csak igen drasztikus eszkzkkel, a lipid ketts rteg teljes megbontsval nyerhetk ki. Az integrns membrnfehrjknek a membrnon thalad rsze az elektronmikroszkpos kpen nem ltszik, mivel hidrofb tulajdonsg (hidrofb klcsnhats a lipidek zsrsavlncaival!), s gy a membrn kzps, vilgos rtege a metszeti kpen folyamatosnak tnik. Az integrns membrnfehrjk, klnsen a nagyobb tmegek azonban mgis lthatv tehetk egy specilis elektronmikroszkpos preparatv mdszerrel, a fagyasztva trssel. A jgkristlymentesen lefagyasztott szvetdarabot eltrjk, s a trsfelletrl rnykolt replikt ksztnk, majd ezt vizsgljuk az elektronmikroszkpban. A membrnok ezzel az eljrssal hidrofb bels skjukban hasadnak, s az gy szabadd vl hasadsi felleten az integrns membrnfehrjk kis, szemcseszer rszecskkknt (intramembrn rszecskk) jelennek meg. A membrnhoz egyb fehrjk is tartoznak, melyek a membrn kls vagy bels oldal-
Forrs: http://www.doksi.hu
40
BEVEZETS A SEJTBIOLGIBA
membrn sejtburok, alatta a membrn
1/6. bra. A sejtmembrn keresztmetszeti kpe nagy nagyts elektronmikroszkpos felvteleken (Rhlich P. felv.). A) A membrn kt szls stt s az ltaluk kzrefogott vilgos rtegbl ll (trilaminris szerkezet, 287 000x). B) A rutniumvrssel elkezelt sejten a sejtburok is lthatv vlik, mert a rutniumvrs a sejtburok negatv tltseihez ktdik (300 000x).
perifris membrnfehrje oligoszacharidlnc
sejtburok extracellulris felszn
a membrn elektronmikroszkpos kpe
lipid ketts rteg
integrns membrnfehrjk
citoplazmatikus felszn
1/7. bra. A membrn molekulris szerkezetnek modellje.
hoz asszocildnak, tbbnyire egy-egy integrns fehrjhez kapcsoldnak vagy egy hozzjuk kapcsolt zsrsavlnccal lehorgonyzdnak a lipid ketts rtegbe (perifris membrnfehrjk). A sejtmembrnnak az extracellulris tr fel nz felszne gazdag sznhidrtokban. Ezt a sznhidrtds, sokszor a membrn vastagsgt is elr vagy jval meghalad vastagsg rteget sejtburoknak vagy glikokalixnak nevezzk. A sznhidrtok ltalban oligoszacharidlncok formjban fordulnak el, s a membrnfehrjkhez vagy a lipidekhez kapcsoldnak (glikoproteinek, glikoli-
pidek). Sznhidrtok glukzaminogliknokknt is elfordulhatnak, fehrjhez ktve (membrnproteogliknok). A sejtburoknak a sejtmembrn kls oldaln val elhelyezkedse azzal magyarzhat, hogy a glikozilci nagyrszt a Golgi-appartus ciszterninak bels oldaln zajlik, majd miutn az innen levl transzportvezikulk a sejtmembrnnal fuzionltak, a vezikula kinylsval a membrnnak ezen glikozillt rsze az extracellulris tr fel fordul (l. ksbb). A sejtburkot sznhidrtokban s savi csoportokban (szialinsav) val gazdagsga miatt elektronmikroszkpos
Forrs: http://www.doksi.hu
AZ L SZERVEZETEK ALAPVET KATEGRII: PRO- S EUKARITASEJTEK
41
hisztokmival (jelzett, sznhidrtkt tulajdonsg fehrjk, un. lektinek, ill. pozitv tlts fmkolloidok ktse a negatv tlts savi csoportokhoz) tudjuk lthatv tenni. A sejtmembrn jelenlte biztostja azt a diffzis gtat, amely a sejtet elklnti a klvilgtl, s megakadlyozza a kontrolllatlan, szabad diffzit a citoplazma s az extracellulris tr kztt. Ugyanakkor a membrnfehrjk teszik lehetv a sejtnek a klvilggal val kapcsolatt, gy anyagoknak ellenrztt felvtelt vagy leadst, a sejt sajtos bels krnyezetnek a biztostst, ingerek, kls informcik felvtelt, ms sejtekkel val trsulst stb. Mindezek rvn rthet, hogy a sejtmembrn eltvoltsa, nagy rsznek porzuss ttele a sejt pusztulshoz vezet.
1.3.1.2.2. Sejtmag
A DNS-t tartalmaz (1/8-9. bra) sejtmag (nukleusz), arnylagos nagysgnl s jellegzetes festdsi sajtsgainl fogva a sejt legfeltnbb komponense, amelyet mr korn, a XIX. sz. els felben lertak (Boveri, 1831). Alakja szabadon mozg sejtekben tbbnyire gmbly, de gyakran kveti a sejt alakjt is, pl. simaizomsejtben plcika alak, laphmsejtben lelapult, hengerhmsejtben megnylt. A mellette elhelyezked centriolum benyomhatja a mag felsznt, miltal a mag vese alakv vlik, egyes specilis sejtekben pedig a mag tbb helyen befzdhet, s gy lebenyezett, karjozott magv sejtek alakulnak ki. A mag llomnya bzikus (pozitv tlts) festkekkel festhet, teht n. bazofil festdsi sajtsgot mutat. Ennek oka a DNS (neutrlis pH-n mutatott) savi jellege a sok negatv tlts foszftcsoport miatt. A festd llomny, melyet a rgi citolgusok kromatinnak neveztek el, lehet finoman elosztott, laza szerkezet (eukromatin), de alkothat nagy, tmtt, ersen festd tmbket is (heterokromatin). A kromatinnak ez a kiss rgies jelleg fogalma j rtelmet kapott a DNS-llomny tmrtsvel kapcsolatos molekulris felfogs rvn. Tekintettel arra, hogy egy tlagosan 5-10 m nagysg sejtmagban risi hosszsg
(emberi sejt esetben kb. 2 m hossz) DNS-t kell elhelyezni, szksg van a DNS sszecsomagolsra, tmrtsre. A tmrts els szintjt egy 8 db hisztonfehrjbl ll kis (csak elektronmikroszkppal lthat) rszecskre val felcsavarods jelenti (nukleoszma). gy gyngysorszer kpzdmny jn ltre, egy 10 nm vastag lncot alkotva, ahol a gyngyknek a nukleoszmk, a fonlnak a DNS fel nem tekeredett kzti szakasza felel meg. Ezt a laza lncot egy jabb hisztonfehrje hzza ssze hosszban, miltal a nukleoszmk szorosan egyms mg kerlnek. Ez a fonl tovbb tekeredik spirlis formban, ennek rvn egy kb. 30 nm vastag kromatinfonl jn ltre, mely hurkokat alkotva tovbb csomagolja a DNS-t. A kromatinnak a fny- s elektronmikroszkpban lthat szerkezete a kromoszmk klnbz fokban szt- vagy sszetekeredett llapottl, azok trbeli elhelyezkedstl fgg. A lazbb szerkezet (eukromatinban gazdag) sejtmag a DNS kevsb tmr csomagolsa miatt nagyobb trfogat, mg az ersen heterokromatikus magllomny kisebb sejtmagban is elhelyezhet. A heterokromatin tmtt szerkezete akadlyozza a gnek trst a DNS-rl. Az ilyen, a gnexpresszi szempontjbl inaktv terletek a sejt lete sorn vgig megmaradhatnak (konstitutv heterokromatin). Lehetnek azonban olyan heterokromatikus terletek is, amelyek idszakosan fellazulnak, s ezzel lehetv teszik az ennek megfelel DNSszakaszokon a gnkifejezdst (fakultatv heterokromatin). A sejtmagnak egyik jellegzetes komponense a magvacska (nuklelusz), amely egy vagy tbb, tmtt szerkezet rgcske formjban fordul el. A kromatinhoz hasonlan szintn bazofil festds, ez azonban elssorban nem a DNS, hanem az itt felhalmozott RNS kvetkezmnye. Ma mr ismert, hogy ez az RNS a riboszmkba bepl RNS-nek (rRNS) felel meg, ezzel a magvacska a riboszmk kpzdsnek a kzpontja. Az emberi haploid kromoszmakszletben 5 olyan kromoszma tallhat, amelynek bizonyos szakaszn riboszomlis gnek fordulnak el (ez a kromoszma nuklelusz organiztor rgija, NOR). Az ilyen
Forrs: http://www.doksi.hu
42
BEVEZETS A SEJTBIOLGIBA
maghrtya magvacska eukromatin szli heterokromatin durva ER
1/8. bra. Mjsejt magjnak elektronmikroszkpos kpe. A laza kromatinszerkezet (eukromatikus) sejtmag kzepn dominl a magvacska, a kevs heterokromatikus rszlet a maghrtya bels felsznhez tapad hozz (11 600x, Rhlich P. felv.).
citoplazma kls membrn
bels membrn kromatin
1/9. bra. Maghrtya s magprusok. A maghrtya a kls s bels membrnbl s az ltaluk kzrezrt trbl ll. A prusokra nyilak mutatnak r. A prus szln a kt membrn thajlik egymsba, az gy ltrejtt kerek nylsba pruskomplex van beptve. Elektronmikroszkpos felvtel (38 000x, Rhlich P. felv.).
Forrs: http://www.doksi.hu
AZ L SZERVEZETEK ALAPVET KATEGRII: PRO- S EUKARITASEJTEK
43
kromoszmk NOR-szakaszuk mentn sokszor egymshoz trsulnak, s az ott termeld rRNS, valamint riboszma-elalakok krlttk felhalmozdva egyetlen nagy tmeget alkotnak, ez a fnymikroszkppal lthat magvacska. Elfordul azonban, hogy nem mindegyik kromoszma trsul, hanem kln llva tbb magvacskt hoz ltre. Az elektronmikroszkp a magvacska llomnyban finoman granulris, ill. filamentris terleteket tud lthatv tenni. A magvacska tmege a riboszmakpzs intenzitst tkrzi, ezrt olyan sejtekben, ahol ers fehrjeszintzis folyik, s nagy a riboszmk irnti igny, egy vagy tbb nagy nukleluszt figyelhetnk meg. Az ilyen sejteket nagy magvacska mellett (a citoplazma riboszmagazdagsga miatt) bazofil festds citoplazma jellemzi. Eltnik a magvacska a sejtosztds profzisban, amikor a kromoszmk kompaktt vlnak, viszont jra megjelenik a sejtosztds vgn a kromoszmk fellazulsakor. A magvacska nem knnyen klnthet el a mag heterokromatikus terleteitl, ezrt felismersre olyan festseket alkalmazunk, amelyek a DNS-t s az RNS-t klnbz sznben tntetik fel (pl. a metilzld a DNS-t zldre, a pironin az RNS-t vrsre festi meg). A patolgiban rkos sejtek vizsglatban alkalmaznak mg specilis ezstimpregncikat, de a nuklelusz szelektv kimutatsra a legmegbzhatbb eljrs az immuncitokmia, melynek sorn a magvacskban elfordul fehrjket mutatnak ki specifikus ellenanyagokkal. Az eukarita sejtben a sejtmagot a citoplazmtl maghrtya (magmembrn, magburok) vlasztja el. A fnymikroszkpban egyszer vonalnak ltsz maghrtya az elektronmikroszkpban kt, egymssal prhuzamosan fut membrnra bonthat, melyek keskeny rst (perinukleris rs) zrnak kzre. A maghrtya teht a kromatint krlvev lapos zsknak (ciszternnak) felel meg, hasonlan az endoplazmatikus retikulum s a Golgi-appartus ciszternihoz. A durva felszn endoplazmatikus retikulummal (durva ER) ez a ciszterna egyes helyeken kzlekedik is, st a citoplazma fel es felsznhez riboszmk is tapad-
nak. A maghrtynak tartst a bels (a mag bels tere fel nz) oldalhoz rgzl vkony rostos rteg ad (lamina fibrosa, nukleris lamina). Ez a rteg a citoszkeleton 10 nm-es filamentumaihoz tartoz, n. laminfehrjkbl (lamin-A, -B, -C) ll filamentumokbl pl fel. A maghrtya csak a sejtosztds profzisban tnik el, amikor a rostos rteg (a laminok foszforilcijra) lebomlik, s a magmembrn egyedi kis hlyagocskkra, vezikulkra esik szt. Az j maghrtya a telofzisban ezekbl a vezikulkbl ll ssze ismt. A mag s a citoplazma kztt lland kommunikci szksges, melynek rvn a magbl mRNS, tRNS s rRNS, valamint a magvacskban sszeszereldtt riboszmk jutnak ki a citoplazmba, onnan viszont klnbz magfehrjk (hiszton s egyb DNS-csomagol fehrjk, valamint a DNS replikcijban, transzkripcijban, az RNS megktsben s tovbbalaktsban szerepet jtsz fehrjk stb.) s riboszomlis fehrjk szlltdnak be a magba. Ezt a nukleocitoplazmatikus ktirny transzportot a maghrtya kerek nylsai, a magprusok teszik lehetv (l. 1/9. bra). A prus nylsba sokfle fehrjbl felpl, risi molekulris komplex illeszkedik be (pruskomplex), ez biztostja, hogy a magba csak az odatartoz (a maglokalizcis jellel rendelkez) fehrjk jussanak be, s teszi lehetv az RNS-ek, riboszmk kijutst a citoplazmba. A maghrtyt tbbnyire sok szz ilyen prus lyuggatja t, melyek szma sejttpusonknt s a sejt funkcionlis llapottl fggen ersen vltozhat.
1.3.1.2.3. Riboszmk
25 nm nagysg s ezrt csak elektronmikroszkppal lthat rszecskk a citoplazmban. A riboszmk nagy felolds elektronmikroszkppal vizsglva egy nagyobb s egy kisebb alegysgbl tevdnek ssze. Kmiai szerkezetket illetleg mindkt alegysg ribonukleinsavbl (rRNS) s fehrjbl ll, a nagyobb alegysg 3 ribonukleinsav-molekult s kb. 50 fehrjt, a kisebb alegysg pedig egy ribonukleinsavat s kb. 35 fehrjt tartalmaz.
