Вам также может понравиться
- 2022 1 Ene Kornuta Et AlДокумент8 страниц2022 1 Ene Kornuta Et AlIvanaОценок пока нет
- Veterinary Microbiology: Nagah Arafat, Abdelfattah H. Eladl, Basma H. Marghani, Mohamed A. Saif, Reham A. El-ShafeiДокумент9 страницVeterinary Microbiology: Nagah Arafat, Abdelfattah H. Eladl, Basma H. Marghani, Mohamed A. Saif, Reham A. El-ShafeiSlimaneOuriacheОценок пока нет
- Wafaa Abd El-Ghany: Poultry Dis. Dept., Fac. Vet. Med., Cairo UnivДокумент70 страницWafaa Abd El-Ghany: Poultry Dis. Dept., Fac. Vet. Med., Cairo Univthanh ba matОценок пока нет
- 1 s2.0 S0264410X11000405 MainДокумент7 страниц1 s2.0 S0264410X11000405 MainJaime MunozОценок пока нет
- Control of Avian Influenza in PoultryДокумент43 страницыControl of Avian Influenza in PoultryAnjum IslamОценок пока нет
- 3.03.04 - Avian InfluenzaДокумент28 страниц3.03.04 - Avian InfluenzaAkhmalia SuniawatiОценок пока нет
- Avian Influenza Vaccines@JstorДокумент25 страницAvian Influenza Vaccines@JstorEcthelonОценок пока нет
- Deteksi Respon Antibodi Dengan Uji Hemaglutinasi Inhibisi Dan Titer Proteksi Terhadap Virus Avian Influenza Subtipe H5N1Документ6 страницDeteksi Respon Antibodi Dengan Uji Hemaglutinasi Inhibisi Dan Titer Proteksi Terhadap Virus Avian Influenza Subtipe H5N1HendroОценок пока нет
- Paper AvicolaДокумент13 страницPaper AvicolanikololitaОценок пока нет
- IFN-γ establishes interferon-stimulated gene-mediated antiviral state against Newcastle disease virus in chicken fibroblasts PDFДокумент13 страницIFN-γ establishes interferon-stimulated gene-mediated antiviral state against Newcastle disease virus in chicken fibroblasts PDFMarisol OrtizОценок пока нет
- 3355 4636 1 SM PDFДокумент7 страниц3355 4636 1 SM PDFHamdan UbaidillahОценок пока нет
- Efficacy of Commercial Vaccines Against Newly Emer PDFДокумент6 страницEfficacy of Commercial Vaccines Against Newly Emer PDFMoatazОценок пока нет
- Inhaled Nitric Oxide Therapy Fails To Improve Outcome in Experimental Severe InfluenzaДокумент6 страницInhaled Nitric Oxide Therapy Fails To Improve Outcome in Experimental Severe InfluenzaDesti EryaniОценок пока нет
- 3.03.04 AiДокумент26 страниц3.03.04 Aiivanfco11Оценок пока нет
- Fact Sheet Influenza Vaccine 2020 2021Документ8 страницFact Sheet Influenza Vaccine 2020 2021gsus17Оценок пока нет
- Long Term Persistence of Ige Anti-Influenza Virus Antibodies in Pediatric and Adult Serum Post Vaccination With Influenza Virus VaccineДокумент6 страницLong Term Persistence of Ige Anti-Influenza Virus Antibodies in Pediatric and Adult Serum Post Vaccination With Influenza Virus VaccineDesti EryaniОценок пока нет
- Neutralization Ability of Specific Antibody of Avian Influenza H5 To Several Viruses of H5N1 Field IsolatesДокумент12 страницNeutralization Ability of Specific Antibody of Avian Influenza H5 To Several Viruses of H5N1 Field IsolatesWenti Az Zahra MumtahanahОценок пока нет
- 0005-2086-47 s3 1047 PDFДокумент4 страницы0005-2086-47 s3 1047 PDFJosé María Valenzuela GraciaОценок пока нет
- Vaccine: Greg G. WolffДокумент5 страницVaccine: Greg G. WolffRennan Lima M. CastroОценок пока нет
- Virology Journal: Cross-Reactive Human B Cell and T Cell Epitopes Between Influenza A and B VirusesДокумент19 страницVirology Journal: Cross-Reactive Human B Cell and T Cell Epitopes Between Influenza A and B VirusesLateecka R KulkarniОценок пока нет
- Approaches For Vaccine Development Against Lympy Skin DiseaseДокумент12 страницApproaches For Vaccine Development Against Lympy Skin DiseaseAwais GulОценок пока нет
