Академический Документы
Профессиональный Документы
Культура Документы
Castillo Paterna, María Del Mar
Загружено:
Iiaii Espinoza IimiiАвторское право
Доступные форматы
Поделиться этим документом
Поделиться или встроить документ
Этот документ был вам полезен?
Это неприемлемый материал?
Пожаловаться на этот документАвторское право:
Доступные форматы
Castillo Paterna, María Del Mar
Загружено:
Iiaii Espinoza IimiiАвторское право:
Доступные форматы
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante.
Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
UNI VERSI DAD
DEALI CANTE
PROGRAMADE
DOCTORADOEN
BI OTECNOLOGI A
YBI OLOG A
EXPERI MENTAL
MODELOSDEENVEJ ECI MI ENTO
I NVI TROEI NVI VO,
ESTRSOXI DATI VOY
PROTECCI ONANTI OXI DANTE
MEMORI A
Quepara optar
al grado dedoctora
enBi ol og a
presenta
MARI ADELMARCASTI LLOPATERNA
Al i cante,
2002
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Da. Mari a Mercedes
Pal mero Cabezas,
prof esora t i t ul ar de E. U. de
Farmacol og a, yD.
J uanLorenzoBel l ot
Bernab, prof esor t i t ul ar de E. U.
de
Farmacol og a
del aUni versi dadde
Al i cant e.
CERTI FI CAN:
Que el t rabaj o
experi ment al
correspondi ent e al a present e
memori a
t i t ul ada
"Model os de envej eci mi ent o
i n vi t ro e i n vi vo,
est rs oxi dat i vo y
prot ecci nant i oxi dant e",
hasi do real i zado por
Mari adel Mar Cast i l l o Pat erna
baj o nuest ra di recci n
y que, a
nuest ro j ui ci o, reune l as
condi ci ones
adecuadas
paraser present aday
j uzgadapor el t ri bunal
correspondi ent epara
aspi raraal gradode
doct or enBi ol og a.
WHO
Al i cant e a5
Di ci embrede 2002
Fdo. Dra. M. M. Pal mer
o Cabezas
Fdo. Dr. J . L. Bel l ot Bernab
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Este trabaj o hasi do f i nanci ado conunaayudadel Fondode
I nvesti gaci ones Sani tari as de l a Seguri dad Soci al (FI SS 99/ 1264) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Dedi cadaa
Al f redo Orts
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
AGRADECIMIENTOS
Sontantas l as
personas quehanhechoposi bl e que
este
trabaj o
haya
l l egadoa suf i n, quel o ms probabl e
sea queal i ntentar nombrarl as a todas
seguro que me ol vi de
de al gui en, as que desde este prraf o y
a todas esas
personas l es doyl as graci as.
Megustar a destacar :
Al grupo
de i nvesti gaci ndel Dr. Orts por
acogerme y hacerme senti r
comouna ms entanbreve
ti empo, yal por of recerme l a
oportuni dad de
real i zar l a tesi s
doctoral ensul aboratori o.
AMercedes Pal mero, por su l abor como
codi rectora de este trabaj o y
transmi ti rme sus conoci mi entos
sobre l a materi a, y l os buenos
ratos
comparti dos enel
l aboratori oysobretodopor su
"paci enci a" conmi esti l o de
redacci n.
AJ uanl o
Bel l ot, por sudedi caci n.
A
Cri sti na Garc a, por todos
esos buenos ratos tantodentro
comof uera
del l aboratori o, como
en
Pal ma
2000.
AMi guel
ngel Company, por
i ni ci armeyensearmetodotodo
sobre el
mundodel os cul ti vos cel ul ares
.
AMi guel Sanz, por
todos l os ratos comparti dos
enel oxi graf o, mi di endo
ATPyLDHy
trasmi ti rmesus conoci mi entos
sobrei nf ormti ca.
AAnamar a Madari aga, por
escucharme, por todos
sus buenos consej os
yl os buenos
ratos comparti endobancada.
AJ ai me Mi quel , por
su ayuda desi nteresada y sus
consej os sobre
envej eci mi ento
yanti oxi dantes.