Forrs: http://www.doksi.hu
44
BEVEZETS A SEJTBIOLGIBA
Ez az risi nukleinsavfehrje komplex a fehrjeszintzis (transzlci) kzponti eleme, amelynek legfbb funkcija az aminosavak peptidktsekkel val sszektse, s ezzel a polipeptidlnc kialaktsa. Az (aminosavspecifikus) tRNS-ek ltal odaszlltott aminosavak kzl az mRNS bzissorendjnek megfelelen a soron kvetkezt kivlasztja, s vele a polipeptidlncot tovbbpti. Mindez a molekulris sszetevk (tRNS, tRNS-transzferz enzim, mRNS) riboszma ltali trbeli sszerendezdst, szablyozott klcsnhatsait ignyli. A riboszmk a legtbb sejtben nagy szmban vannak jelen, rszben szabadon fordulnak el a citoplazmban (szabad riboszmk), rszben ktdhetnek az endoplazmatikus retikulum membrnjhoz (membrnhoz kttt riboszmk, l. durva felszn endoplazmatikus retikulum). Klnsen sok van bellk olyan sejtekben, melyek intenzv fehrjeszintzist folytatnak (pl. gyorsan szaporod sejtekben, fehrjt szecernl mirigysejtekben stb.), az ilyen sejtek a fnymikroszkpban a riboszmk nukleinsav-tartalma miatt bazofil festdst mutatnak, teht bzikus (pozitv tlts) festkekkel festdnek.
sima felszn ER). Br a kett tbbnyire kapcsolatban van egymssal, a ktfle megjelensi forma egyben nagyrszt eltr molekulris szerkezetet s sejtbiolgiai funkcit is takar. Durva felszn endoplazmatikus retikulum. Az regrendszert hatrol membrn kls (citoplazmatikus) felsznhez egymshoz kzel, azonos tvolsgban riboszmk ktdnek, melyek kis stt granulumok formjban ltszanak, s az endoplazmatikus retikulum felsznt durvv teszik (rviden durva ER vagy rER az angolszsz nevezktan szerint, l. 1/10-11. bra). A hlzatos regrendszert elgaz, majd jra egyesl csszer elemek, ms helyeken pedig lelapult zskszer kpzdmnyek, n. ciszternk alkotjk. Az utbbiak klnsen olyan sejtekben fordulnak el nagyobb mennyisgben, amelyekben intenzv fehrjeszintzis zajlik. Ilyenkor a ciszternk egymssal prhuzamosan rendezdnek, s a sejt citoplazmjnak jelents trfogatt foglaljk el. Az ilyen rendezett szerkezet, nagy tmeg durva ER-t (a XIX. sz.-ban alkotott s az elektronmikroszkpos citolgiban jra felhasznlt szval) ergasztoplazmnak nevezzk. A citoplazmnak ez a terlete a nagyszm riboszma miatt (ribonukleinsav!) a fnymikroszkpban bzikus festkekkel festdik (bazofil), az ergasztoplazma elnevezst is eredetileg e tulajdonsga miatt kapta. Klnsen fejlett ergasztoplazmt tallunk intenzv fehrjeszintzisre specializlt sejtekben (fehrjeszekrtumot termel mirigysejtekben, mint pl. a hasnylmirigy, a nylmirigyek mirigysejtjeiben, a ktszvet fibroblasztjaiban, fiatal porc- s csontsejtekben, a fog dentinjt termel odontoblasztokban, stb.). A durva ER-ben a riboszmk jelenlte utal f funkcijra, a fehrjeszintzisre. Csak a fehrjknek bizonyos csoportja termeldik itt, mgpedig azok, amelyeket a sejt majd vagy az extracellulris trbe ad le (export vagy szekrcis fehrjk), vagy egy membrnnal hatrolt organellumban, a lizoszmban trol (lizoszomlis fehrjk), illetve amelyek a sejtmembrnba plnek be (membrnfehrjk). Mindegyik esetben a sejt a szintzis sorn keletkez naszcens, mg nem feltekeredett
1.3.1.2.4. Endoplazmatikus retikulum
Ez a membrnnal rendelkez organellum csvecskk (tubulusok) s lelapult zskszer elemek (ciszternk) egymssal kzleked labirintikus rendszerbl ll. Az gy ltrejv hlzatot (retikulum) elektronmikroszkpos vizsglattal a XX. sz. tvenes veiben rtk le, s mivel elszr csak a citoplazma bels, maghoz kzeli rszben (endoplazma) talltk meg, endoplazmatikus retikulumnak (rvidtve ER) neveztk el. Az ER teht egy membrnnal hatrolt labirintikus regrendszer, mely behlzza a citoplazma jelents rszt (1/10-13. bra). Kiterjedse a klnbz sejttpusokban igen vltoz, de az ER ltalban mindig megtallhat. Aszerint, hogy kls felsznhez tapadnak-e riboszmk vagy nem, az elektronmikroszkpban az ER felszne durva vagy sima lehet (durva felszn ER, ill.
Forrs: http://www.doksi.hu
AZ L SZERVEZETEK ALAPVET KATEGRII: PRO- S EUKARITASEJTEK
burkos gdr (coated pit)
45
1/10. bra. Kzepesen fejlett durva felszn endoplazmatikus retikulum. A nhny ciszterna egymssal kzlekedve hlzatot alkot. Elektronmikroszkpos felvtel petefszek tszhmsejtjbl (22 670x, Rhlich P. felv.).
mitokondrium
sejtmag
intermedier filamentumokbl ll kteg
ER ciszterna
1/11. bra. Fejlett durva felszn endoplazmatikus retikulum. A prhuzamosan rendezett ciszternk felsznhez riboszmk tapadnak. Elektronmikroszkpos felvtel prostata hmsejtjbl (22 000x, Rhlich P. felv.).
sejtmag
ER ciszternk
Forrs: http://www.doksi.hu
46
BEVEZETS A SEJTBIOLGIBA
polipeptidlncot juttatja t a membrnon (teljes egszben: export, ill. lizoszomlis fehrjk, illetve rszlegesen: membrnfehrjk esetben). Az itt szintetizld fehrjk teht keresztben, a membrn skjra merlegesen cssznak t a transzlci kzben a membrnon (vektorilis transzlci). A termeld fehrje kezdeti szakaszn tallhat meghatrozott aminosavsorrend (szignlszekvencia) szabja meg, hogy melyek ezek a fehrjk. A szignlszekvencit felismer mechanizmus a riboszmt a durva ER membrnjhoz irnytja. A durva ER-ben a termelt fehrjnek kialakul a majdnem vgleges trbeli alakzata, s mr kezdeti mdosulsokon (pl. az n. N-glikozilci kezdeti szakasza) is keresztlesik. A durva ER klnlegesen talakult ciszternjnak tekinthet a maghrtya (ezrt perinukleris ciszternaknt is emlegetik). Ennek a citoplazma fel nz oldaln mg megtallhatk a riboszmk, a sejtmag rege felli oldaln azonban nem, helyette itt a maghrtya rostos rtege (nukleris lamina) ktdik a membrnhoz. Sima felszn endoplazmatikus retikulum. A sima felszn endoplazmatikus retikulum (sima ER vagy sER) elgaz, majd jra egyesl, 30-100 nm tmrj tubulusok szvevnyes hlzatbl ll (l. 1/12-13. bra). A sejtek tbbsgben arnylag kevss fejlett, ilyenkor az elektronmikroszkpos metszeti kpen csak nhny kerek vagy hosszks, membrnnal hatrolt metszeti kpt lthatjuk. Bizonyos sejttpusokban azonban igen ers fejlettsget rhet el, amikor is a citoplazma jelents terlett a sima ER foglalja el, sokszor olyan mrtkben, hogy a tbbi sejtorganellumot gyszlvn kiszortja magbl. A sima ER nagy membrnfelszne rvn egy sor, membrnhoz kttt enzim funkcijhoz biztostja a szksges intracellulris membrnfelletet. Itt helyezkednek el pl. a lipidek szintzishez szksges enzimek, s ezzel a sima ER tbbek kztt a membrnok lipid ketts rtegnek a kialaktsban mkdik kzre. Ezen ltalnos funkcijn tl a sima ER klnbz egyb folyamatokban is szerepet jtszik. A szteroidhormon-szintzis egyes enzimei pl. a sima ER membrnjhoz kttmitokondrium durva ER maghrtya
sima ER
sejtmag
1/12. bra. Sima felszn endoplazmatikus retikulum. A kp fels felt a retikulum egymssal kapcsolatban ll szablytalan csvecski tltik ki. Elektronmikroszkpos felvtel mjsejtbl (16 000x, Rhlich P. felv.).
tek, s ezrt a szteroidhormon-termel sejtekben (mellkvese-kregllomny sejtjei, a here Leydig-sejtjei, a petefszek srgatestsejtjei) igen kiterjedt sima ER-t tallhatunk. De a szteroidhormonokkal rokon koleszterin szintzise is a sima ER-hez kttt, ennek megfelelen a szintzis legfontosabb helyn, a mjsejtben, fejlett a sima ER. Ezen tlmenen a mj mregtelent funkcijrt felels enzimek is a sima ER membrnjban lokalizldnak, st a glikogn-anyagcsere egyes enzimei (pl. glukz-6-foszfatz) szintn a sima ER membrnjban foglalnak helyet. Az a tny, hogy az izomsejt sima ER-je (az n. szarkoplazmatikus retikulum) fontos Ca2+-trol hely, ahonnan a Ca2+ adott ingerre szablyozottan ramlik ki a citoszol-
Forrs: http://www.doksi.hu
AZ L SZERVEZETEK ALAPVET KATEGRII: PRO- S EUKARITASEJTEK
sima ER
47
durva ER
1/13. bra. A sima s durva felszn endoplazmatikus retikulum kapcsolata (rvid nyilak). A sima ER itt elssorban prhuzamosan rendezett ciszternkbl ll. Elektronmikroszkpos felvtel petefszek srgatest sejtjbl (17 200x, Rhlich P. felv.).
ba, arnylag rgta ismert. jabban kiderlt, hogy nem izomsejtek sima ER-je hasonl feladatot lthat el.
1.3.1.2.5. Golgi-appartus
Ezt a sejtmag kzelben elhelyezked organellumot Camillo Golgi olasz neuroanatmus rta le a XIX/XX. sz. forduljn. Az idegsejtek feltntetsre hasznlhat ezstimpregncis mdszerek kidolgozsa kzben az idegsejt magja krl a citoplazmban ezsttel sttre festd hlzatot figyelt meg (apparato reticolare interno), melyrl kiderlt, hogy ms sejtflesgekben is elfordul. Ezt az organellumot felfedezjrl a ksbbiekben Golgi-appartusnak (Golgi-kszlknek) neveztk el. A XX. sz. els felben tbben ktsgbe vontk a Golgi-appartus ltezst, mivel preparcis mtermknek tartottk, ltt az elektronmikroszkpia igazolta vglegesen, s egyben rta le finomabb felptst (1/14-15. bra). A Golgi-appartus fnymikroszkppal megfigyelt hlzata kisebb egyedi egysgekbl ll ssze,
ezeket diktioszmknak nevezzk. Elektronmikroszkppal vizsglva minden diktioszma prhuzamosan egymsra rakd lapos zskokbl (ciszternkbl) pl fel, melyek szma 3 s 10 kztt van. A Golgi-ciszternk falt membrn alkotja, amely a ciszterna bels tert elhatrolja a citoplazmatikus llomnytl. A diktioszmt alkot ciszternk (az azokban lokalizld enzimek rvn) fontos szerepet jtszanak az endoplazmatikus retikulumban szintetizlt fehrjknek a mdostsaiban. Ehhez termszetesen az szksges, hogy ezek a fehrjk a durva ER-bl eljussanak a Golgi-appartusba, ami az ER-bl lefzd hlyagocskk rvn trtnik. Ezek a transzportvezikulk a diktioszma sejtmaghoz legkzelebb es ciszternjval (cisz-ciszterna) fzionlva, majd onnan jra lefzdve s az ER-be visszatrve ideoda ingznak a durva ER s a diktioszma kztt, s a fehrjket tszlltjk a Golgi-appartusba. Bonyolult molekulris rendszer szablyozza ezt a folyamatot s biztostja azt, hogy valban csak a mdostand fehrjk kerljenek t. Az egymsra rakd ciszternk enzimkszlete s ennek megfelelen a funkcija is klnbz. A fehrje a ciszciszterna fell fokozatosan thalad az egyes kzbls ciszternkon egszen a legutols (transzciszterna), mikzben fokozatos mdosulsokon (mint pl. az N-glikozilci vgs szakasza, tovbb az O-glikolizci, hidroxills, szulfatls, clzott fehrjehasts, stb.) esik keresztl. Megoszlanak a vlemnyek arrl, hogy a fehrje milyen mechanizmus rvn tovbbtdik a diktioszmn keresztl. Az egyik nzet szerint a ciszternk kztt az anyagtovbbtst ide-oda ingz transzportvezikulk vgzik, a msik vlemny szerint viszont maguk a ciszternk toldnak a cisz oldal fell a transz oldal fel, azltal, hogy a cisz oldalon jak keletkeznek, mg a transz oldalon a ciszternk levlnak. A diktioszma legutols, transz-ciszternja, ill. az ehhez csatlakoz, csvecskkbl s szablytalan zskszer elemekbl ll hlzat (transz-Golgi-hlzat, TGN) az a hely, ahonnan a klnbz mdosult fehrjk vgs helykre elosztdnak. Az innen lefzd vezikulk tbbsge a sejtmembrn fel halad, majd azzal sszeolvadva (exocitzis) a szekrcis
Forrs: http://www.doksi.hu
48
BEVEZETS A SEJTBIOLGIBA
lizoszma mitokondrium
Polarizlt felpts, szekrcis hmsejtekben a Golgi-appartus helyzete is irnytott, mgpedig a sejtmagnak azon az oldaln helyezkedik el, amelyik irnyban a szekrci trtnik. gy pl. hasnylmirigyben a mag s az apiklis (a mirigyvgkamra lumene fel es) sejtmembrn kztt, mjsejtben pedig a sejtmagnak az epekapillris fel es oldaln tallhat meg.
1.3.1.2.6. Lizoszma
Golgi-appartus sejtmag
1/14. bra. Golgi-appartus mellkhere hmsejtjbl. Elektronmikroszkpos felvtel (15 000x, Rhlich P. felv.).
fehrjket leadja az extracellulris trbe, ill. a fzi rvn a vezikula membrnja felvevdik a sejtmembrnba, s ezzel a membrnfehrjk is vgs helykre, a sejtmembrnba jutnak. A durva ERGolgi tvonalon halad fehrjk harmadik csoportjt, a lizoszomlis fehrjket kln vezikulk szlltjk vgs helykre, a lizoszmba. A lizoszmba val eltrtsrt a lizoszomlis fehrjket jell mannz-6-foszft jel s az azt felismer s a vezikulk membrnjba beptett receptor felels.
Tbbnyire a Golgi-appartus krnykn tallhat organellumok (1/16. bra), melyeket membrn hatrol el a citoszol fel, belsejket pedig szemcss, stt llomny tlti ki. Az utbbirl a lizoszmk felfedezse utn rvidesen kiderlt, hogy savi pH-n mkd hidrolitikus enzimek tmege. Mivel ezek az enzimek a sejteket felpt szerves molekulk lebontsban jtszanak dnt szerepet, az elektronmikroszkpban lthat stt testecskt lizoszmnak neveztk el (gr. lysein: oldani, soma: testecske). A lizoszmk nagysga tg hatrok kztt vltozik (0,1-1 m, sejttpustl s a sejt funkcionlis llapottl fggen). Leginkbb gmblyded kpzdmnyek, de szablytalan alak, csszeren megnylt lizoszmk is ismeretesek. Egyszer felptsk miatt nem mindig knny ket az elekt-
szekrcis vakulum kondenzl vakulum transz-ciszterna cisz-ciszterna durva ER ciszterna
1/15. bra. Golgi-appartus szekrcis sejtben. A kpen egy diktioszmt ltunk, amely 7-8 prhuzamosan rendezett ciszternbl ll. A diktioszma konkv oldaln, a transz-ciszterna mellett srsd vladkot tartalmaz, n. kondenzl vakulumokat tallunk. Elektronmikroszkpos felvtel (28 700x, Rhlich P. felv.).