- LTI A ReviewДокумент22 страницыLTI A Reviewamamùra maamarОценок пока нет
- Immunizatuon ChissДокумент51 страницаImmunizatuon ChissMariam AntonyОценок пока нет
- Converted 4Документ61 страницаConverted 4miss peppa pigОценок пока нет
- Immunization DR T V RaoДокумент68 страницImmunization DR T V Raotummalapalli venkateswara rao100% (1)
- Influenza ("Bird Flu"), Inflammation and Anti-Inflammatory Analgesic DrugsДокумент8 страницInfluenza ("Bird Flu"), Inflammation and Anti-Inflammatory Analgesic DrugsAndersinclair AndersinclairОценок пока нет
- Ausiello2019 Chapter HumanImmuneResponsesToPertussiДокумент15 страницAusiello2019 Chapter HumanImmuneResponsesToPertussiAGUS DE COLSAОценок пока нет
- AvianInsightVol3 03Документ2 страницыAvianInsightVol3 03Bryan NicollОценок пока нет
- Vaccines of Poultry: October 2017Документ24 страницыVaccines of Poultry: October 2017Vinod ChaudhariОценок пока нет
- CatfluДокумент13 страницCatflumvdaniela.santanaОценок пока нет
- Adwar T., Lukešová DДокумент6 страницAdwar T., Lukešová Dmoekyaw7171Оценок пока нет
- Pathogenicity and Immunogenicity of Different Recombinant Newcastle Disease Virus Clone 30 Variants After in Ovo VaccinationДокумент10 страницPathogenicity and Immunogenicity of Different Recombinant Newcastle Disease Virus Clone 30 Variants After in Ovo VaccinationGorby CanОценок пока нет
- Newcastle Disease Virus Detection From Chicken Organ Samples Using Reverse Transcriptase Polymerase Chain ReactionДокумент14 страницNewcastle Disease Virus Detection From Chicken Organ Samples Using Reverse Transcriptase Polymerase Chain ReactionKasmira miraОценок пока нет
- AguaDoce and GracaДокумент8 страницAguaDoce and GracaLuis GracaОценок пока нет
- Khalequ Zaman, Sheikh Farzana Zaman, Farzana Zaman, Asma Aziz, Sayeed-Bin Faisal, Magali Traskine, MD Ahsan Habib, Javier Ruiz-Guiñazú, Dorota BorysДокумент9 страницKhalequ Zaman, Sheikh Farzana Zaman, Farzana Zaman, Asma Aziz, Sayeed-Bin Faisal, Magali Traskine, MD Ahsan Habib, Javier Ruiz-Guiñazú, Dorota Borysputri dewiОценок пока нет
- 1 s2.0 S0264410X17317875 Main PDFДокумент9 страниц1 s2.0 S0264410X17317875 Main PDFputri dewiОценок пока нет
- A One-Time Screening of Newcastle Disease Hi Antibodies Before Slaughter As An Indicator of Vaccine Performance in Broiler ChickensДокумент5 страницA One-Time Screening of Newcastle Disease Hi Antibodies Before Slaughter As An Indicator of Vaccine Performance in Broiler ChickensLin DanОценок пока нет
- Q1. What Is Vaccine?: Vaccinology Lecture NotesДокумент31 страницаQ1. What Is Vaccine?: Vaccinology Lecture Notesحوراء موجد هادي غاويОценок пока нет
- Article No14Документ6 страницArticle No14rubén_sánchez_28Оценок пока нет
- 02 ImmunizationДокумент4 страницы02 ImmunizationIsha BhattОценок пока нет
- Shibuya, Tamiya Et. Al. (2021)Документ7 страницShibuya, Tamiya Et. Al. (2021)Sol ArusshaОценок пока нет
- Control of Coccidiosis in Chickens by VaccinationДокумент8 страницControl of Coccidiosis in Chickens by VaccinationAndres ForeroОценок пока нет
- The Bovine DiseaseДокумент16 страницThe Bovine DiseaseNimai Nitai DasОценок пока нет
- Jordan2017 Infectious BronchitisДокумент28 страницJordan2017 Infectious BronchitisVicente Vega SanchezОценок пока нет
- Comparative Mycoplasma Hyopneumoniae Vaccine Efficacy (Makhanon)Документ2 страницыComparative Mycoplasma Hyopneumoniae Vaccine Efficacy (Makhanon)nick224Оценок пока нет
- Oie 2018 Avian Influenza (Ai)Документ23 страницыOie 2018 Avian Influenza (Ai)nasrullahОценок пока нет