Ami s ami gos que aunque
nol os nombre me acuerdo
de todos el l os,
graci as, yenespeci al
aAsun, InmayMa Carmenpor
todoel apoyoof reci do en
l os
momentos ms di f i ci l es.
Por l ti mo, qui ero dar l as graci as
a mi f ami l i a por todo su apoyoy
el
haber estadoah en
todomomento.
Por todo el l o, ya todos vosotros,
graci as desdel o ms prof undo de mi
corazn.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I NDI CE
I NTRODUCCI ON
1.
Radi cal es l i br es deox genoyni tr genoy
estr s oxi dati vo. . . . . . . . . .
19
1. 1. Pr i nci pal es hi tos
en
el
desar r ol l o deestos conceptos. . . . . . . . . . . . . . . . . . . . .
19
1. 2 . Def i ni ci n
ycl asi f i caci ndel os r adi cal es l i br es. . . . . . . . . . . . . . . . . . . . . . . . . . . .
21
1 . 3
.
Mecani smos def or maci ndel os r adi cal es
l i br es. . . . . . . . . . . . . . . . . . . . . . . . . . .
22
1. 4. Ox genoy
r adi cal es l i br es del ox geno. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
25
1. 5. Pr opi edades del os r adi cal es
l i br es de ox geno. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
26
1. 6. Ni tr genoy
r adi cal es l i br es de ni tr geno. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
28
1. 7.
Pr opi edades del os r adi cal es l i br es deni tr geno
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . 29
2. Si stemas dedef ensacel ul ar
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
31
2. 1. Anti oxi dantes
pr i mar i os. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
31
2. 2.
Anti oxi dantes secundar i os. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
33
2. 3. Vi tami naC. . . . . . . .
.
.
. . . . . . . .
.
. . .
.
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
34
2. 4. N- aceti l - ci ste na
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
.
. .
.
. . .
.
.
. . . . . . . . . . . . . . . . . . . . . . . . . . . 35
2. 5. ci doti azol i d n
car box l i coo ti opr ol i na. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
36
3. Dao
cel ul ar por estr s oxi dati vo . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
37
3. 1. Daoapr ote nas
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
.
.
. . .
.
.
38
3. 2 . Daoal i pi dos
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
.
. .
. . . . . . . . . . . . . . . . . . . . .
39
3
. 3. Daoal ADN. . . . . . . . . . . . . . . . . . . . . . .
.
. .
. .
.
.
.
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
40
3. 4. Daoacar bohi dr atos. . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
.
. .
. . . . . . . . . . . . . . 41
4. Apoptosi s
i nduci dapor estr s oxi dati vo
. . . . . . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . .
41
4
. 1. Regul aci ndel os mecani smos de muer te
cel ul ar. . . . . . . . . . . . . . . . . . . . . . . . . . . 42
5. Patol og as
r el aci onadas conel estr s oxi dati vo. . . . . . . . . . . . . . . . . . . . . . . . . . . . .
45
6.
El envej eci mi entobi ol gi co
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
46
6. 1. Factor es
quei nf l uyenenel envej eci mi ento. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
47
6. 2 . Teor as sobr el as causas del envej eci mi ento
nor mal . . . . . . . . . . . . . . . . . . . . . . . . 51
6. 2. 1. Teor adel
"envenenami ento" por l os r adi cal es l i br es de
ox geno. .
. .
.
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . .
51
6. 2. 2. Teor adel os r adi cal es
l i br es. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
51
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
6. 2 . 3. Teor a mi tocondri al del envej eci mi ento. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
52
6. 2 . 4. Teor a
de
l a mutaci n
genti ca. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
53
6
. 2 . 5. Rel evanci a gerontol gi ca de l os estudi os sobre tel meros y
tel omerasa. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. 54
6. 3 . Concepto i ntegrador de teor as cl si cas ymodernas . . . . . . . . . . . . . . . . . . . . . . .
56
7. Pri nci pal es enfermedades
ocul ares asoci adas conel estrs
oxi dati vo yel envej eci mi ento. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
57
7. 1 . Degeneraci n
macul ar asoci ada al a edad. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 57
8.