Forrs: http://www.doksi.hu
AZ L SZERVEZETEK ALAPVET KATEGRII: PRO- S EUKARITASEJTEK
49
lizoszmk
lizoszma
1/16. bra. Lizoszmk elektronmikroszkpos kpe (L. Kiss Anna felv.). A) A Golgi-appartus krnykn a mitokondriumnl tbbnyire kisebb, egyszer membrnnal hatrolt, stt tartalm kpletek formjban lthatk a lizoszmk (makrofg, 14 600x). B) Lizoszmk kimutatsa savas foszfatz hisztokmiai reakcival (a reakcitermk a lizoszmk belsejben stt csapadk formjban lthat, makrofg, 30 000x).
ronmikroszkpos kpen egyrtelmen azonostani, biztos felismersket az ket jellemz enzimek enzimcitokmiai vagy immuncitokmiai kimutatsa teszi lehetv. A lizoszmkat kitlt enzimek igen sokflk lehetnek, gyakorlatilag minden, a sejtet felpt szerves polimer molekult kpesek ptkveire bontani. gy vannak kztk nuklezok (DNz, RNz), protezok, glikozidzok, lipzok, foszfatzok, szulfatzok, lizozim stb. Ezek a hidrolitikus enzimek savi pH-n mkdnek optimlisan (savi hidrolzok), az ehhez szksges 5-5,5 pH-rtket
a lizoszma membrnjba ptett lizoszomlis protonpumpa (proton-ATP-z) biztostja, amely a lizoszma belsejbe H+-ionokat juttat be ATPbl szrmaz energia segtsgvel. A lizoszma bontenzimei a sejt szmra potencilis veszlyt jelentenek. A sejtet a lizoszmk rvn val nemsztdstl (autolzis) rszben a lizoszmt a citoszoltl elvlaszt lizoszmamembrn, rszben pedig a citoplazma kzel semleges pH-ja menti meg. A lizoszma enzimraktr, amely kszenltben ll, hogy a sejt szmra idegen szerves anyagokat,
Forrs: http://www.doksi.hu
50
BEVEZETS A SEJTBIOLGIBA
makromolekulkat lebontson. Ezek az anyagok (elhalt sejtek vagy azok maradvnyai, baktriumok, makromolekulk) tbbnyire kvlrl jutnak a citoplazmba azltal, hogy a sejtmembrnhoz tapadnak, majd a membrn e rszletnek a betremkedsvel s lefzdsvel felvevdnek a sejtbe (endocitzis). A felvett rszecskket membrn hatrolja el a citoplazma tbbi rsztl, mely az endocitzis sorn lefzdtt sejtmembrnnak felel meg. Ennek rvn kt membrnnal hatrolt kompartmentum ll rendelkezsre, az egyik a felvett s lebontand anyagot tartalmazza (fagoszma), a msik a lizoszma, mely a bont enzimek raktra. A kt kompartmentum membrnjnak a fzijval1 kommunikci jn ltre azok bels tartalma kztt, a kt kompartmentum egyetlen kompartmentumm olvad ssze, s a lizoszomlis enzimek megkezdhetik a felvett anyag lebontst. Ennek a mechanizmusnak az a nagy elnye, hogy a bont enzimek gy jutnak a lebontand anyaggal kapcsolatba, hogy egyetlen pillanatra sem rintkeznek a citoplazma sajt anyagval. Az emsztsi folyamat sorn a felvett anyag ptkveire bontdik, majd ezek a lizoszmamembrnon transzportldva bejutnak a citoszolba, s ott jra felhasznldnak. A lizoszmt abban az llapotban, mikor mg fagoszmval nem fuzionlt, primer lizoszmnak nevezzk, mg az sszeolvads utn keletkez kplet neve szekunder lizoszma vagy fagolizoszma. Br az idegen rszecske vagy anyag tbbnyire maradktalanul lebontdik a szekunder lizoszmban, egyes esetekben a lebonts nem tkletes. Klnsen hossz lettartam sejtekben fordul el, hogy a sejtben ilyen emszthetetlen anyagot tartalmaz maradvnytestek halmozdnak fel. Ezek sokszor lipideket is tartalmaz, srgsbarns szn granulumok formjban lthatk a fnymikroszkpos ksztmnyben (lipofuszcin granulumok, regedsi pigment). A lizoszma egyb, kvlrl felvett anyagok vgs llomst is jelenti. Az n. receptorfgg endocitzis rvn fehrjetermszet hormonok,
glikoproteinek, lipoproteinrszecskk is felvevdnek a sejtbe, majd az gy kialakult vezikulk egymssal sszeolvadva nagyobb vezikulkat (endoszmk) hoznak ltre, amelyek vgl szintn lizoszmval fuzionlnak. Vgl a lizoszmk arra is jk, hogy a sejt sajt citoplazmjnak egy rszt lebontsk, amelyre tbbnyire akkor kerl sor, ha a sejt kedveztlen krlmnyek kz kerl. Ehhez arra van szksg, hogy a lebontsra kerl citoplazmarszlet kr hatrol membrn alakuljon ki. Ezt a membrnnal krlvett rszletet (autofagoszma) a lizoszma mr idegenknt ismeri fel, s a tovbbiakban ugyangy kezeli, mint a kvlrl felvett anyagot tartalmaz fagoszmt, teht fuzionl vele s lebontja (autofgia). A fagocitzisra specializldott sejtek a lizoszmk segtsgvel a soksejt szervezetben fontos feladatokat ltnak el. Az intracellulris emszts rvn megtiszttjk a krnyezetet az elregedett sejtektl s az elhalt sejtek maradvnyaitl, fontos szerepk van az antibakterilis vdekezsben, rszben a baktriumok ellsvel, rszben a flig lebontott molekuladaraboknak a sejtfelsznre juttatsval s azoknak az immunsejtek szmra val bemutatsval. Ezen tlmenen bizonyos sejtekben a lizoszmk bont tevkenysgkkel biolgiailag aktv molekulkat (tiroxint a tireoglobulinbl, koleszterint az LDL-rszecskkbl) trnak fel.
1.3.1.2.7. Peroxiszma
A lizoszmkkal azonos mret vagy nmileg kisebb sejtorganellumok, amelyek morfolgiailag is hasonltanak a lizoszmkra (1/17. bra). Membrnnal hatrolt kpletek, szemcss szerkezet, nha kristlyt is tartalmaz beltartalommal. A hasonlsg abban is megmutatkozik, hogy a szemcss szerkezet enzimeknek felel meg, ezek azonban teljesen klnbznek a lizoszomlis enzimektl s tlnyomrszt oxidatv folyamatokat katalizlnak. Az egyik jellegzetes enzime a peroxidz, amelynek eredmnyeknt a szubsztrtmolekula oxidldik, s mellktermkknt hidrognperoxid keletkezik. A msik jellegzetes enzim a
Nem eldnttt, hogy kzvetlen sszeolvads trtnik, vagy kzvett vezikulk rvn kerl a fagoszma tartalma a lizoszmba.
Forrs: http://www.doksi.hu
AZ L SZERVEZETEK ALAPVET KATEGRII: PRO- S EUKARITASEJTEK
glikognszemcsk
51
mitokondrium peroxiszma
fehrjekristly
1/17. bra. Peroxiszmk mjsejtben. A peroxiszmt egyszer membrn hatrolja, belsejt finoman szemcss anyag tlti ki, amelybe itt stt fehrjekristly (urt-oxidz) van begyazva. A peroxiszmk felletes megtekintsre sszetveszthetk a mitokondriumokkal, a mitokondriumok azonban ltalban nagyobbak, ketts membrn hatrolja ket, s a mtrixtrben krisztk vehetk ki. Elektronmikroszkpos felvtel mjsejtjbl (17 500x, Rhlich P. felv.).
katalz, amely az ersen mrgez hidrogn-peroxidot tovbb bontja vzre s oxignre. Biztos azonostsuk is ezeknek az enzimeknek enzim- vagy immuncitokmiai kimutatsn alapszik. Jelentsgk mg ma sem ismert elgg. Rszt vesznek tbbek kztt zsrsavak oxidatv lebontsban, alkohol oxidcijban, hinyuk vgzetes lehet a szervezetre. Klnbz sejttpusokban klnbz gyakorisggal fordulnak el, leggyakrabban mj- s vesehmsejtekben tallhatjuk meg ket. A bennk jelenlv enzimek s az ltaluk katalizlt folyamatok alapjn egyesek gy vlik, hogy az si eukarita sejt mregtelent organellumrl lehet sz.
1.3.1.2.8. Mitokondrium
A sejt szmos energiaignyes feladatot (bioszintetikus, ozmotikus, mechanikai munka) lt el, amelyhez a szksges energit a krnyezetbl felvett tpanyagok tartalmazzk. Az energinak a tpanyagokbl a sejt szmra hasznlhat formban val val kinyerse bonyolult folyamat, ezrt a sejt arra rendezkedett be, hogy az gy kinyert energit egy knnyen felhasznlhat formban trolja. Az energiatrol molekula (a folyamatok tbbsgben) az adenozin-trifoszft (ATP), ahol az utols kt foszftcsoport ktse energiban gazdag. Az utols foszftcsoport levlasztsa (ATP-hasts) felhasznlhat energit szabadt fel, mg rktse (ATP-
szintzis) energit trol. Ezltal az ATP a legtbb energiaignyes munkhoz felhasznlhat akkumultorknt mkdik, melyet jra s jra fel kell tlteni energival. Ezt a feladatot a sejtben egy igen fontos organellum, a mitokondrium ltja el. A mitokondriumok tbbnyire plcika alak organellumok, melyek vastagsga nhny tized m, hossza akr tbb m is lehet (1/18. bra). Ez a mret a mitokondriumokat mr a fnymikroszkpban is lthatv teszi, bels, finomabb szerkezetket azonban csak az elektronmikroszkp tudta feltrni. A mitokondriumot kt, prhuzamosan elrendezd membrn s az ltaluk kzrezrt terek ptik fel. A kt membrn kzl a mitokondrium specifikus funkcijhoz az n. bels membrn az elengedhetetlen, amely lnyeges felletnagyobbtson is keresztlesik azltal, hogy lemezszeren tbb helyen betremkedik (mitokondrilis krisztk formjban) az ltala bezrt trbe. Az utbbit, az t kitlt stt, szemcss szerkezet anyag (mtrix) utn mtrixtrnek nevezzk, mg a kt membrn kztti lapos rst intermembranlis trnek hvjuk. Ettl az ltalnos smtl morfolgiai eltrsek elfordulhatnak, pl. egyes sejtekben a bels membrn nem lapos redk, hanem csvek formjban tremkedik be (n. tubulris szerkezet mitokondriumok), a mitokondrium alakja nem plcikaszer, hanem gmbly (pl. egyes mjsejtek esetben), a mitokondrium elgazdhat stb. Szmuk a nhnytl tbb ezerig terjedhet egyetlen
Forrs: http://www.doksi.hu
52
BEVEZETS A SEJTBIOLGIBA
tubulus
kriszta
mtrix-tr
A bels membrn kls membrn
1/18. bra. Kt klnbz tpus mitokondrium elektronmikroszkpos felvtele (Rhlich P. felv.). A) A bels mitokondriummembrn lemezszer kettzeteket bocst a mitokondrium belsejbe (kriszta tpus mitokondrium, 60 000x). B) A bels membrn finom csvecskk formjban nylik bele a mitokondrium belsejbe (tubulus tpus mitokondrium, mellkvese kregllomnybl, 33 000x)
sejtben, tbbnyire a sejt energiaignynek megfelelen Egyes sejttpusokban az energiafelhasznls helyhez kzel rendezdnek (pl. a spermium mitokondrilis hvelye, vesehmsejtek n. bazlis cskolata formjban). A mitokondriumban lezajl folyamatoknak az ADP-bl s foszftbl val ATP-szintzis csupn legvgs, br egyben legfontosabb mozzanata. Ezt azonban egy sor fontos elkszt lps elzi meg, amelyek sorn a tpanyagokbl az energia felszabadul, majd troldik. A citoszolban indul meg a glikogn s a zsr lebontsa cukrokk, ill. zsrsavakk. A cukorbonts (glikolzis) vgn hrom sznatomos molekula, a piruvt (piroszlsav) keletkezik, mely a zsrsavakkal egytt a kt mitokondrilis membrnon keresztl belp a mitokondrium belsejbe. A ktfle molekula oxidcis folyamatok rvn kt sznatomos ecetsavv bontdik tovbb, amely bekerl az
ugyancsak oxidcis folyamatokbl ll n. citrtkrbe. Az oxidcik rvn a sznatomok szndioxidd gnek el (mellktermk), ami azonban fontosabb, a sznatomokrl a hidrognatomokat hidrognszllt molekulk (NAD, FAD) viszik a bels mitokondriummembrnhoz. Ezutn kvetkezik az energia trolsa a mitokondriummembrnba ptett n. elektrontranszportlnc kzremkdsvel. A lnc egyms utn kapcsolt fehrjekomplexekbl ll, amelyek kzl az els a hidrognt protonra s elektronra bontja. A nagy energij elektront egyik komplex tovbbadja a msiknak, mikzben az elektron energijnak egy rszt arra hasznlja fel, hogy a membrnon protont pumpljon ki. A lnc vgn az elektron vgs akceptora az oxign, amely protonokkal egytt (mellktermkknt) vizet kpez. A protonpumpls kvetkeztben a membrn kls oldaln lnyegesen tbb proton gylik ssze, mint a bels oldalon,
Forrs: http://www.doksi.hu
AZ L SZERVEZETEK ALAPVET KATEGRII: PRO- S EUKARITASEJTEK
53
s ez a protongrdiens potencilis energiaraktrknt szerepel. A grdiens olyan vztoronyhoz hasonlthat, ahov energival vizet szivattyzunk fel, s ahonnan a vizet leeresztve ismt mozgsi energihoz jutunk (pl. egy turbint hajthatunk). Ugyangy a protonoknak a membrnon keresztl val ellenrztt visszaengedsvel ebbl az energiaraktrbl energia szabadthat fel, ami felhasznlhat az ATP-szintzishez. Ez az utols lps olyan, a membrnba ptett fehrjekomplexen keresztl megy vgbe, amelyen ioncsatorna vezet keresztl, s amely egyben adenozin-difoszftbl s foszftbl a felszabadult energia segtsgvel adenozin-trifoszftot szintetizl (ATP-szintetz). Az elektrontranszportlnc, az ltala ltrehozott protongrdiens s az ATP-szintzis egyarnt azt ignyli, hogy a bels mitokondriummembrn ersen impermebilis legyen, s rajta csak ellenrztt mdon jussanak ionok, molekulk keresztl. Mitokondrilis transzporterek felelsek azrt, hogy a piruvt, a zsrsavak, az ADP, a foszft bejussanak, mg a ksz ATP kijusson a mitokondriumbl. A bels membrnnal ellenttben a mitokondrium kls membrnja nem jelent komoly diffzis gtat, mert a kls membrnba csatornafehrjk (mitokondrilis porinok) vannak beptve, amelyek egy bizonyos molekulatmegig mindent tengednek. Ha meghzzuk a mitokondrilis folyamatok vgs egyenlegt, akkor azt mondhatjuk, hogy a mitokondrium szerves molekulkat hasznl fel, ezek oxidcijval (oxignigny!) szndioxidot s vizet kpez, a felszabadul energit pedig az ATP-be pti be. Pontosan az ellenkezje zajlik ezzel szemben a nvnyi kloroplasztiszban: a nvny a nap energijnak a felhasznlsval szndioxidbl s vzbl szerves anyagokat szintetizl, mikzben mellktermkknt oxign keletkezik. Meglep felfedezs volt, amikor a mitokondrium-mtrix terben DNS, tRNS s riboszmk jelenltt rtk le, ami arra utal, hogy ennek az organellumnak sajt genetikai informcija s gnexpresszis rendszere van. Kiderlt azonban, hogy ez csak a mitokondrilis proteinek mindssze kb. 5%-t lltja el, mg a maradk 95%-ot illetleg a mitokondrium a sejtmag DNS-nek s
gnexpresszis rendszernek a fggvnye. A citoplazmban szintetizlt fehrjk mitokondrilis lokalizcis jellel rendelkeznek, s ennek segtsgvel ismerik fel a mitokondriumot, majd jutnak t a kt membrnon vagy maradnak az egyik, ill. msik membrnban. A mitokondrium teht csak rszben fggetlen a sejttl, amelyre nagyrszt r van utalva, s ezrt nll letre kptelen (szemiautonm organellum). A sajt DNS s expresszis rendszer felvetette annak a gyanjt, hogy a mitokondrium az eukarita sejt evolcija sorn kvlrl felvett prokarita sejt volt. Valban, a mitokondrium molekulris sszetevi kztt szmos olyan vons van, amely a prokaritkra jellemz. Ezrt ma mr egyre elfogadottabb vlik az a nzet, hogy a mitokondrium valaha prokarita sejt lehetett, amelynek DNS-llomnya nagyrszt integrldott a gazdasejt nukleris DNS-be, s gy sejten belli szimbizis jtt ltre (endoszimbizis-elmlet).
