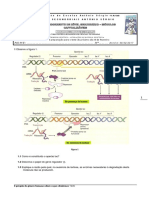
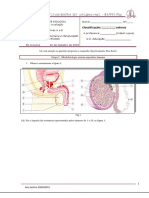
Вам также может понравиться
- 090 Ficha de Revisao - 20 3 2017Документ5 страниц090 Ficha de Revisao - 20 3 2017Ana JordãoОценок пока нет
- 071 Ficha de Revisc3b5es1Документ6 страниц071 Ficha de Revisc3b5es1Ana JordãoОценок пока нет
- Epigenética e Memória Celular Marcelo FantappieДокумент5 страницEpigenética e Memória Celular Marcelo FantappieLuiz Antonio MottaОценок пока нет
- Vacinas Anti Covid 19Документ9 страницVacinas Anti Covid 19Ana JordãoОценок пока нет
- Aspectos Comparativos Entre Os Ciclos de Vida de Diferentes PlantasДокумент3 страницыAspectos Comparativos Entre Os Ciclos de Vida de Diferentes PlantasAna JordãoОценок пока нет
- 072 Teste Escrito - 13 02 2017Документ6 страниц072 Teste Escrito - 13 02 2017Romeu VieiraОценок пока нет
- A Estranha Razão Das Listas Das ZebrasДокумент2 страницыA Estranha Razão Das Listas Das ZebrasAna JordãoОценок пока нет
- Almôndegas Com CarilДокумент1 страницаAlmôndegas Com CarilAna JordãoОценок пока нет
- Almôndegas Com CarilДокумент1 страницаAlmôndegas Com CarilAna JordãoОценок пока нет
- 1 Teste vf1 PDFДокумент9 страниц1 Teste vf1 PDFBárbara Lage GarciaОценок пока нет
- 7 Alongamentos em 7 Minutos para Alívio Completo Da Dor Nas CostasДокумент3 страницы7 Alongamentos em 7 Minutos para Alívio Completo Da Dor Nas CostasAna JordãoОценок пока нет
- 056-Ficha Revisao Dez 2016Документ5 страниц056-Ficha Revisao Dez 2016Ana JordãoОценок пока нет
- Actividade 6Документ2 страницыActividade 6Ana JordãoОценок пока нет
- Alterações ClimáticasДокумент4 страницыAlterações ClimáticasAna JordãoОценок пока нет
- Microsoft Word - Protocolo Experimental - FAZER QUEIJOДокумент1 страницаMicrosoft Word - Protocolo Experimental - FAZER QUEIJOAna JordãoОценок пока нет
- ConveccaoДокумент1 страницаConveccaoAna JordãoОценок пока нет
- Atividade Laboratorial - Subistemas - Proposta de SoluçãoДокумент1 страницаAtividade Laboratorial - Subistemas - Proposta de SoluçãoAna JordãoОценок пока нет
- Acidez Oceanos v3Документ4 страницыAcidez Oceanos v3Ana JordãoОценок пока нет
- Acidez Oceanos v3Документ4 страницыAcidez Oceanos v3Ana JordãoОценок пока нет
- AN FolhaCalculoДокумент4 страницыAN FolhaCalculoAna JordãoОценок пока нет
- Apoio Ao Estudo - Apoio ExameДокумент2 страницыApoio Ao Estudo - Apoio ExameAna JordãoОценок пока нет
- Centro Recursos Cienciaviva Fazer Sabonete 14666265755d95Документ7 страницCentro Recursos Cienciaviva Fazer Sabonete 14666265755d95Ana JordãoОценок пока нет
- A Pílula Masculina Está Cada Vez Mais PróximaДокумент4 страницыA Pílula Masculina Está Cada Vez Mais PróximaAna JordãoОценок пока нет
- Acidez Oceanos v3Документ4 страницыAcidez Oceanos v3Ana JordãoОценок пока нет
- Parecer Acerca Da Prova de Exame Nacional Do Ensino SecundárioДокумент2 страницыParecer Acerca Da Prova de Exame Nacional Do Ensino SecundárioAna JordãoОценок пока нет
- Esclerose Lateral Amiotrófica (ELA)Документ11 страницEsclerose Lateral Amiotrófica (ELA)Ana JordãoОценок пока нет
- As Picadas e Mordidas Mais Perigosas Do Mundo AnimalДокумент1 страницаAs Picadas e Mordidas Mais Perigosas Do Mundo AnimalAna JordãoОценок пока нет
- Carlos Marques Da Silva - Fósseis Ao Virar Da EsquinaДокумент5 страницCarlos Marques Da Silva - Fósseis Ao Virar Da EsquinaAna JordãoОценок пока нет
- Investigadores Que Ganharam Prémio Champalimaud Curam Cegueira HereditáriaДокумент24 страницыInvestigadores Que Ganharam Prémio Champalimaud Curam Cegueira HereditáriaAna JordãoОценок пока нет