Model os para el estudi o del estrs oxi dati vo y
envej eci mi ento. . . .
58
8. 1 . Epi tel i o pi gmentari o de reti na
como model o de envej eci mi ento
cel ul ar i n
vi tro. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
.
.
. . . . . 58
8. 2 . RatnOF1 como model o de envej eci mi ento i n
vi vo
. . . . . . . . . . . . . . . . . . . . . . . .
59
OBJETIVOS. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. . . . .
65
MATERIAL
YMETODOS
1 . Materi al . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
69
1 . 1 .
Materi al bi ol gi co. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. . . . . .
. . .
. . . . . . . . . . . . . . . . . . . . .
69
1 . 2 . Reacti vos. . . . . . . . . . . . . .
.
. . . . . . . . . . .
.
. .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
69
1 . 3 . Aparataj e. . . .
. . . . . . . . . . .
. .
.
.
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. . . . .
70
2. Mtodos de l os estudi os i n vi tro. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
71
2 . 1 . Cul ti vo de cl ul as del epi tel i o pi gmentari o
de reti na. . . . . . . . . . . . . . . . . . . . .
71
2 . 1 . 1 .
Cul ti vos pri mari os. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. . . . . .
71
2 . 1 . 2 . Subcul ti vos de EPRb. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
73
2 . 2 . Inducci ndel dao cel ul ar por estrs
oxi dati vo. . . . . . . . . . . . . . . . . . . . . . . . . . . .
74
2 . 2 . 1
.
Inducci nde estrs oxi dati vo por agentes qu mi cos
. . . . . . . . . . .
74
2 . 2 . 2 . Inducci nde cl ul as de
EPRbenunaatmsfera conbaj a
concentraci n de ox geno. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
75
2 . 2 . 3 . Inducci n de estrs oxi dati vo por
peroxi ni tri to. . . . . . . . . . . . . . . . . . .
76
2 . 3. Tratami ento conanti oxi dantes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
76
2 . 4 . Determi naci n
del dao cel ul ar . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
76
2 . 4
. 1 .
Determi naci n de l a acti vi dad l actato
deshi drogenasa. . . . . . . .
76
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
2. 4 . 2. Determi naci ndel a concentraci ni ntracel ul ar deATP
. . . . . . 77
2
. 4
. 3.
Detecci ndel a apoptosi s. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
78
a) Ti nci nconHoechst 33258(bi s- benzami da) . . . . . . . . . . . . . . . . . . . . . . . . . .
78
b) El ectrof oresi s engel deazarosa. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
79
3. Mtodos del
model odeenvej eci mi ento i nvi vo. . . . . . . . . . . . . . . . . . . . . . . . . . .
83
3. 1 .
Ani mal es de experi mentaci n. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
83
3. 2. Estudi odeparmetros no i nvasi vos f i si ol gi cos
yconductual es. . . . .
84
3. 2. 1 . Consumopromedi o
deal i mento. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
84
3. 2. 2. Determi naci ndel peso corporal . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
84
3. 2. 3. Parmetros
deconducta. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
84
a) Medi dade parmetros devi gor ycoordi naci n
neuromuscul ar 85
b) Medi dadel a acti vi dad
expl oratori a espontnea. . . . . . . . . . . . . . . . . . . . . .
86
3. 3 . Obtenci n
demuestras de tej i dos paraestudi os
bi oqu mi cos. . . . . . . . .
88
3. 4 . Ai sl ami entodemi tocondri as
deh gadoyderi nydetermi naci n
deprote nami tocondri al . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
88
3. 5. Determi naci n
del consumodeox geno. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
91
3. 6.
Determi naci ones enzi mti cas. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
92
3. 6. 1 . Acti vi dad
ci tocromocoxi dasa. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
92
3. 6. 2. Acti vi dadci trato si ntetasa
. . . . . . . . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . .
. . . . . . . . .
93
3. 6. 3. Determi naci n
demal ondi al deh do. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
94
4.