1.3.1.2.9. Sejtvz (citoszkeleton)
Az eukarita sejtekben vkony fonlszer struktrkbl ll bels tmasztk van (1/19-21. bra), amelyet sejtvznak vagy citoszkeletonnak
mikrotubulusok
intermedier filamentuok
mikrofilamentumkteg
1/19. bra. A sejtvz (citoszkeleton) elemei. Elektronmikroszkpos felvtel fibroblasztsejtbl (23 200x, Rhlich P. felv.).
Forrs: http://www.doksi.hu
54
BEVEZETS A SEJTBIOLGIBA
sejtmag MTOC
mikrotubulusok
1/20. bra. A citocentrum. A kt centriolum finoman szemcss llomnyba (MTOC) van begyazva, amelybl mikrotubulusok sugrzanak ki minden irnyba. Elektronmikroszkpos felvtel fibroblasztsejtbl (19 900x, Rhlich P. felv.).
centriolum keresztmetszete
sejtmag
1/21. bra. Centriolum keresztmetszetben. Figyeljk meg a 9x3 elrendezdst. Elektronmikroszkpos felvtel thymus-limfocitbl (46 000x, Rhlich P. felv.).
neveznk. Ez a vz biztostja a sejt mechanikai ellenll-kpessgt, stabilizlja jellegzetes formj
sejtekben a sejt alakjt, de mdot ad arra is, hogy a sejt vltoztassa az alakjt, mozogjon, sszehzdjk, a sejten bell aktv mozgsok jjjenek ltre. A fehrjetermszet fonalak hrom csoportba oszthatk, ezek a mikrotubulusok, mikrofilamentumok s az intermedier filamentumok. A feloszts eredetileg elektronmikroszkpos megjelensk alapjn kszlt: a mikrotubulusok csszer struktrk 25 nm-es vastagsggal, a mikrofilamentumok vkony (5-7 nm vastagsg) fonalak, mg az intermedier filamentumok szintn fonlszer struktrk, amelyek vastagsga (10 nm) az elbbi kett kz esik, innen nyertk az intermedier nevet. Az eltr morfolgihoz eltr fehrjetpus s molekulris szerkezet is tartozik. A hromfle sejtvzkomponens tulajdonsgait sszehasonltva a legtbb hasonlsgot a mikrotubulusok s mikrofilamentumok kztt talljuk. Mindkett esetben a cs- vagy fonlszer elemek globulris fehrjk (monomerek) sszerendezdsbl jnnek ltre (polimerizci), az gy ltrejtt struktrk azonban jra szteshetnek ptelemeikre (depolimerizci), teht ezek a sejtvzkomponensek dinamikus struktrk. Mivel a polimerizci s depolimerizci lnyeges kvetkezmnyekkel jr, ezek pontos szablyozsa rendkvl fontos, ami egy sor jrulkos fehrjnek ksznhet (mikrotubulushoz, ill. mikrofilamentumhoz trsult fehrjk). Egy tovbbi fontos hasonlsg a ktfle elem kztt, hogy azokhoz motorfehrjk trsulhatnak: pl. a mikrofilamentumokhoz a klnbz miozinok, mg a mikrotubulusokhoz a dineinek s kinezinek. E fehrjk jellegzetessge az, hogy ATP-hastsbl szrmaz energia felhasznlsval alakvltozson esnek keresztl, s ezzel kt fix struktra kztt elmozdulst tesznek lehetv. Ennek az elvnek alapjn mozognak a sejtfelszn csilli, halad elre a spermium az ostor csapkod mozgsai rvn, hzdik ssze az izomsejt, jutnak el anyagok a sejt legtvolabbi rszbe az intracellulris transzport rvn, osztdik kett a citoplazma a sejtosztds vgn stb. Mind a mikrotubulris, mind a mikrofilamentris rendszer si kpzdmny, az ket felpt fehrjk keveset vltoztak az evolci folyamn (konzervatv fehrjk).
Forrs: http://www.doksi.hu
AZ L SZERVEZETEK ALAPVET KATEGRII: PRO- S EUKARITASEJTEK
55
Az intermedier filamentumok ezzel szemben filamentzus fehrjk ktegekbe rendezdsbl jnnek ltre, csak ritkn bontdnak le, motor fehrjkkel nem trsulnak, s ezzel a sejtvz tisztn mechanikai tmasztkot jelent stabil elemei. Az evolci sorn jelentsen vltoztak, st egyes sejttpusokra jellemz vltozatai is kialakultak.
1.3.1.2.9.1. Mikrotubulusok
A mikrotubulusok (25 nm tmrj, nem elgaz csszer struktrk) a tubulinfehrjbl plnek fel. A gobulris fehrje egymstl kiss eltr formban (- s -tubulin) fordul el, melyek egymssal heterodimert kpeznek, ez alkotja a mikrotubulus alapegysgt. A dimerek egyms vghez kapcsoldva hossz lncokat (protofilamentumokat) hoznak ltre, 13 db ilyen protofilamentum pedig egy hengerpalst mentn egymshoz prhuzamosan kapcsoldva hozza ltre a mikrotubulust. A mikrotubulusok labilis, az egyik vgkn knnyen leboml kpzdmnyek, stabilizlsukhoz kmiai mdostsok s specilis fehrjk (mikrotubulushoz asszocilt proteinek: MAPok) jrulnak hozz, le- s felplsket kalciums magnziumionok s GTP szablyozzk. A mikrotubulusok prhuzamosan rendezdve valdi vzszer struktrt hozhatnak ltre, amely stabilizlja a sejt alakjt. Ha mikrotubulusokat n. mikrotubulusmrgekkel lebontjuk, a sejt elveszti az eredeti alakjt, legmblydik. A mikrotubulusok egyik funkcija teht az alaktarts. Fontosabb ennl azonban, hogy a mikrotubulusok a citoplazmban olyan plykat kpeznek, amelyek mentn a motorproteinek anyagokat, vezikulkat, esetleg organellumokat mozgatnak. A motorfehrjk egyik vgkn a szlltand rakomnyhoz tapadnak, msik, mozgkony vgkkel pedig a mikrotubulus mentn vndorolnak, mindezt termszetesen ATP-hastsbl szrmaz energia segtsgvel. A mikrotubulusok tovbbi fontos szerepe, hogy komplex struktrkat, organellumokat hoznak ltre, ilyenek a centriolumok, a csillk s az ostorok, ill. az utbbiak n. bazlis testje. A cent-
riolum a sejt kzepn, a sejtmag mellett elhelyezked organellum, a citocentrum (sejtkzpont) kzpontjban elhelyezked hengerded testecske, amely 9 db, krben rendezd mikrotubulustripletbl ll (l. 1/20-21. bra). Tbbnyire kt centriolum tallhat a citocentrum kzepn egyms mellett (diploszma), mgpedig egymsra merleges llsban. A centriolum kialakulsa, sejtbiolgiai szerepe mg ma sem tisztzott. Az azonban biztos, hogy az t krlvev s -tubulint tartalmaz n. pericentriolris llomny a citoplazmatikus mikrotubulusok kiindulhelye (mikrotubulusorganizl centrum, MTOC), ahonnan egyre hosszabbra nve a sejt szli rsze fel sugrzanak szt. Klnsen feltn ez a jelensg a sejtosztdsban, amikor az osztdsi ors ltrejttekor a mikrotubulusok a sejt kt plusn elhelyezked citocentrumokbl sugrzanak ki. gy tnik, hogy a citocentrumnak a mikrotubulusorganizl funkcijhoz a centriolum nem okvetlenl szksges, mert egyes nvnyi sejtekben a centriolum hinyzik, mikrotubulusorganizl centrum mgis ltezik ehhez a pericentriolris anyag elegendnek ltszik. A mikrotubulusokbl felpl msik komplex struktra a csill (cilium, kinocilium (1/22. bra), valamint a hozz hasonl szerkezet, de hosszabb ostor (flagellum). Ezek egyes sejttpusok felsznrl kinyl, hengeres nylvnyok, amelyek jellegzetes csapkod vagy kgyz mozgst vgeznek. A mozgs htterben a mikrotubulusok s a hozzjuk trsul dinein motor molekulk llnak. Elektronmikroszkpban a csill (s az ostor) keresztmetszetn krben kilenc mikrotubuluspr keresztmetszete lthat, mg kzpen szintn kt mikrotubulus helyezkedik el (9x2+2 mintzat). A mikrotubulusprok kztti rsben dineinkomplexek (dineinkarok) tallhatk, ezek talpa az egyik mikrotubulushoz tapad, mg mozgkony feji rsze a szomszdos mikrotubulust mozdtja el hosszanti irnyban. A kt szomszdos mikrotubuluspr gy elcsszik egyms mellett, ami a csill meggrblshez vezet. Ehhez a mechanikai munkhoz termszetesen ATP-hastsbl szrmaz energia is szksges. A csillt kvlrl sejtmembrn bortja be, tvnl pedig a centriolum-
Forrs: http://www.doksi.hu
56
BEVEZETS A SEJTBIOLGIBA 1/22. bra. Csillk keresztmetszeti kpe. Egy-egy csilln bell jl ltszik a kzps kt mikrotubulus, a sugaras kllk, a 9 szli mikrotubuluspr, egyik oldalukon a dineinkarokkal. A csillt a sejtmembrn bortja be. Elektronmikroszkpos felvtel (122 000x, Rhlich P. felv.).
sejtmembrn kzps kt mikrotubulus
dineinkar
perifris mikrotubuluspr
kll
mal azonos szerkezet testecske, az n. bazlis test tallhat. A csill s az ostor ltal keltett mozgs ktflekppen hasznosul, aszerint, hogy a sejt szabadon mozog vagy rgztett. Szabad sejt esetben a csill az elremozgst szolglja (pl. csills s ostoros egysejtek, spermiumok), mg egy hmrtegbe ptett csills sejt a hmrteg felsznt bort folyadk ramlst biztostja (a lgcs s a petevezetk csills hmja).
1.3.1.2.9.2. Mikrofilamentumok
Az 5-7 nm vastag mikrofilamentumok globulris fehrjbl, az aktinmolekulbl (G-aktin) plnek fel gy, hogy azok egyms vghez kapcsoldva hossz lncot alkotnak (F-aktin). Kt ilyen lnc egyms krl csavarodva alkot egy mikrofilamentumot. Aktin-mikrofilamentumok jelents mennyisgben vannak a citoplazmban, s gyakran alkotnak ktegeket vagy sr szvedket. Az utbbit szinte rendszeresen megtallhatjuk a sejtek szli, sejtmembrn alatti znjban (sejtkortex), ahol a sejtmembrnnak ad tmasztkot, mechanikai ellenllkpessget. Az aktinmikrofilamentumok szvedke a citoplazmt viszkzuss teszi (a kolloidikai gl-lla-
pothoz hasonl llapotv), mg a filamentumok gyors lebomlsa, depolimerizcija a citoplazma folykonyabb vlst idzi el (szol-llapot). gy a citoplazma konzisztencija nagymrtkben fgg az aktin polimerizcijtl vagy depolimerizcijtl. A mikrofilamentumok felplsben s lebomlsban, a mikrofilamentum-szvedkek, -ktegek stabilizcijban, a membrnhoz ktsben egy sor fehrje jtszik szerepet (aktinhoz trsult fehrjk). Tekintve hogy a szolgl talakuls adott jelekre helyileg is ltre tud jnni a citoplazmban, miltal egyes terletek viszkzusabb, msok folysabb vlnak, a sejt alakjt tudja vltoztatni, s a citoplazmban ramlsok jhetnek ltre. Egy felsznhez letapadt sejt helyvltoztatsban, ksz, vndorl mozgsban (amboid mozgs) ez a loklis szolgl talakuls lnyeges szerepet jtszhat. A mikrofilamentumoknak e dinamikus sajtsgai mellett fontos formakonzervl szerepk is van. Tipikus pldi ennek a sok sejt felsznrl kill, plcikaszer nylvnyok, az n. mikrobolyhok (1/23. bra). Egy-egy ilyen mikroboholy vzt 30-40 mikrofilamentumbl ll kteg kpezi, amelyben a filamentumokat keresztkt fehrjk rgztik egymshoz s a mikroboholy felsznt kpez sejtmembrnhoz. A prhuzamosan ll, me-
Forrs: http://www.doksi.hu
AZ L SZERVEZETEK ALAPVET KATEGRII: PRO- S EUKARITASEJTEK
57
1/23. bra. Mikrobolyhok keresztmetszetben. A mikrobolyhokat sejtmembrn bortja, belsejkben 8-10 mikrofilamentum pontszer keresztmetszeteit ltjuk. Elektronmikroszkpos felvtel vkonyblhmsejt kefeszeglybl (57 500x, Rhlich P. felv.).
rev mikrobolyhok rendszere a sejtmembrn felsznt jelentkenyen (10-20-szorosra) megnvelheti, ami olyan sejttpusokban jelents, ahol a membrnba fontos enzimek vagy transzportfehrjk vannak beptve (a vkonybl hmja, proximlis vesetubulus hmja). A sejt szabad felsznt ilyenkor a mikrobolyhok szzai-ezrei borthatjk igen szablyos rendszert alkotva, ahol a mikrobolyhok a kefe szrszlaihoz hasonlan srn egyms mellett helyezkednek el (kefeszegly). A mikroboholyhoz hasonlan plnek fel, de annl nagyobbak az n. sztereociliumok (nem mozgkony nylvnyok!), ezeket fknt egyes rzksejtek szabad felsznn talljuk meg (a hall- s egyenslyoz-szerv szrsejtjei, zlelbimbk rzksejtjei). Az aktinmikrofilamentumoknak azonban nemcsak formatart, vzszer funkcijuk van, hanem a filamentumok a sejtek dinamikus, aktv mozgsfolyamataiban is igen jelents szerepet jtszanak. Az aktinhoz ugyanis mozgat (motor-) fehrje, a miozin ktdhet, ami energiafgg alakvltozsval elmozdulsokat tesz lehetv. A miozinnak klnbz hosszsg farki s mozgkony feji rsze van, mely utbbi az aktin-mikrofilamentumhoz tud ktdni. A feji rsz konformcivltozssal begrbl, s a mikrofilamentumot
ezzel kiss elcssztatja. Majd ATP ktsvel a feji rsz levlik az aktinrl, ATP-hastsra kiss kiegyenesedik, s jra a mikrofilamentumhoz ktdik, miltal a ciklus jrakezddik. A miozin s aktin egymshoz viszonytott elmozdulst a sejtek ktflekpen hasznostjk. Az egyik lehetsg az, mikor a mikrofilamentum rgztett s a miozin a mozgkony: ilyenkor ATP jelenltben a miozin vndorol az aktinmikrofilamentum, mint plya mentn. Amennyiben a miozin ms, mozgathat kplet (pl. vezikulk) felsznhez ktdik, a miozin (V. tip. miozin) ugyangy rszt vehet ezek intracellulris szlltsban, mint ahogy azt a mikrotubulusok s a hozzjuk tartoz motor molekulk (dinein, kinezin) esetben lttuk. A msik lehetsg, amikor a miozin-molekulk (II. tip. miozin) hossz, farki rszk rvn asszocildnak, s hosszan megnylt, virgcsokorszer ktegeket alkotnak. Kt ilyen kteg szimmetrikus sszekapcsoldsval jnnek ltre az izomsejt n. vastag (miozin-) filamentumai. Az utbbiak, az aktinmikrofilamentumok kztt elhelyezkedve (a miozinfejek begrblsvel) az aktinmikrofilamentum-rendszert a vastag filamentumokhoz kpest elcssztatjk, s gy a kt filamentumflbl ll egyttes kteg megrvidl (kontrakci). Az gy ltrejv kontrakcis rendszer a legnagyobb fejlettsget a harntcskolt izomban ri el, ott is fedeztk fel, majd tanulmnyoztk a legrszletesebben, azonban nem izomsejtekben is elfordul, termszetesen sokkal egyszerbb formban. Ennek egyik tipikus pldja az llati sejtek osztdsnak utols fzisban kialakul n. kontrakcis gyr. Ez az aktinmikrofilamentumokbl s miozinbl ll kteg krkrs gyrt kpez a sejtmembrn alatt, majd fokozatosan rvidlve kettvlasztja a citoplazmt.