- Jiy103 - A Universal Influenza Vaccine The Strategic Plan For TheДокумент8 страницJiy103 - A Universal Influenza Vaccine The Strategic Plan For ThePGIndikaОценок пока нет
- Co-Administration of Avian Influenza Virus H5 Plasmid DNA With Chicken IL-15 and IL-18 Enhanced Chickens Immune ResponsesДокумент14 страницCo-Administration of Avian Influenza Virus H5 Plasmid DNA With Chicken IL-15 and IL-18 Enhanced Chickens Immune Responsesvirtualsagar4maxОценок пока нет
- Etiologyipi131983 PDFДокумент11 страницEtiologyipi131983 PDFAndi Dytha Pramitha Sam100% (1)
- Antibody-Dependent Enhancement (ADE), Sometimes LessДокумент12 страницAntibody-Dependent Enhancement (ADE), Sometimes LesssumantonoОценок пока нет
- Probing Into The Edible Vaccines: Newer Paradigms, Scope and RelevanceДокумент13 страницProbing Into The Edible Vaccines: Newer Paradigms, Scope and Relevanceنوره نورОценок пока нет
- 283 541 1 SM PDFДокумент5 страниц283 541 1 SM PDFarum shofyaОценок пока нет
- 10 1111@sji 12937Документ34 страницы10 1111@sji 12937Excelsis VallejoОценок пока нет
- GIUDICE 2018 - Correlates of Adjuvanticity A Review On Adjuvants in Licensed VaccinesДокумент8 страницGIUDICE 2018 - Correlates of Adjuvanticity A Review On Adjuvants in Licensed VaccinesJoão Pedro NunesОценок пока нет
- 2 Review Diphtheria Results April2017 Final Clean PDFДокумент20 страниц2 Review Diphtheria Results April2017 Final Clean PDFchameleonОценок пока нет
- Fase 3.1Документ9 страницFase 3.1Verliatesya TugasОценок пока нет
- Vaccination Against Coccidial Parasites. The Method of Choice?Документ10 страницVaccination Against Coccidial Parasites. The Method of Choice?Gopi MarappanОценок пока нет
- CHN Immunization NotesДокумент9 страницCHN Immunization NotesJustin AncogОценок пока нет
- ABM For COVID 19Документ12 страницABM For COVID 19Fuma AjiОценок пока нет
- Title PagesДокумент2 страницыTitle PagesKhalid FarooqОценок пока нет
- HI Titres 0 - 35 p-66Документ1 страницаHI Titres 0 - 35 p-66Khalid FarooqОценок пока нет
- ANOVA p68Документ1 страницаANOVA p68Khalid FarooqОценок пока нет
- Table 1p 8,9Документ2 страницыTable 1p 8,9Khalid FarooqОценок пока нет
- New RefrencesДокумент19 страницNew RefrencesKhalid Farooq100% (1)
- AGPT Titre 0 - 35 p-67Документ1 страницаAGPT Titre 0 - 35 p-67Khalid FarooqОценок пока нет
- Results Complete p48-65Документ18 страницResults Complete p48-65Khalid FarooqОценок пока нет
- Review of Literature SequencedДокумент33 страницыReview of Literature SequencedKhalid FarooqОценок пока нет
- Introduction NewДокумент5 страницIntroduction NewKhalid FarooqОценок пока нет
- Material and MethodsДокумент6 страницMaterial and MethodsKhalid FarooqОценок пока нет
- Avian Influenza (AI) Has Been Recognized As A Highly ContagiousДокумент1 страницаAvian Influenza (AI) Has Been Recognized As A Highly ContagiousKhalid FarooqОценок пока нет
- List of AbbreviationsДокумент2 страницыList of AbbreviationsKhalid FarooqОценок пока нет
- Green Life Cycle Science Butterfly Graphic OrganizerДокумент1 страницаGreen Life Cycle Science Butterfly Graphic OrganizerAlexia SalvadorОценок пока нет
- Week 2 - Text 1. Ecosystems and Human InterferencesДокумент2 страницыWeek 2 - Text 1. Ecosystems and Human Interferences458 269 Cinta Alivia AthaОценок пока нет
- Focus Smart Science M2 - AK (WB)Документ9 страницFocus Smart Science M2 - AK (WB)2422 ทัศน์พล เพ็ญสังข์Оценок пока нет
- Position Paper Genetic EngineeringДокумент6 страницPosition Paper Genetic EngineeringAJ Lemuel Carolino100% (2)
