Вам также может понравиться
- Paradigmas en EspañolДокумент16 страницParadigmas en EspañolLuifer AgudeloОценок пока нет
- AMPC, Insulina, Glucogeno y Fosfoinosintoles. Cascadas de SeñalisaciónДокумент3 страницыAMPC, Insulina, Glucogeno y Fosfoinosintoles. Cascadas de SeñalisaciónMilton Alexi Sepulveda Castro100% (1)
- B-Arrestin Mediated Receptor Trafficking and Signal TraductionДокумент13 страницB-Arrestin Mediated Receptor Trafficking and Signal TraductionSILVIA FERNANDA CASTILLO GOYENECHEОценок пока нет
- Receptores Acoplados A Proteína G y Segundos MensajerosДокумент6 страницReceptores Acoplados A Proteína G y Segundos MensajerosOriana CoОценок пока нет
- Cascadas de SeñalizaciónДокумент2 страницыCascadas de SeñalizaciónAaron CasañasОценок пока нет
- TransferasasДокумент2 страницыTransferasasRenata OdalОценок пока нет
- Estructura e Interacciones Biologicas de Heparina y Heparán SulfatoДокумент12 страницEstructura e Interacciones Biologicas de Heparina y Heparán SulfatoJosé Alberto S GОценок пока нет
- 3regulacion Coordinada Glucolisis-GluconeogenesisДокумент9 страниц3regulacion Coordinada Glucolisis-GluconeogenesisSanti GarciaОценок пока нет
- Tema 3Документ6 страницTema 3Xisco Lozano SalvaОценок пока нет
- Glucolisis y Ciclo de KrebsДокумент2 страницыGlucolisis y Ciclo de KrebsAgustinaОценок пока нет
- Ajuste InducidoДокумент7 страницAjuste InducidoFer AmezagaОценок пока нет
- Gaba Casi CompletoДокумент3 страницыGaba Casi CompletoRocio Méndez FrancoОценок пока нет
- Receptores de MembranaДокумент17 страницReceptores de MembranaUriel YépezОценок пока нет
- VIA WNT ExpliДокумент1 страницаVIA WNT ExpliCarlos SotoОценок пока нет
- Regulacion Del Ciclo de KrebsДокумент5 страницRegulacion Del Ciclo de KrebsDrigo S NavarroОценок пока нет
- Regulación de La Actividad Enzimática Alosterica y Covalente en GlucolisisДокумент5 страницRegulación de La Actividad Enzimática Alosterica y Covalente en GlucolisisAnyi ColmenarezОценок пока нет
- El AMP CíclicoДокумент6 страницEl AMP CíclicoCeci MartinezОценок пока нет
- LisinaДокумент4 страницыLisinaVALERIA TORRES VIÑUELAОценок пока нет
- Curva Dosis Respuesta y Accion Combinada de FXДокумент13 страницCurva Dosis Respuesta y Accion Combinada de FXAnette VillaОценок пока нет
- Receptores de MembranaДокумент9 страницReceptores de MembranaAlexy SanchezОценок пока нет
- Temario Biomol 3er ExmДокумент6 страницTemario Biomol 3er ExmMARCO ANTONIO SOSA AGUILARОценок пока нет
- Carboxipeptidasa AДокумент3 страницыCarboxipeptidasa ACristian CCОценок пока нет
- Beta AntagonistaДокумент3 страницыBeta AntagonistajoseОценок пока нет
- Procesos Oxidativos 1Документ71 страницаProcesos Oxidativos 1alexander naspiranОценок пока нет
- Dudas BiomolДокумент3 страницыDudas BiomolLoreto GuazoОценок пока нет
- La Actina PDFДокумент5 страницLa Actina PDFItzaetlSantanaM0% (1)
- Transducción de La Señal Por Receptores Unidos A Proteína GДокумент3 страницыTransducción de La Señal Por Receptores Unidos A Proteína GClaudia Gonzalez RuizОценок пока нет
- TRADUCCIONДокумент2 страницыTRADUCCIONAngie Nathalia MartinezОценок пока нет
- Solución Caso 7Документ6 страницSolución Caso 7Edisson Abdiel Rodriguez TorresОценок пока нет
- 04.AB Discusion y ConclusionesДокумент29 страниц04.AB Discusion y ConclusionesAline Sahad Hernández HernándezОценок пока нет
- 2 - Transducción de SeñalesДокумент5 страниц2 - Transducción de SeñalesHernán GuietОценок пока нет
- TEMA 7 "Ciclo de Los Ácidos Tricarboxílicos (TCA) "Документ9 страницTEMA 7 "Ciclo de Los Ácidos Tricarboxílicos (TCA) "Ana Maria Jaen GarciaОценок пока нет
- AMPc ExposicionДокумент5 страницAMPc ExposicionLaura Sofia Rintha JaraОценок пока нет
- Ciclo de Krebs, Transporte de Electrones, Fosforilación OxidativaДокумент18 страницCiclo de Krebs, Transporte de Electrones, Fosforilación OxidativasebastianroseroОценок пока нет