1.3.1.2.9.3. Intermedier filamentumok
A 10 nm vastag intermedier filamentumok (1/24. bra) fibrzus fehrjemolekulk egyms mell s egyms vgeihez val illeszkedsbl jnnek ltre. Kifejezetten mechanikai szerepk van, sejtek alakjt stabilizljk, vagy a sejteknek
Forrs: http://www.doksi.hu
58
BEVEZETS A SEJTBIOLGIBA
1.3.1.3. A nvnyi sejt specifikumai
Az elbbiekben az eukarita sejt jellegzetessgeit egy llati sejt pldjn mutattuk be. A nvnyi sejt felptse alapjaiban hasonl az llati sejthez, mgis nhny jellegzetes klnbsget mutat (1/25. bra). A legfontosabb klnbsg a sejt energiaforgalmban rejlik, mert a nvny a nap energijt hasznostja, s pt fel ezzel szndioxidbl s vzbl szerves anyagokat (fknt sznhidrtokat) egy specilis organellumban, a kloroplasztiszban. Ugyanakkor rendelkezik az llati sejt lehetsgeivel is, mert szerves anyagokat lebontva a mitokondriumokban ATP-t tud szintetizlni.
sejtmag intermedier filamentumok mitokondrium
1/24. bra. Intermedier filamentumokbl ll kteg. Elektronmikroszkpos felvtel makrofg sejtbl (18 800x, Rhlich P. felv.).
nagy mechanikai ellenllkpessget adnak. Klnbz sejttpusokban ugyanannak a molekulacsaldnak klnbz vltozatait talljuk meg. gy pl. a mechanikai hatsoknak leginkbb kitett brhm sejtjei nagy mennyisgben tartalmazzk az itt keratinfehrjkbl ll intermedier filamentumokat, az idegsejtekben a neurofilamentumfehrjbl ll intermedier filamentumok biztostjk a sejt nylvnyos alakjnak a megrzst, az idegrendszer tmasztsejtjeiknt ismert gliasejtekben ugyanezt a feladatot egy msik fehrjbl (GFAP) felpl intermedier filamentumrendszer ltja el. Az intermedier filamentumok egy fajtja azonban minden sejtflesgben megtallhat, mgpedig a sejtmagot krlvev maghrtynak a mag belseje fel es oldaln. Itt a laminfehrjkbl felpl intermedier filamentumok egy vkony rostos rteget (nukleris lamina) alkotnak, amely a maghrtya rugalmas mechanikai ellenllkpessgt biztostja.
A kloroplasztisz a mitokondriumhoz bizonyos mrtkig hasonl szerkezet, de annl jval nagyobb sejtorganellum, melynek f feladata szerves molekulk szintzise a fny energijnak felhasznlsval (fotoszintzis). Hasonlan a mitokondriumhoz, a kloroplasztiszt is kt membrn hatrolja. Ezek kzl a kls membrn knnyen tjrhat szmos anyag szmra, a bels membrn ezzel szemben gtat kpez a szabad diffzival szemben, s rajta csak ellenrztten (transzportmolekulk segtsgvel) juthatnak t kivlasztott anyagok. A bels membrn a mitokondrium mtrixhoz hasonlan egy sztrmnak nevezett anyagot hatrol krbe, ez klnbz enzimeket, a plasztisz sajt gnexpresszis rendszernek egyes elemeit (DNS, tRNS, riboszmk), valamint a plasztisz ltal szintetizlt anyagokat, elssorban kemnytt tartalmaz. Ezenkvl a sztrmba begyazva lelapult zskok, n. tilakoidok tallhatk, melyek thzdhatnak az egsz sztrmn, helyenknt pedig korongokat alkotva pnztekercsszeren egymsra rendezdnek (n. granumok, tsz.: grana). A tilakoidok bels tere (tilakoidtr) nem kzlekedik a kloroplasztisz kls s bels membrnja kztti trrel (intermembranlis tr), viszont az egyes tilakoidok bels tere egymssal igen. A tilakoidmembrn igen fontos a kloroplasztisz mkdsben, mert ez tartalmazza a fotoszintzist lebonyolt molekulris mechanizmus szmos elemt.
Forrs: http://www.doksi.hu
AZ L SZERVEZETEK ALAPVET KATEGRII: PRO- S EUKARITASEJTEK
59
1/25. bra. Magasabb rend nvnyi sejt, vzlat.
A kloroplasztiszban lezajl folyamatokat gy sszegezhetjk, hogy fnyenergia felhasznlsval vzbl s szn-dioxidbl sznhidrtok szintetizldnak, ennek sorn mellktermkknt oxign keletkezik. Ez a folyamat sok tekintetben a mitokondriumban lezajl trtnsek fordtottjaknt foghat fel. Lnyegben kt rszre bonthat. Az els rsz hasznostja a fny energijt vz felhasznlsval, ennek sorn a sznhidrtok ksbbi (a sttszakaszban lezajl) szintzishez szksges ATP (energia) s NADPH (redukl er) termeldik. Ezen n. fnyszakaszhoz teht fnyenergia szksges, amelynek hasznostst kt, klorofillt s fehrjket tartalmaz komplex (fotoszisztma I s II) vgzi. A foton ltal a fotoszisztma II klorofilljbl kittt nagy energij elektron egy (a tilakoidmembrnba ptett) elektrontranszport-lncon vezetdik keresztl, s
protongrdiens ltrehozsval ugyanolyan mechanizmuson t vezet ATP-szintzishez, mint a mitokondriumban. A klorofillbl kittt elektron helyt a vzmolekula bontsbl szrmaz elektron foglalja el (2H2O 4e + 4 H+ + O2). Az elektrontranszport-lnc vgn a cskkent energij elektron bevezetdik a fotoszisztma I be, ahol egy msik foton abszorpcijval a klorofillbl kittt nagy energij elektron helyre esik be. A nagy energiaszintre emelt elektron energijt egy enzim, a NADP-reduktz hasznlja fel arra, hogy proton segtsgval H keletkezzk s az a H-transzporter NADP-molekulra ktdjk (NADPH). A fnyszakasz vgtermke teht ATP, NADPH s oxign. A fotoszintzis msodik rszhez mr nem kell fny (sttszakasz). Ez a szndioxid-asszimilci folyamata, melynek sorn szndioxidbl s vzbl,
Forrs: http://www.doksi.hu
60
BEVEZETS A SEJTBIOLGIBA
a fnyszakasz folyamn termeldtt energia (ATP) s redukl er (NADPH) felhasznlsval 3 C-atomos sznhidrt-molekula, a glicerinaldehid-3-foszft jn ltre. A szndioxid ciklusosan egy anyagcserekr mentn (Calvin-kr) asszimilldik, ahol kiindulsknt egy 5 C-atomos molekula, a ribulz-1,5-biszfoszft szerepel. A glicerinaldehid-3-foszft a nvnyi sznhidrt-anyagcsere kzponti molekulja, amelybl mg a kloroplasztiszon bell, a sztrmban klnbz cukrok keletkeznek. A keletkez glukz kemnytv polimerizldik, s akkora tmeget rhet el, hogy a sztromban mint kemnytszemcse fny- s elektronmikroszkposan is lthatv vlik. A nvnyi sejt szvesen hasznlja transzportcukorknt a glukzbl s fruktzbl ll diszacharidot, a szacharzt is, s exportlja fotoszintzissel nem rendelkez sejteknek. A glicerinaldehid-3-foszft visszacsatoldhat mg a Calvin-krbe vagy talakulhat piruvtt, amikor is a mitokondriumban az ATP-szintzist tpllja. Itt jegyzend meg, hogy a nvnyi sejt az energiaignyes folyamatokhoz nem a fnyszakasz sorn termeldtt ATP-t hasznlja fel, hanem az ATP-t az llati sejthez hasonlan a mitokondrilis ATP-szintzisbl nyeri (amihez termszetesen oxignt hasznl fel). Vgl a fotoszintzis rvn termelt sznhidrtot a sejt kiindulpontknt hasznlja egyb molekulk (zsrsavak, aminosavak stb.) szintzishez is. A kloroplasztisz a fenti szerkezeti s bizonyos mrtk funkcionlis hasonlsgok mellett abban is hasonlt a mitokondriumhoz, hogy sajt gnexpresszis rendszere van. A sztrmban tallhat gyr alak DNS a kloroplasztisz sajt fehrjinek kb. 30%-t kdolja, ezek az ugyancsak itt tallhat tRNS-ek s riboszmk segtsgvel a kloroplasztisz belsejben szintetizldnak. Az sszes tbbi fehrjt a sejtmagban lv DNS kdolja, s ezek a citoszolban szintetizlt fehrjk a rajtuk lv lokalizcis jel segtsgvel msodlagosan jutnak be a kloroplasztiszba. A mitokondriumhoz hasonlan a kloroplasztisz esetben is felttelezhet, hogy az evolci sorn a plasztisz a sejtbe felvett s azzal szimbizisba lpett prokarita sejtbl szrmazik, ami ebben az esetben valsznleg egy fotoszintetizl cianobaktrium volt.
A kloroplasztiszon kvl a nvnyi sejt egyb jellegzetessgeket is mutat. Az egyik ezek kzl a sejtet krlvev merev sejtfal. Anyagt maga a sejt termeli s vlasztja ki maga kr. A sejtfal nagy rszt cellulzrostok kpezik, melyek egy poliszacharidbl s fehrjkbl ll alapanyagba vannak begyazva. A merev falbl ll kamra biztostja a sejt alakjt, vdi a sejtet a mechanikai hatsoktl s az ozmotikus duzzadstl. Ez a kamra volt az, amit Robert Hooke a XVII. sz.-ban egyszer mikroszkpjval felfedezett, ennek alapjn nevezte ezt el cellnak, amelyrl a sejt ksbb a nevt kapta. A nvnyi sejtet jellemzi mg a sejt kzepn tallhat, membrnnal hatrolt reg, az n. centrlis vakulum. A benne uralkod savi pH s a lizoszomlis enzimek jelenlte miatt a nvnyi sejt lizoszmjnak is tekinthetjk, egyben azonban a nvnyi sejt lerakhelye is, ahol sk, az anyagcsere mellktermkei s egyb anyagok troldnak. A nvnyi sejt peroxiszmi klnleges feladatokat lthatnak el, ilyen a kloroplasztiszban lezajl sznasszimilci egyik mellktermknek a feldolgozsa vagy a csrzs alatt a mag zsrtartalkainak talaktsa cukorr. Nvnyi sejtek, az ket krlvev sejtfal miatt nem rintkezhetnek kzvetlenl egymssal, mint az az llati sejteknl gyakran megszokott. Magasabb rend nvnyekben a kapcsolatot a szomszdos sejtek kztt a sejtfalon keresztlfrd citoplazmahidak (plazmodezmk) tartjk fenn, melyek rvn a kt sejt citoplazmja folytonos kapcsolatban ll egymssal.
1.3.1.4. Az egysejt eukaritk specifikumai
Olyan eukarita sejtek, amelyek nem szervezdnek soksejtes szervezetekbe, hanem nll letre kpesek. letben maradsukhoz klnbz stratgikat hasznlnak, ami a sejt felptsben is sokszor tkrzdik. Itt csupn kt kpviseljket emltjk meg. Az lesztsejt az egyik legegyszerbb eukarita sejt, amelyet egysejt gombnak tekinthetnk.
Forrs: http://www.doksi.hu
AZ L SZERVEZETEK ALAPVET KATEGRII: PRO- S EUKARITASEJTEK
61
Az ovlis sejt nagysga a 6 m-t nem haladja meg, alakjt az arnylag merev sejtfal biztostja. A sejt citoplazmja tartalmazza mindazokat az organellumokat, amelyek az eukarita sejt mkdshez szksgesek, gy az endoplazmatikus retikulumot, a Golgi-appartust, a lizoszmt, a mitokondriumot, a riboszmkat stb. Sznhidrtban gazdag krnyezetben gyorsan szaporodnak (hasadssal vagy bimbzssal), szmuk nhny ra alatt megkettzdik. Arnylag egyszer felptsk, gyors szaporthatsguk kivl ksrleti modell teszik ket az alapvet sejtbiolgiai mechanizmusok feldertsre. Meglep, hogy az leszt funkcikiesses mutnsai segtsgvel felfedezett fehrjk kzl milyen soknak felelnek meg pl. emlsllatokban hasonl szerkezet s funkcij fehrjk. Amennyiben egy fontos lesztfehrje gnje ismertt vlik, arnylag knny feladat molekulris biolgiai mdszerekkel az emls- (akr emberi) sejtben a megfelel homolg gnt, majd gntermket megtallni. Az itt megemltend msik egysejt az amba, amely az lesztsejttel ellenttben igen nagy sejt, hossza ellapult llapotban az egy millimtert is meghaladhatja, s trfogata az lesztsejtnek akr tezerszerese is lehet. Maga a sejtmag is risi, tbb szz kromoszmval s tbb tucat magvacskval. Az amba nagysga j alkalmat nyjtott a sejtbiolgusok szmra, hogy mikrosebszi beavatkozsokkal (pl. a sejtmag eltvoltsa) egyes sejttani mechanizmusokat tanulmnyozzanak. A sejt tpllkt l baktriumok, algk s egyb egysejtek fagocitzisval nyeri, ezrt benne a fagocitzis s az intracellulris emszts klnsen fejlett. Ennek rdekben a sejtmembrn kls felsznt egy tapads, sznhidrtokban gazdag rteg bortja, a citoplazmban pedig nagyszm lizoszma vrakozik arra, hogy a bekebelezett tpllkot tartalmaz n. tpllkvakulummal fzionljon, s ezzel azt lebontsa. Az amba egy msik, a fagocitzissal rokon tulajdonsga, hogy szilrd felsznhez letapadva azon vndorolni kpes. Sejtnylvnyok (llbak, grgl pszeudopdiumok) kibocstsval vndorol, amibe a citoplazma tbbi rsze fokozatosan tmlik, s ezzel a sejt a helyt vltoztatja. Ilyen sejtvndorls ma-
gasabb rend llati szervezetekben is fontos szerepet jtszik (pl. makrofgok, fehrvrsejtek esetben), a mozgsnak ezt a tpust az ambrl amboid mozgsnak neveztk el. Az desvizi amba a citoplazmba jutott felesleges vztl n. kontraktilis vakulum segtsgvel szabadul meg. A vakulum vizet tartalmaz vezikulk sszeolvadsbl alakul ki, majd a vakulum kell nagysgot elrve fzionl a sejthrtyval, s sszehzdva kirti tartalmt a krnyezetbe.