- How Do We As Nuwaupians View HomosexualityДокумент15 страницHow Do We As Nuwaupians View HomosexualityNathaniel Adams100% (1)
- Phylogenetic Trees Activity - Farizah AliДокумент2 страницыPhylogenetic Trees Activity - Farizah AliFarizah AliОценок пока нет
- Biology Feynman TechniquesДокумент9 страницBiology Feynman TechniquesmuthulaxmiОценок пока нет
- A Method For Turbidimetric Measurement of BacterialДокумент4 страницыA Method For Turbidimetric Measurement of BacterialCrisMayaОценок пока нет
- Photosynthesis AnswersДокумент1 страницаPhotosynthesis AnswersChristian Hovsep KaradaghlianОценок пока нет
- Neural TubeДокумент4 страницыNeural TubeRuth Danielle GasconОценок пока нет
- Retinoid Receptors and Therapeutic Applications of RAR/RXR ModulatorsДокумент23 страницыRetinoid Receptors and Therapeutic Applications of RAR/RXR ModulatorsAKellyNMОценок пока нет
- Stool AnalysisДокумент8 страницStool AnalysisAbed AbusalemОценок пока нет
- Lecture D111L Week 05 S13Документ64 страницыLecture D111L Week 05 S13Tahir RizviОценок пока нет
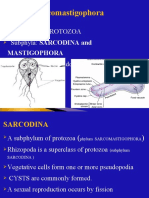
- Sarcomastigophora: A Phylum of PROTOZOA Subphyla: SARCODINA and Have Flagella or PseudopodiaДокумент20 страницSarcomastigophora: A Phylum of PROTOZOA Subphyla: SARCODINA and Have Flagella or Pseudopodiaabel semuОценок пока нет
- Year 10 DPP Biology Term 3 Exam RevisionДокумент8 страницYear 10 DPP Biology Term 3 Exam RevisionPoojit VenugopalaОценок пока нет
- MS MLPA Protocol One Tube MSP v008Документ19 страницMS MLPA Protocol One Tube MSP v008Sp PpvОценок пока нет
- CV Aeman 2018 NewДокумент4 страницыCV Aeman 2018 NewAeman KhalidОценок пока нет
- Pneumacult™-Ali: For Bronchial Epithelial CellsДокумент2 страницыPneumacult™-Ali: For Bronchial Epithelial CellsDunkMeОценок пока нет
- Orange Blue Shop Creative Trifold BrochureДокумент1 страницаOrange Blue Shop Creative Trifold BrochureBeverly PantohanОценок пока нет
- Role of Free Radicals in Health and DiseaseДокумент49 страницRole of Free Radicals in Health and DiseaseKuzhandai VeluОценок пока нет
- Algae Anatomy Biochemistry 0Документ17 страницAlgae Anatomy Biochemistry 0Nguyen Hien QuanОценок пока нет
- Bacteriophage Replication ProcessДокумент3 страницыBacteriophage Replication ProcessNicole JulianОценок пока нет
- Nonmalignant Leukocyte Disorder Chapter 26-35Документ15 страницNonmalignant Leukocyte Disorder Chapter 26-35Reizel GaasОценок пока нет
- Plant Symbionts InteractionДокумент13 страницPlant Symbionts InteractionIram TahirОценок пока нет
- Nutrition Answers FinalДокумент9 страницNutrition Answers FinalelizabethОценок пока нет
- Student Exploration: Building DNAДокумент4 страницыStudent Exploration: Building DNAShiper 33Оценок пока нет
- Fermentation FundamentalsДокумент5 страницFermentation FundamentalsWade ColemanОценок пока нет
- Data Based Question: Eaten by HerbivoresДокумент3 страницыData Based Question: Eaten by HerbivoresSulochana Devi PalanisamyОценок пока нет
- Animal Transport SystemДокумент24 страницыAnimal Transport SystemCamille TolentinoОценок пока нет
- PlanktonДокумент12 страницPlanktonRen ObeñaОценок пока нет
- Summary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedОт EverandSummary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedРейтинг: 5 из 5 звезд5/5 (80)
- The Obesity Code: Unlocking the Secrets of Weight LossОт EverandThe Obesity Code: Unlocking the Secrets of Weight LossРейтинг: 4 из 5 звезд4/5 (6)
- ADHD is Awesome: A Guide to (Mostly) Thriving with ADHDОт EverandADHD is Awesome: A Guide to (Mostly) Thriving with ADHDРейтинг: 5 из 5 звезд5/5 (1)