- Maduración PostranscripcionalДокумент15 страницMaduración Postranscripcional431732267% (3)
- Temario Tercer Parcial de BiomolДокумент7 страницTemario Tercer Parcial de BiomolMARCO ANTONIO SOSA AGUILARОценок пока нет
- FINAL BIOQUÍMICA 13/12 TEMA A y BДокумент6 страницFINAL BIOQUÍMICA 13/12 TEMA A y BKristoff DamianОценок пока нет
- PartessДокумент4 страницыPartessjahuer steven rueda tibaguizaОценок пока нет
- MAP KinasasДокумент1 страницаMAP KinasasLuzaren RiveraОценок пока нет
- Clase 11 - Enzimas ReguladorasДокумент81 страницаClase 11 - Enzimas ReguladorasJaime Saavedra Guevara100% (3)
- Apuntes Proteinas GДокумент40 страницApuntes Proteinas GYulsAelbert ReRoОценок пока нет
- Metabolismo Glucogeno ResumenДокумент8 страницMetabolismo Glucogeno Resumen210220Оценок пока нет
- Ciclo de KrebsДокумент10 страницCiclo de KrebsMariana crystel Cabanillas SalcidoОценок пока нет
- Taller 7b Proteínas 2022 ResoluciónДокумент6 страницTaller 7b Proteínas 2022 Resoluciónjuanita mendezОценок пока нет
- 2022 - P2 - TEMA B RespuestasДокумент2 страницы2022 - P2 - TEMA B RespuestasvaalenmediОценок пока нет
- Seminario de Bioquimica 1Документ7 страницSeminario de Bioquimica 1Teddy luisОценок пока нет
- Proteina GДокумент6 страницProteina GThalina PayaresОценок пока нет
- Pregunta 2 Mecanismo de Accion de La Insulina e HipoglucemiantesДокумент6 страницPregunta 2 Mecanismo de Accion de La Insulina e HipoglucemiantesJorge GonzalesОценок пока нет
- Final Bioquimica FMED UBA 14-2-19 TEMA AДокумент2 страницыFinal Bioquimica FMED UBA 14-2-19 TEMA ACande AmiconeОценок пока нет
- Proteina GДокумент6 страницProteina GAngela Gonzales HerreraОценок пока нет
- DevlinДокумент15 страницDevlinFacundo Ezequiel CorralesОценок пока нет
- Los Peptidos Cortos Se Auto-Ensamblan para Producir Amiloides CataliticosДокумент8 страницLos Peptidos Cortos Se Auto-Ensamblan para Producir Amiloides CataliticosJuan BolañoОценок пока нет
- Tarea Final Rutas MetabolicasДокумент14 страницTarea Final Rutas MetabolicasÁngelОценок пока нет
- Señalización de Proteínas G y AMP CiclicoДокумент7 страницSeñalización de Proteínas G y AMP CiclicoAmílkar Ocegueda MaderaОценок пока нет
- Cascadas de SeñalizaciónДокумент2 страницыCascadas de SeñalizaciónLuna LolsitoОценок пока нет
- Cerp Centro Julio 2020 1era Prueba Parcial Bioquimica y Farmacologia - NadiagomezДокумент5 страницCerp Centro Julio 2020 1era Prueba Parcial Bioquimica y Farmacologia - NadiagomezTatiana GómezОценок пока нет
- Proteinas GДокумент3 страницыProteinas Gcorrales.gersonОценок пока нет
- Ciclo de KrebsДокумент9 страницCiclo de KrebsMileОценок пока нет
- Informe Repuesto Ecografo Aviso 2018Документ2 страницыInforme Repuesto Ecografo Aviso 2018Walter RamirezОценок пока нет
- Anemia 1 4 5Документ31 страницаAnemia 1 4 5Walter RamirezОценок пока нет
- Fisio CirculatorioДокумент12 страницFisio CirculatorioWalter RamirezОценок пока нет
- Inf Braun Emergencia Octubre 2018Документ1 страницаInf Braun Emergencia Octubre 2018Walter RamirezОценок пока нет
- MaterialesДокумент21 страницаMaterialesWalter RamirezОценок пока нет
- TDR Sillones Dentales Osce-Essalud 2019Документ9 страницTDR Sillones Dentales Osce-Essalud 2019Walter RamirezОценок пока нет
- TDR - Servicio de Mantenimiento IteДокумент3 страницыTDR - Servicio de Mantenimiento IteWalter RamirezОценок пока нет
- FangoterapaДокумент29 страницFangoterapaWalter RamirezОценок пока нет
- TDR - Servicio de Mantenimiento IteДокумент3 страницыTDR - Servicio de Mantenimiento IteWalter RamirezОценок пока нет
- Biografía de Jorge BasadreДокумент6 страницBiografía de Jorge BasadreWalter RamirezОценок пока нет