1.3.2. Az let peremn
A legkisebb s egyben legegyszerbb prokarita sejtek a mikoplazmk. tmrjk nem tbb, mint 0,1 m, teht elri a nagy vrusok nagysgt, s gy a legtbb baktrium-szrn is kpesek tjutni. Burokkal nem rendelkeznek, DNS-k kb. 700 klnbz fehrjt kdol, s ez mg ppen elegend, hogy szmukra nll ltezst biztostson. Citoplazmjukban a riboszmk szmt 400-ra becslik. Ezek az let als hatrn l sejtek a legkisebb ltez autonm l rendszerek, s gy a legprimitvebb sejteknek (minimum sejt) tekinthetk. Talajban l szaprofitk (azaz nem l szervezetekbl szerzik tpllkukat), de krokozknt tdgyulladst, nylmirigy-gyulladst is okozhatnak. A vrusok s fgok valamilyen nukleinsavat (DNS vagy RNS) tartalmaz s tbbnyire fehrjeburokkal rendelkez rszecskk, melyekhez egyes esetekben membrnburok is trsulhat. nll letre kptelenek, mivel sem a szaporodsukhoz, sem az anyagcserhez szksges appartussal nem rendelkeznek, nmagukban letjelensget nem mutatnak. Szaporodni mgis kpesek, mgpedig gy, hogy valdi sejtekbe (fgok esetben prokaritkba, virusok esetn eukarita sejtekbe) hatolnak be, s az ilyen n. gazdasejt nukleinsavs fehrje-szintetizl rendszert hasznljk fel nmaguk jratermelsre. A virusok s fgok teht nem valdi sejtek, hanem csak parazita-jelleg, szemi-autonm makromolekulris komplexek, melyek egyes elkpzelsek szerint valdi sejtekbl kiszakadva jttek ltre az evolci sorn.
Forrs: http://www.doksi.hu
2.
A sejt legfontosabb anyagi sszetevi s alapvet molekulris mechanizmusai. A sejtbiolgia molekulris biolgiai eszkztra.
SZAB GBOR S NAGY LSZL
A sejt legfontosabb anyagi sszetevi s alapvet molekulris mechanizmusai A fehrjk A sejtek energiaforgalmnak kulcsszereplje: az ATP A DNS szerkezete DNS-replikci DNS-repair DNS mRNS (transzkripci), mRNS fehrje (transzlci) 2.1.7. 2.1.8. 2.1.9. 2.1.10. 2.1.11. 2.2. 2.2.1. 2.2.2. A naszcens RNS mRNS-s rse A fehrjk poszttranszlcis mdostsai s clba juttatsa A gnexpresszi szablyozsa ltalnos szablyozsi alapelvek A DNS heterogenitsa A sejtbiolgia molekulris biolgiai eszkztra Mdszerek Modellllnyek
2.1. 2.1.1. 2.1.2. 2.1.3. 2.1.4. 2.1.5. 2.1.6.
2.1. A sejt legfontosabb anyagi sszetevi s alapvet molekulris mechanizmusai
2.1.1. A fehrjk
Egy baktriumsejt f makromolekulris sszetevit s azok szintzishez szksges kismoleku-
lkat mutatja a 2/1A bra. A sejtek letmkdseiben a legalapvetbb szerepet a fehrjk tltik be, mint strukturlis feladatokat ellt molekulk (struktrfehrjk) vagy mint folyamatok katalizlst vgz enzimek. Trszerkezetk kialakulsa (2/1B bra) elssorban az aminosavszekvencia ltal meghatrozott (elsdleges szerkezet). Az egydimenzis aminosavlnc, szekvencijnak megfelelen kialakul hidrognhidak rvn vagy heliklis (n. alfa-heliklis), vagy n. redztt (merev, lemezszer elemekbl hajtogatott, pleatedsheat) msodlagos szerkezeti szintet alkot. Az alfa-
Forrs: http://www.doksi.hu
64 A SEJT LEGFONTOSABB ANYAGI SSZETEVI S ALAPVET MOLEKULRIS MECHANIZMUSAI. A SEJTBIOLGIA MOLEKULRIS BIOLGIAI ESZKZTRA.
A
baktriumsejt ionok s kismolekulk (4%) foszfolipidek (2%)
30% szerves s szervelen vegyletek
a sejtet felpt anyagok cukrok zsrsavak aminosavak nukleotidok
a sejtet alkot nagyobb egysgek poliszaharidok zsrok/lipidek/membrnok fehrjk nukleinsavak
DNS (1%)
RNS (6%)
makromolekulk
70% H2 O fehrjk (15%) adenozin trifoszft (ATP) O O P O poliszaharidok (2%) O O P O O O P O H H O N HC
NH2 C C C N CH2 O H H OH OH 1 N CH N
aminosavak
az amjnosavakbl felpl polipeptidlnc a fehrje elsdleges szerkezete
antignek
-lemez
-hlix
a fehrjk msodlagos szerkezett az aminosavak kztt kialakul H-ktsek hozzk ltre
antign antignkt fragment
-lemez
a fehrjk harmadlagos szerkezett az -hlixek s -lemezek kztt ltrejv kmiai hatsok alaktjk ki
-hlix
a fehrjk negyedleges szerkezett tbb polipeptidlnc alaktja ki
antitest
2/1. bra. A sejt legfontosabb anyagi sszetevi (A). A fehrjk szerkezete (B, C).
Forrs: http://www.doksi.hu
A SEJT LEGFONTOSABB ANYAGI SSZETEVI S ALAPVET MOLEKULRIS MECHANIZMUSAI
65
heliklis vagy redztt msodlagos konformcit felvett elemek egytt hozzk ltre a fehrje n. harmadlagos struktrjt. Tbb ilyen alegysg (fggetlen polipeptidlnc) kapcsoldsa rvn negyedleges szerkezet jhet ltre, melynek egy pldja az idegen anyagokat (antigneket) felismer antitest (immunoglobulin, 2/1C bra). Az utbbi knny s nehz lnca vgei ltal kpzett felismer hely rendkvl sokfle lehet, az antitest kpzdst kivlt testidegen molekulknak (antigneknek) megfelelen. Az enzimatikus mkds fehrjk az ltaluk talaktand anyagcseretermkeket (szubsztrtokat) aktv centrumukban megktik, s a reakcit azltal katalizljk, hogy azt j tra terelve annak vgbementelhez szksges aktivcis energiahegyet lecskkentik. Ezltal mr a krnyezet termikus energija is elg ahhoz, hogy a reaktnsok szmra energetikailag kedvez, mlyebb energiaszintet eredmnyez reakci vgbe is menjen. Egy enzimnek egy msik fehrje is lehet szubsztrtja, pl. foszforillhat egy msik fehrjt, mint szubsztrtjt. Az enzimmkdst befolysolhatjk kisebb szablyoz anyagok is, melyek a fehrje egy msik rszn, az n. alloszterikus kthelyen megktdve modulljk az enzim konformcis llapott.
szksgk glukz felvtelre a klvilgbl, hiszen fotoszintzis rvn maguk szintetizljk a nap fnyenergijt felhasznlva, CO2-bl s vzbl (autotrfok).
2.1.3. A DNS szerkezete
A sejtmagba csomagolt rktanyag, a DNS cukor-foszft vzbl s ahhoz kapcsold bzisokbl ll dupla-heliklis struktra, melynek kt fele az egyms fel nz bzisok kztti H-ktsek ltal kapcsoldik egymshoz (2/2A bra). Ngyfle bzis van a DNS-ben, adenin (A), guanin (G), timin (T) s citozin (C). A DNS ltal preferencilisan felvett konformcis llapotban a lehetsges prosodsok: A-T (kt H-hddal) s G-C (3 H-hddal). A DNS bizonyos krlmnyek kztt ettl az (n. B-) konfigurcitl eltr mdon is tekeredhet (ez az n. Z-DNS), st tbbszl konformcik, hajtszer kpzdmnyek keletkezse s klnbz DNS-molekulk kztti bonyolult, sszefgg struktrt eredmnyez kapcsoldsok is lehetsgesek ezen alternatv llapotok fiziolgis (lben betlttt) szerepe mg nem tisztzott. Az eukarita genomon (kromoszmkon) kvl a mitokondriumokban s a kloroplasztiszokban is van (kr alak) DNS, ill. n. episzm alakban, nllan replikld, extrakromoszomlis elemekknt fordulhat el, pl. daganatos sejtekben vagy virlis fertzs kapcsn. A baktriumok egyetlen kromoszmja szintn kr alak DNS, mely egy ponton a membrnhoz rgzl. A baktriumok belsejben is tallhatk extrakromoszomlis DNS-elemek, pl. a gygyszerrezisztenciagneket hordoz plazmidok.
2.1.2. A sejtek energiaforgalmnak kulcsszereplje: az ATP
Az llati (heterotrf) sejtek letmkdseik energiaignyt tpllkozsuk sorn felvett szerves anyagok, elssorban glukz elgetsbl nyerik. Ez a folyamat sok, enzimek katalizlta lpsben megy vgbe, mely lpsek sorn adenozin-trifoszft (ATP) keletkezik (l. 2/1A bra). Az ATP vglls foszftcsoportjnak lehasadsa az anyagcsere-folyamatok energiaignyvel sszemrhet energit mobilizl, ezrt az ATP-t a sejt univerzlis energiahordozknt hasznlja. A cukor elgetse nvnyi sejtekben is hasonl mdon s cllal megy vgbe, a nvnyeknek azonban nincs
2.1.4. DNS-replikci
A sejt osztdsa sorn a DNS gy replikldik (2/2B bra), hogy a szttekered lncokhoz azokkal komplementer j DNS-szl szintetizldik. Ezt a mechanizmust szemikonzervatv replikcinak nevezzk, mert az j dupla-hlix egyik szla korbbi
Forrs: http://www.doksi.hu
66 A SEJT LEGFONTOSABB ANYAGI SSZETEVI S ALAPVET MOLEKULRIS MECHANIZMUSAI. A SEJTBIOLGIA MOLEKULRIS BIOLGIAI ESZKZTRA.
A
DNS molekula
foszftcsoport
cukor
adenin vagy guanin bzisok
citozin vagy timin
B
5
j szl DNS-polimerz
3 vezet templt helikz ksleked templt
egyszl DNS-hez ktd fehrjk
szli DNS 5
5 DNS-polimerz Okasaki fragment RNS primer DNS-primz
2/2. bra. A DNS szerkezete (A) s replikcija (B).
Forrs: http://www.doksi.hu
A SEJT LEGFONTOSABB ANYAGI SSZETEVI S ALAPVET MOLEKULRIS MECHANIZMUSAI
67
DNS-molekulbl szrmazik (regebb, mint a msik szl). A DNS kt szla rszben eltr biokmiai mechanizmussal szintetizldik. Ennek az az oka, hogy a DNS polimerizcijt vgrehajt DNS-polimerz enzim a cukor-foszft vz ribznak ts (n. 5) sznatomjhoz kapcsold foszfthoz tudja csak a kvetkez bzis-cukor-foszft alegysget csatlakoztatni, teht az j lnc 5 > 3 irnyban tud csak nni. Az n. vezet lnc (leading strand) folyamatosan szintetizldik, ahogy helikz enzim segtsgvel tekeredik szt az anya-DNS. A msik lncon foly szintzis szakaszos, ti. ellenkez irnyban zajlik (n. lagging strand, ksleked szl), s rvid RNS-lncok (primerek) beptse elzi meg magt a DNS-szintzist. Utlag az RNS-lncok lebomlanak, DNS-szakaszok (Okasaki-fragmentumok) szszektdsvel (enzimatikus ligcijval) helyrell a DNS folytonossga.
2.1.6.DNS mRNS (transzkripci), mRNS fehrje (transzlci)
A DNS-ben rejl informci kifejezdse, expresszija gy trtnik, hogy elszr a kifejezd szakasz (pl. egy fehrjt kdol gn) bzissorrendjvel komplementer RNS-molekula szintetizldik RNS-polimerz enzim(ek) ltal ezt a folyamatot nevezzk trsnak, transzkripcinak (2/3A bra). Ez az elsdleges (naszcens) RNSmolekula bizonyos, a 2/3B brn demonstrlt talakts (rs, processzls) utn mr mint messenger RNS (mRNS, kldnc RNS) a magbl a citoplazmba transzportldik, ahol a riboszmkhoz kapcsoldik. A riboszmk, hatalmas, fehrjkbl s rRNS- (a riboszmk struktrjhoz tartoz, riboszomlis RNS) molekulkbl ll enzimkomplexek, melyek a mRNS-molekulk bzissorrendje alapjn fehrjt szintetizlnak. Ez gy trtnik (2/3C bra), hogy minden egymst kvet bzishrmasnak (kodon) egy-egy aminosav felel meg. Pl. az UCG (az RNS-ben uridin helyettesti a DNS timinbzist) a szerin nev aminosavnak felel meg. A megfeleltetst az egyes aminosavak s bzishrmasok kztt az biztostja, hogy ugyanazon enzim hordozza a megfelel aminosavat s azt a transzfer-RNS-t (tRNS), mely megfelel bzishrmassal komplementer bzisokat (antikodon) tartalmaz. gy a tRNS a megfelel mRNS-bzishrmashoz vezeti ezt az enzimet. Ezltal minden mRNS-bzishrmashoz, az enzim kzbeiktatsval, egy adott aminosav kapcsoldik. Az aminosavakbl, peptidktsek rvn, polipeptidlnc keletkezik. Az utbbi aminosavszekvencija tkrzi teht az mRNS, ill. a DNS bzissorrendjt. Az egsz lvilgra kiterjed, univerzlis, bzishrmas aminosav megfeleltets a genetikai kd (2/3C bra).
2.1.5. DNS-repair
A Darwin ltal sokrten s maradand hitelessggel bizonytott evolcis alaptrvny, a vltozkonysg + szelekci elve sejtpopulcik viselkedst is vezrli, in vitro s in vivo egyarnt. A sejt genetikai stabilitsa pontosan szablyozott, egyrszt a DNS-szintetizl enzimappartusok hibaszzalka szintjn, msrszt a statisztikailag elfordul beplsi hibkat s a krnyezeti felttelek ltal elidzett (sugrzs, reaktv oxigngykk) bzissorend-vltozsokat, mutcikat kijavt mechanizmusok rvn. A sejt hatkony mechanizmusokkal rendelkezik mindenfle DNS-szerkezet-mdosuls kijavtsra (repair). Folytonossghinyok, egy vagy mindkt szlon bekvetkezk, perceken bell a reparciban rszt vev fehrjk helysznre vndorlst, odaktdst vltjk ki. E fehrjk egy rsze in vitro is szabadvg-kt tulajdonsg, msoknak, pl. a nukleznak enzimatikus (DNS-hast) aktivitsuk van, egyes fehrjk ms rsztvevket foszforillni kpesek. A soklpses folyamatot fehrjefehrje kapcsolatok integrljk.