- Think This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeОт EverandThink This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeРейтинг: 2 из 5 звезд2/5 (1)
- The Age of Magical Overthinking: Notes on Modern IrrationalityОт EverandThe Age of Magical Overthinking: Notes on Modern IrrationalityРейтинг: 4 из 5 звезд4/5 (24)
- LIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionОт EverandLIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionРейтинг: 4 из 5 звезд4/5 (404)
- Summary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisОт EverandSummary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisРейтинг: 4.5 из 5 звезд4.5/5 (42)
- Raising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsОт EverandRaising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsРейтинг: 5 из 5 звезд5/5 (1)
- The Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaОт EverandThe Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaРейтинг: 4.5 из 5 звезд4.5/5 (266)
- Outlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisОт EverandOutlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisРейтинг: 4 из 5 звезд4/5 (1)
- By the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsОт EverandBy the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsОценок пока нет
- Why We Die: The New Science of Aging and the Quest for ImmortalityОт EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityРейтинг: 4 из 5 звезд4/5 (3)
- Cult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryОт EverandCult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryРейтинг: 4 из 5 звезд4/5 (44)
- Dark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.От EverandDark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.Рейтинг: 4.5 из 5 звезд4.5/5 (110)
- Gut: the new and revised Sunday Times bestsellerОт EverandGut: the new and revised Sunday Times bestsellerРейтинг: 4 из 5 звезд4/5 (392)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisОт EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisРейтинг: 3.5 из 5 звезд3.5/5 (2)
- The Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsОт EverandThe Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsРейтинг: 3.5 из 5 звезд3.5/5 (3)
- The Marshmallow Test: Mastering Self-ControlОт EverandThe Marshmallow Test: Mastering Self-ControlРейтинг: 4.5 из 5 звезд4.5/5 (58)
- Dark Psychology: Learn To Influence Anyone Using Mind Control, Manipulation And Deception With Secret Techniques Of Dark Persuasion, Undetected Mind Control, Mind Games, Hypnotism And BrainwashingОт EverandDark Psychology: Learn To Influence Anyone Using Mind Control, Manipulation And Deception With Secret Techniques Of Dark Persuasion, Undetected Mind Control, Mind Games, Hypnotism And BrainwashingРейтинг: 4 из 5 звезд4/5 (1138)
- Sleep Stories for Adults: Overcome Insomnia and Find a Peaceful AwakeningОт EverandSleep Stories for Adults: Overcome Insomnia and Find a Peaceful AwakeningРейтинг: 4 из 5 звезд4/5 (3)
- Raising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsОт EverandRaising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsРейтинг: 4.5 из 5 звезд4.5/5 (169)
- Mindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessОт EverandMindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessРейтинг: 4.5 из 5 звезд4.5/5 (328)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsОт EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsРейтинг: 4.5 из 5 звезд4.5/5 (6)