Mtodos estad sti cos. . . . . . . . . . . . . . . . . . . . . . . . .
. . . . . . .
.
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
95
RESULTADOS
1
.
Model odeenvej eci mi ento cel ul ar i n
vi tro. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
99
1 . 1 . I nducci nde daocel ul ar por
peroxi ni tri to. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
99
1 . 2. I nducci n
deestrs oxi dati vo por agentes qu mi cos. . . . . . . . . . . . . . . . . . . . . . .
104
1 . 3 . I nducci ndedaocel ul ar por
hi poxi a. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 108
1 . 4 .
I nducci ndeapoptosi s por estrs oxi dati vo. . . .
. . . . . . .
.
. . . . . . . . . . . . . . . . . . . . . . . 110
1 . 4 . 1 . Detecci nmedi anteti nci n
conHoechst. . . . . . . . . . . . . . . . . . . . . . . . . . . . . 110
1 . 4
. 2. El ectrof oresi s engel deazarosa. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
116
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
2. Model odeenvej eci mi entoi nvi vo. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
119
2. 1. Supervi venci ayparmetros f i si ol gi cos del ratn. . . . . . . . . . . . . . . . . . . . . . . . . .
119
2. 1. 1. Supervi venci adel a pobl aci n. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
119
2. 1. 2. PesoCorporal . . . . . . . .
.
. . . . . . . . . . .
.
. .
.
. .
. . . .
. . . . . .
.
.
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . 120
2. 1. 3 .
Consumopromedi odepi enso. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 129
2. 2. Ef ectos sobreparmetros deconducta. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
130
2. 2. 1
.
Pruebadecoordi naci nneuromuscul ar yvi gor. . . . . . . . . . . . . . . . . .
130
2. 2. 2. Ef ectos sobrel aacti vi dadexpl oratori a. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
132
2. 3 . Determi naci ones
bi oqu mi cas. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
135
2. 3. 1.
Consumodeox genomi tocondri al . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
135
2. 3. 2. Acti vi dadCi tocromoc oxi dasa. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
138
2. 3 . 3 . Acti vi dadCi trato
si ntetasa. . . . . . . . . . . . . . . . . . . . . . . . . .
.
. .
. .
. . . . . . .
. . . . . . . .
141
2. 3 . 4. Determi naci n
deMal ondi al deh do. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
143
DI SCUSI N
1. Model odeenvej eci mi entocel ul ar i nvi tro. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
147
2. Model odeenvej eci mi ento
i nvi vo. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
157
CONCLUSI ONES. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . .
173
BI BLI OGRAFI A
. . .
. .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
172
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
ABREVIATURAS
-ATP: adenosi na-5' -di f osf at o
.
-BSA: al bmi na
sr i ca bovi na
-CTE: cadenade t r anspor t e de
el ect r ones.
-DMAE: degener aci n
macul ar asoci ada al a edad.
-DMEM
: medi o de Dul beco modi f i cado por Eagl e.
-e- : el ect r n.
-EPRb: epi t el i o pi gment ar i o
de
r et i na
bovi na.
-ERN:
especi es r eact i vas del ni t r geno.
-ERO: especi es r eact i vas del ox geno
.
-GSH: gl ut at i on
.
-GSSG: gl ut at i on oxi dado.
-H202: per xi do
de hi dr geno.
-LDH
: l act at o deshi dr ogenasa.
-MDA: mal ondi al deh do
.
-NAC: n-acet i l -ci st e na
.
-NAD:
ni cot i nami da adeni na
di nucl et i do.
-NADPHoxi dasa:
ni cot i nami da adeni na di nucl et i do
f osf at o oxi dasa.
-NMDA:
N-met i l -D-aspar t at o.
-NO: xi do n t r i co.
-NOS: xi do
n t r i co si nt et asa.
-NS: no super vi vi ent es.
-PBS:
t ampnf osf at o sal i no de Dul beco.
-PFA: par af or mal deh do.
-PN: per oxi ni t r i t o.
-RL:
r at ones l ent os
-RR: r at ones r pi dos.
-S: super vi vi ent es
.