Forrs: http://www.doksi.hu
68 A SEJT LEGFONTOSABB ANYAGI SSZETEVI S ALAPVET MOLEKULRIS MECHANIZMUSAI. A SEJTBIOLGIA MOLEKULRIS BIOLGIAI ESZKZTRA.
1. transzkripci
DNS RNS-polimerz tRNS aminosavak mRNS rRNS RNS nukleotidok fehrjk
antikodon
polipeptidlnc magmembrn riboszma kodon
2. transzlci
mRNS
B
PROKARITK
DNS TRANSZKRIPCI mRNS TRANSZLCI fehrje
ser
tyr
tRNS
EUKARITK
DNS
citoplazma sejtmag
intronok exonok
U C A A G U
antikodon kodon
A U G U A C
mRNS
a msodik bzis a kodonban a harmadik bzis a kodonban
gn TRANSZKRIPCI
az els bzis a kodonban
elsdleges RNS transzkriptum RNS sapka
U U C A G
Phe Phe Leu Leu Leu Leu Leu Leu Ile Ile Ile Met Val Val Val Val
C
Ser Ser Ser Ser Pro Pro Pro Pro Thr Thr Thr Thr Ala Ala Ala ala
G
U C A G U C A G U C A G U C A G
mRNS
5 sapka s poli-A farok kpzdse AAAA RNS-rs AAAA EXPORT
mRNS fehrje
AAAA TRANSZLCI
Cys Tyr Cys Tyr STOP STOP STOP Trp Arg His Arg His Arg Hln Arg Gln Ser Asn Ser Asn Arg Lys Arg Lys Gly Asp Gly Asp Gly Glu Gly Glu
2/3. bra. A fehrjk szintzisnek legfontosabb lpsei (A, B) s a genetikai kd (C).
Forrs: http://www.doksi.hu
A SEJT LEGFONTOSABB ANYAGI SSZETEVI S ALAPVET MOLEKULRIS MECHANIZMUSAI
69
2.1.7. A naszcens RNS mRNS-s rse
A szintetizldott RNS tovbbi talakulsokon megy keresztl (l. 2/3B. bra). Elsnek beplt 5 vgt biokmiai folyamatok mdostjk (5 cap), 3 vghez sok azonos A-bl poliA farok szintetizldik. Mivel a gnek kdol (exon) s ezeket elvlaszt nem kdol (intron) szakaszokbl plnek fel, a nem kdol (de trt) mRNSszekvencik utlag kivgdnak, enzimatikusan kicspdnek (splicing) az mRNS-molekulkbl. Ezt a funkcit proteinkomplexek (splicesomes) ltjk el, melyek alegysgei kis, a nukleoplazmban sztszran kimutathat RNSprotein komplexek (snRNP-k) formjba mutathatk ki. Lehetsg van egy gn klnbz exonjainak alternatv felhasznlsra is (alternatv splicing). Ennek rvn egy gn tbbfle fehrjt is kdolhat. (Egyes gntermkek esetben az mRNS bzissorrendjnek utlagos megvltozsa is elfordul ez az RNS-editls jelensge, mely eddigi ismereteink szerint nem tartozik a sejt alapvet szablyozsi folyamatai kz.) A jelenlv mRNSkoncentrci rszben a termeldshez vezet lpsek sebessgmeghatroz mozzanattl, rszben lebomlsa szablyozott temtl fgg.
gomerizcis viszonylatok befolysoljk. A posztszintetikus modifikcik kztt igen jelents a foszforilci: az n. kinz enzimek negatv tlts foszftcsoport kovalens felktdst katalizljk egy msik fehrjre, mely abban jelents konformcivltozst idz el. Ez a vltozs azonban reverzibilis, mert egy megfelel foszfatz enzim a foszftcsoport lehasadsnak kedvez reakciutat nyithat. Egy msik jelents posztszintetikus reakci a ubikvitinci, melynek sorn egy proteinbont enzimek (protezok) ltali degradcira sznt fehrje egy kis mret fehrjvel, az ubikvitinnal konjugldik bonyolult enzimappartus ltal.
2.1.9. A gnexpresszi szablyozsa
A gnexpresszi elssorban az trs szintjn szablyozdik, s egyrszt az aktulis anyagcsere sttusznak s kls krlmnyeknek megfelel pillanatnyi alkalmazkodst, msrszt komplex sejtllapot-mdosulsokat tesz lehetv. Az utbbira akkor van szksg, amikor a sejt szveti sajtossgai vltoznak, egy adott szvetflesgre jellemz gnexpresszis mintzat alakul ki. Ez megtrtnhet in vitro (vegben, vagyis ksrleti krlmnyek kztt) s in vivo (az lben); ltalban ezeket a komplex folyamatokat differencicinak nevezzk. A sejt in vivo differencildik az egyedfejlds (a kifejlett llny sokfle szvetnek egyetlen megtermkenytett petesejtbl val kialakulsa; ontogenezis) sorn vagy a kifejlett llnyben zajl regenercis s szveti differencicis mkdsek kapcsn. A kialakult komplex funkcik sejtgenercirl genercira val megrzst a gnexpresszis mintzat propagldsa (epigenetikus rkldse) biztostja. Az eukarita gnek RNS-be val trsa a gnnel sszefgg DNS-terleten elhelyezked n. cisz-elemektl, ill. ezekhez kapcsold fehrjktl, valamint a kizrlag az rintett gnszakaszhoz kapcsold fehrjefaktoroktl fgg az
2.1.8. A fehrjk posztranszlcis mdostsai s clba juttatsa
Az jonnan szintetizlt fehrjk a szekvencijukat kpez ksbb enzimatikusan kivgott vagy a clba juttatott fehrje ltal is hordozott elemek s az ezekkel kapcsolatba lp clz mechanizmusok klcsnhatsa rvn kerlnek felhasznlsuk helyre (targeting). Jelenlv mennyisgket termeldsk s szablyozott lebomlsuk egyenslya lltja be, enzimatikus mkdsket posztszintetikus modifikcik s ms fehrjkkel val vltozatos klcsnhatsok, oli-
Forrs: http://www.doksi.hu
70 A SEJT LEGFONTOSABB ANYAGI SSZETEVI S ALAPVET MOLEKULRIS MECHANIZMUSAI. A SEJTBIOLGIA MOLEKULRIS BIOLGIAI ESZKZTRA.
2/4. bra. A gnkifejezds szablyozsa a transzkripci szintjn.
utbbiakat transz-faktoroknak nevezzk. A cisz-elemek kz tartozik a promter, amely a gn mRNS-t kdol szakasztl 5 irnyban helyezkedik el, kb. 100-200 bp hossz, s DNS-kt fehrjk szmra biztost kthelyet. A promter vezrli a transzkripcit (az mRNS trst) az mRNS-t szintetizl RNS-polimerz enzim ktdse s elindulsa feltteleinek biztostsval. A legtbb promterben elfordul promterelem az n. TATA box. Ez egy T-A-T-A nukleotidsorrend szakasz, ami 35 bp-nyira (35) helyezkedik el a transzkripci starthelytl (+1). Ehhez ktdik a TATA-kt fehrje (TBP), amelyhez tovbbi ltalnos transzkripcis faktorok [pl. a TFIIDkomplex = TBP + TAFok (TBP Associated Factors)] kapcsoldnak. Ezek egyttesen biztostjk az RNS-polimerz mkdst s a transzkripci elindulst. (Az ltalnos transzkripcis faktorok az RNS-polimerzzal egytt (ami maga is tbb fehrjbl ll) alkotjk az alaptranszkripcis appartust.) A gnexprsszi szablyozsnak* tovbbi fontos cisz-elemei az n. enhanszerek (2/4. bra). Ezek a gn kdol szakasztl akr 5 akr 3 irnyban helyezkedhetnek el, s transzkripcis faktorok kthelyeibl, az n. vlaszad elemekbl llnak. Az enhanszerek ltalban 10-50 bp hosszsg, mindkt orientciban mkdkpes
*
szakaszok. Igen tvol, akr tbb ezer bzisprnyira is elhelyezkedhetnek a befolysolt gntl. Az enhanszerekhez kapcsold transzkripcis faktorok pozitv s/vagy1 negatv irnyban tudjk befolysolni a promter mkdst (az utbbi esetben silencer a nevk). A specifikus transzkripcis faktorok DNS-kt domnjkn2 kvl rendelkeznek egy aktivl s/vagy gtl domnnel is. Mivel ezek a fehrjk ltalban nem lpnek direkt kapcsolatba a promterhez kapcsold fehrjkkel, kzvett fehrjkre van szksg a mkdskhz. Ezeket kofaktoroknak nevezzk: koaktivatoroknak vagy korepresszoroknak, attl fggen, hogy aktivl vagy gtl szerepet tltenek be. Ezek ltalban egy nagyobb fehrjekomplex rszeknt biztostjk a fizikai kapcsolatot az enhanszerhez kapcsold, szekvenciaspecifikus transzkripcis faktor s a promterhez ktd ltalnos transzkripcis faktorok kztt. Ezeknek a komplexeknek tbbfle enzimatikus aktivitsuk is van, gy hiszton-acetilz, deacetilz s esetleg metilz, demetilz vagy kinz s ATP-fgg n. kromatin remodelling aktivitssal, mellyel hozzjrulnak a kromatinszerkezet talaktshoz s a transzkripci sebessgnek nvelshez vagy ppen cskkentshez. A 2/4. bra az eukarita transzkripci szablyozsnak alapelveit mutatja
1
Pl. a magreceptorok ligand (hormon) hinyban gtolnak, jelenltben aktivlnak.
Kis, duplaszl RNS-ek gnexpresszi-szablyozsban betlttt szereprl egyre tbb adat lt napvilgot. Ezen jfajta szablyozs mechanizmusrl mg nincsenek biztos ismereteink.
A domn egy fehrje krlhatrolhat szerkezeti s funkcionlis rszlete.
Forrs: http://www.doksi.hu
A SEJT LEGFONTOSABB ANYAGI SSZETEVI S ALAPVET MOLEKULRIS MECHANIZMUSAI
71
vzlatosan. A TATA boxszal br promterhez kapcsoldik az alap transzkripcis appartus. Ez biztostja az RNS trdst. Ezt az aktivitst modulljk enhancerek-silencerek a hozzjuk kapcsold specifikus transzkripcis faktorokon keresztl. Az brn kofaktorokat s hozzjuk kapcsold fehrjket nem tntettnk fel.
E. coli
10 20 30 40 50 60
a b
BT
2.1.10. ltalnos szablyozsi alapelvek
Az anyagcsere-folyamatok, ill. a differencici szablyozsnak egyik fontos alapelve a visszacsatols (feed-back). Ennek fleg negatv visszacsatolsknt ismert vltozata gyakori, pl. amikor egy keletkezett termk gtolja az t ltrehoz folyamatot. Az enzimatikus reakcik sorbakapcsolsuk rvn kaszkdfolyamatokk rendezdhetnek, melyek sorn egy-egy enzim tovbbiak sokasgt aktivlva robbansszer vltozsokat hozhat ltre a vgs szubsztrt szintjn. Gyakori szablyozsi megolds a trigger-elv: egy folyamatsor mintegy ugrsra kszen vrja azt a stimulust, mely, mint a ravasz meghzsa a goly kireplst (melynek gyorsasga nem fgg a ravasz meghzsnak erejtl), idzi el a reakcilnc beindulst.
70 80 90 100 10
-3
10
-2
10
-1
10
10
10
10
C0 t
2/5. bra. A prokaritk (E. coli) s emls- (borj thymus, BT) DNS reasszocicis viselkedse.
2.1.11. A DNS heterogenitsa
A kromoszmamret DNS rtelmes szlrl trsra kerl, a diploid genomban kt alllban elfordul egyedi gneket (2/5. bra, c) s klnbz mrtkben (a, b) ismtld, a genomban tandem s sztszrtan, sok-sok msolatban jelenlv szekvencikat tartalmaz. A klnbz szekvencik szma a genom n. komplexitsa, mely nagymrtkben klnbzik a fajok kztt. A gnek (s a repetitv elemek egyarnt) egyedi (nem ismtld), nem kdol szakaszokkal (spacerekkel) hatroldnak el egymstl. A spacerek, valamint a gnek intronjai, s az n. pszeudognek (RNS-rl visszafel, DNS-be rt szekvenci-
k) nem kdolnak fehrjt. A denaturlt (>8090 oC-on egyszlv vl) DNS reasszocicis kinetikja tkrzi ezt a heterogenitst. Mint az brn lthat, a bakterilis, ill. eukarita (borj csecsemmirigy, thymus, BT) DNS reasszocicis viselkedse kevesebb, ill. tbb (a teljes DNS-llomnynak jelentsebb rszt alkot) ismtld szekvenciaarnyra utal. A C 0t rtk a DNS (nukleotidokra szmolt) molris koncentrcijnak s az eltelt idnek a szorzata, melynek fggvnyben a reasszocici %-os mrtkt brzoljuk. A genom DNS-szekvencii bzissszettelk (AT/GC arnyuk) szerint is tbb klnbz alosztlyt alkotnak. Az utbbi arny szempontjbl lesen elklnlnek a gnek s a transzkripci szempontjbl inaktv terletek. Ezek vltakozsa az eddig ismertt vlt hosszabb, egybefgg DNS-szekvencik alapjn nagyjbl hurokmret periodicitst mutat. Az ismtlds foka szerint hromfle szekvenciakategrit klnbztetnek meg: ersen repetitv (egyszer, tandem ismtld, rvid szekvencikbl ll nagyobb szakaszok, mint pl. a kromoszmk n. centromerikus rgiinak n. szatellita DNS-e), a kzepesen repetitv s a nem ismtld (egyedi) szakaszokbl ll genomh-
Forrs: http://www.doksi.hu
72 A SEJT LEGFONTOSABB ANYAGI SSZETEVI S ALAPVET MOLEKULRIS MECHANIZMUSAI. A SEJTBIOLGIA MOLEKULRIS BIOLGIAI ESZKZTRA.
nyad (pl. gnek tbbsge). A repetitv szekvencik alkotjk a genom tbb mint felt, jelentsgkre nincs meggyz magyarzat3. Kiemelend a DNS sajt mobilitsnak tnye: olyan rvid szekvenciaelemek lteznek, melyek rvn hoszszabb DNS-szakaszok is nll tra kelhetnek a magon (sejten, ill. populcin) bell, s eredeti helykhz kpest mshova plhetnek be (inszertldhatnak) mRNS-szintje klnbz sejtekben s szvetekben. Gyorsan trnek elre a szintn hibridizcin alapul miniatrizlt n. chiptechnikk. Ilyen a DNS-mikroarray, melynek sorn tbb ezer gnt jellemz DNS-molekula kerl felvitelre egy kicsiny trgylemezre, s ehhez hibridizljk egy adott sejt teljes RNS-kszletnek megfelelen jellt DNS msolatt (komplementer DNS, cDNS). Ezltal egyetlen ksrletben tbb ezer gn expresszis szintje hatrozhat meg. A fehrjk esetben a primer szekvencin alapul hibridizcira nincs lehetsg. Ezzel szemben jl kihasznlhatk a specifikus antignantitest klcsnhatsok fehrjk kimutatsra s jellsre. A fentiekhez hasonl, de fehrjk kimutatsra alkalmas az n. Western blot technika: ennek sorn sejtekbl, szvetekbl nyert fehrjket vlasztunk el mret szerint glelektroforzissel. Szilrd hordoz membrnra transzferljuk (blottoljuk) a sztvlasztott fehrjket, s egy adott fehrje ellen termeltetett specifikus antitesttel kimutatjuk. Ez ltalban gy trtnik, hogy az antitesthez egy enzimet rgztenek, s az enzim ltal katalizlt sznreakci jelenti meg a fehrjt. Ezzel a mdszerrel adott fehrje jelenlte s szintje hatrozhat meg sejt- vagy szvetkivonatokban. A sejtbiolgusok szmra azonban ennl ltalban fontosabb egy fehrje sejten belli (intracellulris) elhelyezkedsnek meghatrozsa. Erre is jl hasznlhat az immunhisztokmia. Ekkor szintn egy adott fehrjre specifikus antitestet hasznlunk, de ebben az esetben izollt sejteken vagy szveti metszeteken vizsgljuk az adott fehrje elhelyezkedst. Az antitesteket gyakran fluoreszcens anyagokkal jellik, s megjelentskhz fluoreszcens mikroszkpit hasznlnak. Ez a mdszer kivlan alkalmas fehrjk sejten, illetve szveten belli elhelyezkedsnek vizsglatra. Fehrjemolekulk klcsnhatsait, adott fehrjkhez kapcsold fehrjket lehet kimutatni a szintn antitestek hasznlatn alapul immunprecipitcis technikval. Ebben az esetben ltalban sejtkivonatokbl csapjuk ki s gyjtjk ssze az antitestekkel kapcsoldni kpes fehrjket. Ezek minsge s mennyisge tovbbi vizsg-
2.2. A sejtbiolgia molekulris biolgiai eszkztra
A molekulris biolgia s mdszereinek gyors fejldse nagy hatssal volt s van a sejtbiolgira. Az albbiakban rviden ismertetjk, a teljessg ignye nlkl, azokat a molekulris biolgiai mdszereket, melyek dnten hozzjrultak a sejtek mkdsnek megismershez.