- EVALUACIÓN 1ER QUIMESTRE Biología 2do.Документ4 страницыEVALUACIÓN 1ER QUIMESTRE Biología 2do.Diana Galiano100% (1)
- Presentación Ácidos NucleicosДокумент31 страницаPresentación Ácidos NucleicosLucy BuitronОценок пока нет
- Genetica Problemas PDFДокумент86 страницGenetica Problemas PDFNina Romero RicciОценок пока нет
- Organelos Endomembranosos y No MembranososДокумент52 страницыOrganelos Endomembranosos y No MembranososmilenkaОценок пока нет
- 8basico - Evaluacion Ciencias CelulaДокумент8 страниц8basico - Evaluacion Ciencias CelulaGerith MuñozОценок пока нет
- Actividad de Aprendizaje Sintesis de ProteinasДокумент12 страницActividad de Aprendizaje Sintesis de ProteinasDaniela León Castillo100% (1)
- Hoja de Trabajo ADN y Síntesis de Proteinas-2-1 PDFДокумент3 страницыHoja de Trabajo ADN y Síntesis de Proteinas-2-1 PDFjupa7953Оценок пока нет
- Operones y Fenómeno de Wobble PDFДокумент2 страницыOperones y Fenómeno de Wobble PDFtania pinargoteОценок пока нет
- Fosfolipasas CДокумент10 страницFosfolipasas CRoxiiHh TNОценок пока нет
- PTM / Processing: Procesamiento de MoléculasДокумент6 страницPTM / Processing: Procesamiento de MoléculasIsabel SánchezОценок пока нет
- Guía Práctica 6 Membrana Celular Modelo de Organización. Transporte A Traves de MembranaДокумент16 страницGuía Práctica 6 Membrana Celular Modelo de Organización. Transporte A Traves de MembranaQuimica TecnologiaОценок пока нет
- Resumen de Biologia de Adn-ArnДокумент6 страницResumen de Biologia de Adn-ArnMIRKO MAURO QUISPE FARFANОценок пока нет
- Aa. y Peptidos Con Interes BiologicoДокумент16 страницAa. y Peptidos Con Interes BiologicoBeatrízLeónОценок пока нет
- ASPECTOS DE INTERÉS SOBRE LAS ACUAPORINAS PLANTAS - 2014 - Prof Jorge A Leyva R PDFДокумент10 страницASPECTOS DE INTERÉS SOBRE LAS ACUAPORINAS PLANTAS - 2014 - Prof Jorge A Leyva R PDFPaula De Alba SurmayОценок пока нет
- Clase 4 Célula y Organelos Subcelulares II 2020 PDFДокумент30 страницClase 4 Célula y Organelos Subcelulares II 2020 PDFIgnacio González VeraОценок пока нет
- Núcleo CelularДокумент5 страницNúcleo CelularMarisela TomalaОценок пока нет
- 5.wuolah Premium Apuntes T5 Sistemas de Membranas CitoplasmaticasДокумент27 страниц5.wuolah Premium Apuntes T5 Sistemas de Membranas CitoplasmaticasMARTA 036Оценок пока нет
- Extraccion de ADN en Vegetales.Документ5 страницExtraccion de ADN en Vegetales.Andro GamesОценок пока нет
- Celula VegetalДокумент9 страницCelula VegetalROSAISABEL JARAОценок пока нет
- Practica Calificada BioinformaticaДокумент2 страницыPractica Calificada BioinformaticaLuis Eduardo Strater Mc LellanОценок пока нет
- Codigo GeneticoДокумент1 страницаCodigo GeneticoBrahyan SuárezОценок пока нет
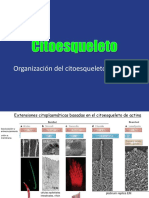
- CITOESQUELETOДокумент32 страницыCITOESQUELETOCeci SRОценок пока нет
- Guía. AMINOACIDOSДокумент6 страницGuía. AMINOACIDOSGeraldine PerezОценок пока нет
- Bandera ZuДокумент203 страницыBandera ZuSofia Chagui TatisОценок пока нет
- Complejo Mayor de Histocompatibilidad - Docbymatias-SamaДокумент12 страницComplejo Mayor de Histocompatibilidad - Docbymatias-SamaTamara Andrea Acuña CastroОценок пока нет
- Clase N 4 ActividadesДокумент2 страницыClase N 4 ActividadesDochis ToriaОценок пока нет
- Universidad Nacional de Cajamarca: Ciencias AgrariasДокумент13 страницUniversidad Nacional de Cajamarca: Ciencias AgrariasMAYRA LIZETH MORI HOYOSОценок пока нет
- 4.1 AminoacidosДокумент51 страница4.1 AminoacidosDarifori100% (1)
- Codigos LaboratorioДокумент22 страницыCodigos LaboratorioAndres Candela Mamani75% (4)
- Epigenética en Obesidad y Diabetes Tipo 2 - Papel de La Nutrición, Limitaciones y Futuras AplicacionesДокумент1 страницаEpigenética en Obesidad y Diabetes Tipo 2 - Papel de La Nutrición, Limitaciones y Futuras AplicacionesTANIA CECILIA VASQUEZ ROMEROОценок пока нет