-SFB
:
suer o f et al bovi no.
-TP: ci do t i azol i d n
car box l i co o t i opr ol i na.
-VC
: vi t ami na C.
-VE: vi t ami na E
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I NTRODUCCI N
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
1. Radi cal es l i bres deox genoy
ni trgeno
.
Estrs oxi dati vo
I ntroducci n
1. 1. Pri nci pal es hi tosenel desarrol l o
deestosconceptos
Actual mente,
el estudi o del os radi cal es l i bres deox genoyasoci adoel
estrs oxi dati vo
es unode l os temas bi omdi cos de mayor
i nters teri co y
prcti co. Esto se debe a
su rel aci n con l a f i si opatol og a de muchas
enf ermedades
crni cas degenerati vas y sobre
todo con el proceso del
envej eci mi ento, que debi do
al aumento de l a l ongevi dad ( y de l as
enf ermedades
l i gadas al a edad) es el probl ema
sani tari o msi mportante de
l os pa ses i ndustri al i zados.
Segnl a revi si n
de Mi quel ( 1998), "Es sorprendente
quel os radi cal es
l i bres
deri vados del ox geno, quesonl a base
del avi daaerobi a, puedanl l evar
tambi n a procesos degenerati vos como
el cncer yel envej eci mi ento". La
expl i caci ndeesta
paradoj a se encuentraenel hechodeque
el ox genopuede
causar
unagranvari edaddeef ectos txi cos
quesonespeci al mentemarcados
enel si stemanervi oso
.
Por otra parte, l a i nvesti gaci n sobre
l a toxi ci daddel ox genotambi n
estuvol i gadaal usocl ni co
deeste gas encmaras deterapi a
hi perbri cayal
descubri mi ento
de que concentraci ones
al tas de ox geno aumentan l a
sensi bi l i dad de l as cl ul as
a l a radi aci n i oni zante ( Grayy
Scott, 1964),
abri endo as el
cami noanumerosos estudi os sobreel
denomi nado"ef ectodel
ox geno.
Estos estudi os conf i rmaron
hi ptesi s anteri ores sobrel a
i mportanci ade
l a acti vaci n del
ox geno, es deci r apoyaron
el concepto de que tanto l os
ef ectos benef i ci osos como
l os ef ectos noci vos de este gas se deben a l a
f ormaci n de
radi cal es l i bres de ox geno a travs
de uncambi o en su
conf i guraci nel ectrni caquel l eva
al af ormaci nderadi cal es l i bres ( Mul l i ken,
1929) . Yestos
mecani smos se termi narondeacl arar
graci as al a hi ptesi s de
Mi chel i s ( 1949) quel a mol cul adeox geno
sereduce gradual mente, es deci r
ganael ectrones deunoenuno,
por l o cual sef ormanf ragmentos mol ecul ares
conunel ectrnnoapareado
yconunamuyal tareacti vi dad, que
danl ugar a
reacci ones noprogramadas.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I ntroducci n
Esta parti cul ari dad de l a qu mi ca
del
ox geno
ti ene gran i mportanci a
bi omdi ca, comoresumeGutteri dge (1986) : "Laventaj ade este procesoparal a
vi da aerobi aes unaconsi derabl e
desacel eraci n de l as reacci ones del ox geno
con compuestos no- radi cal es. El i nconveni ente es que l a reducci n del
02
de
el ectrnenel ectrnproduceespeci es deox genoreacti vas (ERO)" .
Unode l os ef ectos patol gi cos de estas EROquerpi damente
despert
i nters f ue l a peroxi daci n
de l pi dos, que ocurre en el tej i do adi poso de
ani mal es def i ci entes en l a vi tami na anti oxi dante E (DamyGranados, 1945),
conj untamente se observuna
acumul aci nde perxi dos enel tej i do adi poso
al rededor de l as aortas ateromatosas (Gl avi nd et al . , 1952)
.
Por otra
parte, l a
observaci n de perxi dos de l pi dos
que se f orman en l a pi el expuesta a l os
rayos Xo l uz ul travi ol eta l l ev aDuboul oz yDumas (1954, 1955) aproponer
quedi chos perxi dos estnl i gados aunprocesode i nf l amaci nti sul ar.