2.2.1. Mdszerek
A nukleinsavak kmiai termszetbl add tulajdonsga, hogy a komplementarits elve alapjn prosthatak: hibridizlnak (l. a 2/5. brn a DNS reasszocicis viselkedst). Ezt j nhny technika kihasznlja. A Southern hibridizci esetn (2/6. bra) az agarzglen mretk szerint elvlasztott DNS-molekulkat szilrd hordozra transzferljk, azon rgztik, s radioaktvan jellt szondval4 hibridizltatjk (DNS-DNS hibridizci). A Northern hibridizci a Southernhez hasonl, de RNS-molekulk sztvlasztsn s DNS-szondval val jellsn alapszik. Segtsgvel meghatrozhat pldul egy adott gn
3 Ez az n. C-rtk paradoxon (a C-rtk a haploid genom kdolt gnekt meghalad DNS-tartalma).
Angolul probe, ezrt magyarul sokszor prbnak fordtjk.
Forrs: http://www.doksi.hu
A SEJTBIOLGIA MOLEKULRIS BIOLGIAI ESZKZTRA
73
restrikcis endonuklezzal emsztett jelletlen DNS
paprtrlkz-kteg nitrocellulz membrn
a nitrocellulz filter eltvoltsa a szorosan hozzkttt DNS-sel
jellt, ismert mret DNS-darabok
agarzgl szivacs alkalikus oldat
DNS-fragmentek elvlasztsa agarzgl-elektroforzissel
az elvlasztott DNS-fragmentek blottolsa nitrocellulz filterre
pufferben oldott jellt DNS-prba lezrt plasztikzacsk
jellt svok
jellt DNS prba hibridizlsa az elvlasztott DNS-fragmentekhez
a jellt markerek elhelyezkedse
a komplementer DNS svokhoz hibridizlt jellt DNS prba detektlsa autoradiogrfival
2/6. bra. A Southern-blot technika.
latokkal (pl. Western blottal) meghatrozhat. A mdszer kivlan alkalmas fehrjk partnereinek tiszttsra, illetve fehrjekomplexek tanulmnyozsra.
2.2.2. Modellllnyek
A sejtbiolgiai folyamatok megrtshez elengedhetetlen egyszerbb modellszervezetek hasznlata. Ilyen az egyik legegyszerbb eukarita szervezet, a kenyr- vagy srleszt (Saccharomyces cerevisiae), egy egysejt gomba, ami legalbb annyira kzeli rokona a nvnyeknek, mint az l-
latoknak. Kedvez krlmnyek kztt olyan gyorsan szaporodik, mint a baktriumok. Mivel DNS-tartalma csak kt s flszer nagyobb mint az Escherichia coli (E. coli) baktrium, kivlan hasznlhat genetikai ksrletekre is. Az leszt kivlan alkalmas olyan alapvet intracellulris folyamatok vizsglatra, mint a sejtorgaellumok mkdse, sejtciklus, szignltranszdukci, gntrds szablyozsa, DNS-replikci s repair, anyagcsere-folyamatok. Termszetesen az leszt nem alkalmas a soksejt llnyek sajtsgainak modellezsre (sejtsejt kommunikci, differencilds). Ez utbbiak tanulmnyozsra az ecetmuslica (Drosophila melanogaster) s egy nematodk kz tartoz freg, a Caenorrhabditis elegans
Forrs: http://www.doksi.hu
74 A SEJT LEGFONTOSABB ANYAGI SSZETEVI S ALAPVET MOLEKULRIS MECHANIZMUSAI. A SEJTBIOLGIA MOLEKULRIS BIOLGIAI ESZKZTRA.
vlt a legkiterjedtebben hasznlt ksrleti objektumm. Az ecetmuslica a genetikusok egyik kedvelt ksrleti llata, s nagyban hozzjrult olyan alapvet biolgiai folyamatok megrtshez, mint a differencilds folyamata, a test szelvnyezettsgnek kialakulsa s annak szablyozsa. A C. elegans petesejtje rampontossggal alakul a megtermkenyts utn egy pontosan 959 sejtbl ll soksejt szervezett. E folyamat pontos megfigyelse s lersa, valamint az ehhez kapcsold genetikai vizsglatok sokat adtak az idegrendszerrl, differencildsrl, apoptzisrl meglv ismereteinkhez. Kitntetett szerepet jtszik a biolgiai kutatsokban az egr (Mus musculus). Az egr a legtbbet tanulmnyozott emls az ember mellett. Viszonylag rvid reprodukcis ideje (21 nap), beltenysztett, genetikailag identikus trzseinek genetikai manipullhatsga s egyszer tenysztse teszi vonz kutatsi clpontt. Kt technolgia, mellyel knnyen s specifikusan megvltoztathatv vlt az egr genomja, forradalmastotta a biolgiai megismers folyamatt. Az egyik eljrs segtsgvel n. transzgenikus llatokat lehet ltrehozni a megtermkenytett petesejtbe val DNS-injektlssal. Ebben az esetben az injektlt DNS bepl a genomba, s ott kifejezdik. Az ilyen mdon ltrehozott llatok propagljk a stabilan integrldott DNS-t nemzedkrl nemzedkre. Ilyen mdon in vivo tanulmnyozhatv vlik egy kivlrl bevitt gn hatsa. Ezt a technolgit kiegszti a nullmutci ltrehozsnak technikja (knock out technika). Ebben az esetben egy adott, vizsglni kvnt gn kicserlse trtnik meg homolg rekombinci (szekvenciahomolgin alapul DNS-szakasz-kicserlds) segtsgvel embrionlis ssejtekben (ES) in vitro. Ezekbl az ssejtekbl amennyiben azokat blasztocisztkba ltetik (pl. mikroinjektljk) olyan kimraegerek szletnek, melyek egyes sejtjei vagy szvetei a manipullt ESsejtekbl szrmaznak. Ezekbl, ha a kimrizmus az ivarsejteket is rinti, keresztezssel ltrehozhat homozigta null mutns, melynek minden sejtje tartalmazza a mutcit. A transzgn s knock out technikval jl vizsglhat egy-egy gn szerepe az organizmus fejldse s lete sorn.
Вам также может понравиться
- Orvosi BiokémiaДокумент660 страницOrvosi BiokémiaRobert Ohler80% (10)
- Fonyó Attila, Ligeti Erzsébet - Az Orvosi Élettan TankönyveДокумент740 страницFonyó Attila, Ligeti Erzsébet - Az Orvosi Élettan Tankönyveszazalek100% (5)
- 33 kedvenc recept: Bevezetés az egészséges táplálkozásbaОт Everand33 kedvenc recept: Bevezetés az egészséges táplálkozásbaРейтинг: 5 из 5 звезд5/5 (1)
- Bánhegyi Gábor, Sipeki Szabolcs - Biokémia, Molekuláris És Sejtbiológia PDFДокумент550 страницBánhegyi Gábor, Sipeki Szabolcs - Biokémia, Molekuláris És Sejtbiológia PDFÁgnes Zongor100% (1)
- Sejtbiológia Könyv Elte PDFДокумент172 страницыSejtbiológia Könyv Elte PDFHorvathBrigittaОценок пока нет
- Ross - SzövettanДокумент871 страницаRoss - SzövettanMajorosNikolett100% (6)
- A Biokemia Es A Molekularis Kemia Alapjai READERДокумент555 страницA Biokemia Es A Molekularis Kemia Alapjai READERTelek Andrs100% (1)
- Anatomia Essentialis 1Документ203 страницыAnatomia Essentialis 1Dóra Szentkirályi100% (3)
- Dr. Ormai Sándor - Élettan - KórélettanДокумент346 страницDr. Ormai Sándor - Élettan - Kórélettanr2d2robi100% (3)
- Dr. Németh Endre - Biológia Kézikönyv (OCR) PDFДокумент304 страницыDr. Németh Endre - Biológia Kézikönyv (OCR) PDFMedstud100% (3)
- Bazsa-Jegyzet Beta2Документ252 страницыBazsa-Jegyzet Beta2nrita72100% (2)
- Biokémia Kidolgozott TételekДокумент26 страницBiokémia Kidolgozott TételekSzénási Kriszti100% (3)
- Biolemia AlapjaiДокумент563 страницыBiolemia AlapjaiAdrienn Markesz100% (2)
- Anyacsavar és kockafej: A fiam autista, de nem zseniОт EverandAnyacsavar és kockafej: A fiam autista, de nem zseniРейтинг: 5 из 5 звезд5/5 (5)
- Orvosi BiokémiaДокумент356 страницOrvosi BiokémiaZsuzsanna MezeiОценок пока нет
- Kutatási Módszerek a Pszichológiában és a Társadalomtudományokban: Empirikus Társadalomkutatás, Kutatási Módszerek és Értékelés Egyszerűen MagyarázvaОт EverandKutatási Módszerek a Pszichológiában és a Társadalomtudományokban: Empirikus Társadalomkutatás, Kutatási Módszerek és Értékelés Egyszerűen MagyarázvaОценок пока нет
- Bevezetes Az Immunologiaba PDFДокумент84 страницыBevezetes Az Immunologiaba PDFVeres Zsuzsi100% (1)
- MikrobiologiaДокумент669 страницMikrobiologiaMariannCsászárОценок пока нет
- Szabó Gábor - SejtbiológiaДокумент795 страницSzabó Gábor - SejtbiológiaMáté Vas100% (3)
- Humán SzövettanДокумент66 страницHumán SzövettanKatalin GulácsyОценок пока нет
- 4 - Élettan - 2013 - Hetes JegyzetДокумент589 страниц4 - Élettan - 2013 - Hetes Jegyzetnrita72Оценок пока нет
- Szeberenyi Molekularis Sejtbiologia PDFДокумент377 страницSzeberenyi Molekularis Sejtbiologia PDFMonika FurkoОценок пока нет
- MK Szövettan JegyzetДокумент238 страницMK Szövettan JegyzetManka100% (1)
- Alt KortanДокумент31 страницаAlt KortanZoli Künsztler100% (1)
- 0 SzövettanДокумент42 страницы0 SzövettanZoi KalaitziОценок пока нет
- Tanuljon Szerbül - Gyorsan / Egyszerűen / Hatékonyan: 2000 KulcsszóОт EverandTanuljon Szerbül - Gyorsan / Egyszerűen / Hatékonyan: 2000 KulcsszóОценок пока нет
- Sejtalkotok JДокумент50 страницSejtalkotok Jangelika6ill6s100% (1)
- Cytológia És Szövettan Kidolgozott PDFДокумент172 страницыCytológia És Szövettan Kidolgozott PDFLevente Balázs100% (1)
- HaematologiaДокумент82 страницыHaematologiaFanni Anna Domonkos100% (2)
- Biológia JegyzetДокумент412 страницBiológia JegyzetDnaielОценок пока нет
- SzovettanДокумент126 страницSzovettanDrBenjaminButtonОценок пока нет
- Kidolgozott Immunológia KérdésekДокумент15 страницKidolgozott Immunológia KérdésekAzt Hiszem Ez Mindegy100% (1)
- Alapszakdolgozatok, Mesterszakdolgozatok, Diplomamunkák, Szakdolgozatok, Tudományos Megírása Könnyedén (Tudományos Munka)От EverandAlapszakdolgozatok, Mesterszakdolgozatok, Diplomamunkák, Szakdolgozatok, Tudományos Megírása Könnyedén (Tudományos Munka)Оценок пока нет
- ÉlettanДокумент44 страницыÉlettanhermirekaОценок пока нет
- Biofizikia-Kidolgozott-K RD Sek - Szi-F L VДокумент23 страницыBiofizikia-Kidolgozott-K RD Sek - Szi-F L VKatalin Strobl100% (1)
- Elettan Idegrendszer 2005Документ65 страницElettan Idegrendszer 2005Szasz Helen HenriettaОценок пока нет
- Mikrobi TesztekДокумент28 страницMikrobi TesztekCsizmadia Livia100% (1)
- Sejtbiológia 1Документ57 страницSejtbiológia 1kereklovagОценок пока нет
- Biofizika TételekДокумент75 страницBiofizika TételekVas Lilla100% (1)
- hétköznapi szócséplő: unalomtól megmentő rímeket rittyentőОт Everandhétköznapi szócséplő: unalomtól megmentő rímeket rittyentőОценок пока нет
- Biologia ReszletesДокумент111 страницBiologia ReszletesNahimi Csanád100% (1)
- Traumatologia Jegyzet - NoverkepzoДокумент83 страницыTraumatologia Jegyzet - NoverkepzoSzasz Helen HenriettaОценок пока нет
- Kemia Ts Erettsegi EmeltДокумент69 страницKemia Ts Erettsegi EmeltspaОценок пока нет
- Sejbiologia3 5 2009Документ44 страницыSejbiologia3 5 2009szecska5Оценок пока нет
- 2zh ÉlettanДокумент4 страницы2zh ÉlettanHajnika Bancsok100% (1)
- 2011 0079 Wunderlich Szarka BiokemiaДокумент234 страницы2011 0079 Wunderlich Szarka BiokemiafakkerОценок пока нет
- Panda - BiokémiaДокумент69 страницPanda - BiokémiaAngiОценок пока нет
- RuthДокумент54 страницыRuthSzasz Helen HenriettaОценок пока нет
- ÁllatélettanДокумент201 страницаÁllatélettanszpista100% (1)
- ÁllattanДокумент28 страницÁllattanxspatulaxОценок пока нет
- Sejtbiológia I. ZHДокумент18 страницSejtbiológia I. ZHMárton HorváthОценок пока нет
- Jabec Imaja SzerkДокумент2 страницыJabec Imaja SzerkSzasz Helen HenriettaОценок пока нет
- Előtted Az ÉletДокумент133 страницыElőtted Az ÉletbarackoscsokiОценок пока нет
- Előtted Az ÉletДокумент133 страницыElőtted Az ÉletbarackoscsokiОценок пока нет
- Aminosavak AcsДокумент40 страницAminosavak AcsSzasz Helen HenriettaОценок пока нет
- TetelДокумент10 страницTetelSzasz Helen HenriettaОценок пока нет
- FestesekДокумент2 страницыFestesekSzasz Helen HenriettaОценок пока нет
- 37 TetelДокумент18 страниц37 TetelSzasz Helen HenriettaОценок пока нет
- 38 TetelДокумент2 страницы38 TetelSzasz Helen HenriettaОценок пока нет
- 41 TetelДокумент7 страниц41 TetelSzasz Helen HenriettaОценок пока нет