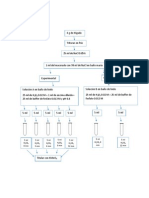
Вам также может понравиться
- Ecoli Bacterias Termo PDFДокумент1 страницаEcoli Bacterias Termo PDFraul1801Оценок пока нет
- PeroxidoДокумент1 страницаPeroxidoraul1801Оценок пока нет
- Gluconeogénesis, Síntesis y Degradacion de Glucógeno y Vía de Las Pentosas de Fosfato - 2Документ15 страницGluconeogénesis, Síntesis y Degradacion de Glucógeno y Vía de Las Pentosas de Fosfato - 2raul1801Оценок пока нет
- RMN & Rayos-XДокумент10 страницRMN & Rayos-Xraul1801Оценок пока нет
- Métodos Extracción ADNДокумент5 страницMétodos Extracción ADNraul1801Оценок пока нет
- 4 AphycomplexasДокумент0 страниц4 Aphycomplexasraul1801Оценок пока нет
- Membranas 2Документ8 страницMembranas 2raul1801Оценок пока нет
- Membranas Transporte 2Документ21 страницаMembranas Transporte 2raul1801Оценок пока нет
- Tema 6. Las RodófitasДокумент24 страницыTema 6. Las Rodófitasraul1801Оценок пока нет
- ManualДокумент218 страницManualraul1801Оценок пока нет
- Tema 13b. Monilophyta - Marattiopsida - PolypodiopsidaДокумент45 страницTema 13b. Monilophyta - Marattiopsida - Polypodiopsidaraul18010% (1)
- CelenteradosДокумент2 страницыCelenteradosraul1801Оценок пока нет
- Degradación de Polifenoles Por Procesos de Oxidación AvanzadaДокумент15 страницDegradación de Polifenoles Por Procesos de Oxidación AvanzadaRicardo GamarraОценок пока нет
- Ejercicios de OrganicaДокумент2 страницыEjercicios de OrganicaMayra MartinezОценок пока нет
- Evaluación Continua 1 Arnold AlejandroДокумент2 страницыEvaluación Continua 1 Arnold AlejandroMrChemy3Оценок пока нет
- Accion de Una Enzima de Tejidos Vegetales y AnimalesДокумент10 страницAccion de Una Enzima de Tejidos Vegetales y AnimalesJodie Rey Romero100% (1)
- MetanolДокумент14 страницMetanolTony PalacioОценок пока нет
- Guía #2 Cinética Enzimática 2020Документ6 страницGuía #2 Cinética Enzimática 2020tomas contrerasОценок пока нет
- Polimerización de Etileno y Esencia de Trementina Con Un Sistema Catalítico Tipo Ziegler PDFДокумент71 страницаPolimerización de Etileno y Esencia de Trementina Con Un Sistema Catalítico Tipo Ziegler PDFNicandroGonzalesОценок пока нет
- Estudio de Los Alquinos-Reacciones QuímicasДокумент18 страницEstudio de Los Alquinos-Reacciones QuímicasCAMILA KARELIS HURTADO ILIGUANОценок пока нет
- Biocatálisis Clase 1 PDFДокумент19 страницBiocatálisis Clase 1 PDFAlejandraОценок пока нет
- Métodos de limpieza y reutilización de catalizadores automotricesДокумент5 страницMétodos de limpieza y reutilización de catalizadores automotricesLuis Javier Montes SalazarОценок пока нет
- Trabajo 2Документ2 страницыTrabajo 2Lizeth Martinez RamirezОценок пока нет
- HIDROGENOДокумент88 страницHIDROGENOJayRo HdezОценок пока нет
- Manual VictorДокумент84 страницыManual VictorJose Eusebio Caro Pacheco80% (5)
- Unidad IV - Parte 2Документ17 страницUnidad IV - Parte 2altagraciaОценок пока нет
- Tecnicas de Analisis y Caracterizacion de MaterialДокумент16 страницTecnicas de Analisis y Caracterizacion de MaterialSergio AlexanderОценок пока нет
- Proyecto de MetanolДокумент55 страницProyecto de MetanolJean Wilmer G HОценок пока нет
- Resumen Analitico en Educacion RaeДокумент8 страницResumen Analitico en Educacion RaeJorge Alberto Carrillo BernalОценок пока нет
- Transferencia de masa en problemas de difusión y reacciónДокумент13 страницTransferencia de masa en problemas de difusión y reacciónKatherine CuevaОценок пока нет
- Hidrólisis y fermentación de biomasaДокумент2 страницыHidrólisis y fermentación de biomasaRicardo HernandezОценок пока нет
- Tarea 2.0 PDFДокумент8 страницTarea 2.0 PDFJean MarcoОценок пока нет
- Estil BencenoДокумент9 страницEstil BencenoMoises Valdivia BaldomeroОценок пока нет
- Syllabus EIQ447 2s 2019Документ7 страницSyllabus EIQ447 2s 2019Elías SepúlvedaОценок пока нет
- Informacion de YesoДокумент14 страницInformacion de YesoShaolin FrankОценок пока нет
- 5to Informe de Laboratorio de Quimica OrganicaДокумент14 страниц5to Informe de Laboratorio de Quimica OrganicaKatty AtocheОценок пока нет
- Determinación de azufre en hidrocarburosДокумент8 страницDeterminación de azufre en hidrocarburosBrenda Rubí Hdz BetancourtОценок пока нет
- Saúl Uriel Corona Bautista Problemas de Cinética EnzimáticaДокумент10 страницSaúl Uriel Corona Bautista Problemas de Cinética EnzimáticaSaúl Uriel Corona Bautista100% (1)
- ¿En Toda Reacción Enzimática Hay Liberación de Energía - Prácticas o Problemas - Andresfrn09Документ4 страницы¿En Toda Reacción Enzimática Hay Liberación de Energía - Prácticas o Problemas - Andresfrn09Kelly BenitezОценок пока нет
- Problemario Capítulo 3 Ingeniería de Procesos MicrobiológicosДокумент10 страницProblemario Capítulo 3 Ingeniería de Procesos MicrobiológicosKenia CarrilloОценок пока нет
- Tratamiento de EnzimasДокумент6 страницTratamiento de EnzimasRaithismer MarinОценок пока нет
- Reacciones de aldehídos y cetonas con aminasДокумент10 страницReacciones de aldehídos y cetonas con aminasdugamianОценок пока нет