Otro dato f undamental para el desarrol l o
de l a bi omedi ci na del as ERO
es
que
l as
prote nas que transportan hi erro (hemogl obi na yotros pi gmentos
hemti cos), as como l as mi tocondri as, ti enen un papel cl ave
en el estrs
oxi dati voquel l eva al a f ormaci n
del i poperxi dos (Tappel et al . , 1961; Tappel ,
1965) . Segncomentaremos ms detal l adamente en otras secci ones
de esta
memori adeTesi s, l as mi tocondri as
nosl o producenEROsi noqueestnmuy
expuestas al resul tante estrs oxi dati vo, pues aproxi madamente
el 90%del
ox geno que usan l os mam f eros se procesa en l a
cadena respi ratori a de l a
membranami tocondri al i nterna.
Al a vez
que se i nvesti gaban l os ef ectos bi ol gi cos ypatol gi cos de
l as
EROse f ue avanzando en l a comprensi nde l os
mecani smos de protecci n
contra el estrs oxi dati vo basados
enl a acci nde l os pri nci pal es compuestos
de acci n anti oxi dante o barredora de radi cal es l i bres, como l a vi tami na
E
(Evans yBi shop, 1922), l avi tami na C(Szent- Gi orgyi ,
1928) yel gl utati on (Vi a,
1990) . As tambi ntuvi eron unai mportanci a f undamental
el descubri mi ento
de l os peroxi somas; orgnul os que conti enen
catal asa ydetoxi f i can perxi dos
(De Duve,
1969) j untoconl a i denti f i caci n por McCordyFri dovi ch (1969)
de
l as superxi do
di smutasas, enzi mas que el i mi nan el radi cal
superxi do
i mpi di endoas suacci npatol gi ca.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I ntroducci n
En base a una extensa revi si n sobre estos temas, concl u a Pryor
( 1982) : "No hace mucho
ti empo l a bi ol og a del os radi cal es l i bres erauntema
extrao,
consi derado con desi nters e i ncredul i dad por l a mayor
parte de l os
bi l ogos. "
Afortunadamente stano es l a
si tuaci n actual , pues l a bi ol og a de l as
EROyl a patol og a
del estrs oxi dati vo, ysuposi bl eprevenci n,
despi ertan un
i ntenso y creci ente i nters, sobre todo en l o que respecta
a l as causas del
proceso de envej eci mi ento.
As , segnel reci ente comentari o de Vi j g y
Mul l er
( 2000) : "Lateor a gerontol gi ca propuesta por
DenhamHarman en 1956an
ofrece l a expl i caci n ms
atracti va de unmecani smo general
responsabl e del
envej eci mi ento. La di ferenci a es que, mi entras
en 1969 sl o hab a ci en
trabaj os publ i cados sobre radi cal es
l i bres, envej eci mi ento y enfermedad,
l a
canti dad aument a
2000 en 1990 yahora sermucho mayor
. El creci ente
nmero
de publ i caci ones se acompaa de un
i mpresi onante aumento en l a
vari edad de l os enfoques
metodol gi cos. Mi entras que al pri nci pi o
l a
i nvesti gaci n en este campo
se l i mi taba a l os estudi os
descri pti vos, ahora
i ncl uye
mtodos tanavanzados como el
usodeani mal es transgni cos"
.
1. 2. Defi ni ci n
ycl asi fi caci ndel os radi cal es
l i bres
Se defi ne como radi cal l i bre a
aquel l a especi e qu mi ca
( tomo o
mol cul a) capaz
de exi sti r i ndependi entemente, y que
conti ene uno o ms
el ectrones desapareados. Los radi cal es l i bres son
especi es dotadas de el evada
capaci dad de reacci n, y por tanto,
capaces de i nterveni r en
numerosas
reacci ones qu mi cas, al terando
el funci onami ento desi stemas
tanto bi ol gi cos
comono
bi ol gi cos ( Pryor, 1976) .