- Boletim 5Документ5 страницBoletim 5Ana JordãoОценок пока нет
- Sistema Circulatório Excretor Respiratorio e ReprodutorДокумент20 страницSistema Circulatório Excretor Respiratorio e ReprodutorSaraiva JuniorОценок пока нет
- Gestao GemelarДокумент50 страницGestao Gemelarnatanael sousaОценок пока нет
- Fichamento ALIMENTAÇÃO NOS PRIMEIROS TRÊS MESES DE VIDA 28 - 07Документ3 страницыFichamento ALIMENTAÇÃO NOS PRIMEIROS TRÊS MESES DE VIDA 28 - 07Danielli PiresОценок пока нет
- INFLAMAÇÃOДокумент8 страницINFLAMAÇÃOBernardo Furtado CruzeiroОценок пока нет
- Epidemiologia Estrategia ConcursoДокумент50 страницEpidemiologia Estrategia ConcursoNagla Maria Bezerra MarquesОценок пока нет
- Epidemiologia - Vigilância EpidemiológicaДокумент13 страницEpidemiologia - Vigilância Epidemiológicadom padroОценок пока нет
- Acupuntura Estética Os Três Pontos Da Obesidade para o EmagrecimentoДокумент4 страницыAcupuntura Estética Os Três Pontos Da Obesidade para o EmagrecimentoRoseli De Oliveira SilvaОценок пока нет
- Prã - Projeto TCCДокумент8 страницPrã - Projeto TCCgabrel fariaОценок пока нет
- Pereira Kenp DR BotДокумент76 страницPereira Kenp DR BotGiovani AngeloОценок пока нет
- Clínica Médica Sistema CardioДокумент16 страницClínica Médica Sistema CardioAdriana NunesОценок пока нет
- Depressão Pós-PartoДокумент7 страницDepressão Pós-PartoCida NicolauОценок пока нет
- Bases Patológicas Das Doenças Do Trato GastrointestinalДокумент80 страницBases Patológicas Das Doenças Do Trato GastrointestinalRafael Trindade Burtet100% (1)
- UFCD 9137 - Anatomia Do Sistema Veno-LinfáticoДокумент35 страницUFCD 9137 - Anatomia Do Sistema Veno-LinfáticoFrederico Dinis Rocha100% (1)
- Uti SilДокумент17 страницUti SilOrion TechОценок пока нет
- Caso Clínico DiabetesДокумент2 страницыCaso Clínico DiabetesBrenno AmorimОценок пока нет
- Massagem DesportivaДокумент35 страницMassagem DesportivaClara ZilligОценок пока нет
- Nefrologia Resumo Litiase Renal TSRS 20160323Документ8 страницNefrologia Resumo Litiase Renal TSRS 20160323Danielle Do NascimentoОценок пока нет
- @biomedicina - BR - Fisiologia Aplicada A Reprodução HumanaДокумент68 страниц@biomedicina - BR - Fisiologia Aplicada A Reprodução HumanaBrena SilvaОценок пока нет
- Protocolo Sepse Sirio LibanesДокумент14 страницProtocolo Sepse Sirio LibanesNina NuzzoОценок пока нет
- MAMOGRAFIAДокумент18 страницMAMOGRAFIAAnynhha Mathos100% (1)
- Cuidados No Pós-PartoДокумент38 страницCuidados No Pós-PartoInês CachaçoОценок пока нет
- Relação Estrutura-AtividadeДокумент43 страницыRelação Estrutura-AtividadePedroОценок пока нет
- Laser Tera PiaДокумент110 страницLaser Tera Piadentistasleao100% (1)
- Atividade 2 - Saúde - Anatomia Humana - 54-2023Документ3 страницыAtividade 2 - Saúde - Anatomia Humana - 54-2023Cavalini Assessoria Acadêmica100% (1)
- Revista 12 Maio Junho 2012Документ68 страницRevista 12 Maio Junho 2012Croitort53Оценок пока нет
- Distúrbios CirculatóriosДокумент29 страницDistúrbios CirculatórioslevivetОценок пока нет
- Anamnese de AdultoДокумент5 страницAnamnese de AdultoArlan Pinto100% (1)
- Avancos em Periodontia e Implantodontia - Paradigmas e Desafios - Cap 03Документ18 страницAvancos em Periodontia e Implantodontia - Paradigmas e Desafios - Cap 03francescogregorioОценок пока нет
- Cuidados Na GestaçãoДокумент4 страницыCuidados Na Gestaçãoisabellacassi.opsОценок пока нет
- Reações de HipersensibilidadeДокумент19 страницReações de Hipersensibilidadeamandaterra99Оценок пока нет