Los radi cal es l i bres
pueden ser de di sti ntos ti pos, segn
l os tomos
i ntegrantes, pero debemos destacar por su
i mportanci a en l os si stemas
bi ol gi cos al os radi cal es
l i bres deri vados del ox genoy rel aci onadas
especi es
reacti vas
del ox geno ( ERO) , as como al os
radi cal es l i bres deri vados del
ni trgenoyrel aci onadas especi es
reacti vas del ni trgeno ( ERN) ( SaranyBors,
1990;
Hal l i wel l , 1992; 1997) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I ntroducci n
1. 3. Mecani smosdeformaci ndel osradi cal esl i bres
Uncompuestopuedeconverti rseenradi cal l i brecaptando
operdi endo
unel ectrn ( e- ) . As mi smo, tambi n
puedenformarseradi cal es l i bres cuando
un enl ace coval ente se rompe, y cada el ectrn de l a parej a
comparti da
permanece en un tomo ( fi si n homol ti ca)
.
Por
otra parte, en l a fi si n
heterol ti ca
unodel os tomos reci bel os e- , quedandocargadonegati vamente,
mi entras que el otro adqui ere una carga posi ti va,
generando i ones y no
radi cal es
. Como consecuenci a de poseer el ectrones desapareados,
estas
especi es qu mi cas son muyreacti vas,
poseen unavi da medi a corta y se
encuentranenmuybaj a
concentraci nenel estadoestaci onari o ( Armstronget
al . , 1984, Pryor,
1986, Webster
y
Nunn. , 1988, Cadenas, 1989) .
Enl ossi stemas bi ol gi cos l a fi si n heterol ti caes l a
mscomn, debi do
a que l a ruptura homol ti ca requi ere
unaenerg a el evada ( temperatura,
radi aci n
ul tra- vi ol eta o radi aci n i oni zante) , as general mentese
producel a
donaci ndee- deunamol cul aaotra como en
reacci ones redox, reacci ones
no enzi mti cas de
transferenci a de e- , reacci ones catal i zados
por metal es o
procesos catal i zadospor enzi mas ( Hal l i wel l y
Gutteri dge, 1989) .
Los radi cal es
l i bres pueden tener un ori gen endgeno o
exgeno al
medi o
bi ol gi co, yadems su formaci n puede estar l i gada a
mecani smos
fi si cos ( comoradi aci nul travi ol etaei oni zante) o
bi enamecani smosbi ol gi cos
como l a producci n cel ul ar de
radi cal es l i bres que puede ocurri r
por
reacci ones enzi mti cas
onoenzi mti cas ( Pryor yStone,
1993) .
A
conti nuaci n
descri bi mos al gunasdeestasreacci ones
:
Reacci ones de
oxi do- reducci n. Como l a transformaci n
de
hi droqui nonaasemi qui nona, enl aquei ntervi ene
el
02
quesetransformaen
superxi do
( 02
. -
)
( Wei ss et al . , 1987) .
Reacci ones de autooxi daci n de pequeas
mol cul as del ci topl asma
cel ul ar como catecol ami nas, fl avi nas,
qui nonas yti ol es, queproducen
oxi -
radi cal es debi do aunareducci n
concomi tantedel ox geno( Fi sher y
Kaufman,
1973; Baccanari ,
1978; Fri dovi ch, 1983; Proctor y
Reynol ds, 1984) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I ntroducci n
Reacci ones
de l a cadena de transporte de el ectrones (CTE) . Comoya
hemos comentado, una
de l as mayores f uentes productoras deradi cal es l i bres
es l a mi tocondri a, debi doaquems del 95%del 02 es uti l i zadopor el l a para
real i zar l a f osf ori l aci noxi dati va, que
se encuentra acopl ada al a cadena de
transporte de el ectrones
(CTE) . La CTE es unav a ni ca ya que enel l a l os
tomos de hi drgeno
(H) sonesci ndi dos enprotones (H+) y
e- . Los e- pasana
travs de una seri e de transportadores
de e- de l a membrana mi tocondri al
i nterna, yenal gunos de estos pasos a
l o l argode l a cadenal os protones yl os
e- se recombi nan
entre el l os. Exi ste unatendenci a aque
estos e- reacci onen
di rectamenteconel
02
f ormandoas el
radi cal
02
- envez de pasar al si gui ente
transportador. Deah quel a
l i beraci nde 02- represente l a mayor
f uente de
producci nderadi cal es l i bres
encondi ci ones f i si ol gi cas (Harman, 1972
;
Nohl
yHegner, 1978;
Chanceet al . , 1979) .
Adems
de l a producci n de 02'
-
por l a CTE
en l a membrana
mi tocondri al i nterna, tambi n
se f orma perxi do de hi drgeno (H202) enl a
membrana mi tocondri al
externa acausade l a desami naci n
oxi dati va de l as
catecol ami nas por el enzi mamonoami no
oxi dasa (FnanyCohen. , 1992) .
Otraf uente i mportante deproducci n
del radi cal
02' -
es el denomi nado
"estal l i do respi ratori o",
queti enel ugar enl as cl ul as
f agoc ti cas cuandostas
son acti vadas por cual qui er
est mul o. Este proceso
es l l evado a cabo por
ni coti nami da
adeni na di nucl eti do f osf ato oxi dasa
(NADPH oxi dasa), que se
encuentra enl a superf i ci e externa
de l a membranapl asmti ca. El proceso
es
el si gui ente: l os
l eucoci tos pol i morf onucl eares y
macrf agos sonacti vados
debi do
al contacto conuna sustanci a
extraa, con l o que aumentan su
consumode ox genoyl o transf orman
enradi cal
02' - ,
que es converti do en
H202
y
ste asuvez enradi cal
OH' , radi cal al que es expuesta l a part cul a
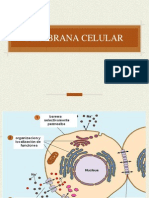
f agoci tada(McCord, 1974; Bl akeet al . , 1987) (Fi gura
1) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I ntroducci n
NADPH NADP+
02
0
02-
H202
OH*
NADPH
Oxi dasa
Fi gura 1.
Producci n de radi cal es l i bres durante el estal l i do respi ratori o
(Marti nez-Cayuel a, 1995)
HCI O
HI O
S ntesi s de prostagl andi nas. Ti ene l ugar aparti r
del metabol i smo del
ci do araqui dni co por medi aci n
de l as ci cl ooxi genasas (Porter, 1980) . La
s ntesi s de prostagl andi nas
ocurre en tres pasos: l i beraci n
del
ci do
araqui dni co de sus precursores l os f osf ol pi dos
del a membrana, oxi genaci n
del ci do araqui dni co
l i bre por prostagl andi na endoperxi do G/H
si ntetasa
(PGG/Hsi ntetasa) y metabol i smo de
endoperxi dos i ntermedi os (s ntesi s de
prostanoi des)(Smi th, 1986) .
Xenobi ti cos. La
f ormaci n de radi cal es l i bres est i mpl i cada
en l a
toxi ci dad de
un ampl i o rango de xenobi ti cos,
pues muchos compuestos
qu mi cos requi eren, para ej ercer su
acci n txi ca unaacti vaci n metabl i ca a
i ntermedi ari os
reacti vos quepuedenser di chos radi cal es l i bres (Mason, 1982
;
Proctor yReynol ds,
1984; Hal l i wel l y Gutteri dge, 1989)
. La f ormaci nde estos
radi cal es l i bres i ntermedi ari os
est catal i zada por l a
NADPH-ci tocromo P450
reductasa,
una f l avoprote na que uti l i za
NADPHcomo dador el ectrni co
(Kappus ySi es, 1981; Trushet al
. , 1982) . Enestos casos, seestabl eceunci cl o
redox en el que hay
ungran consumode NADPHy ox geno
y generaci n de
radi cal es
superxi do y otras especi es
reacti vas del ox geno (Fi gura 2) . Este
ci cl o redoxconduceaunasi tuaci n
deestrs oxi dati voy, encasos
extremos, a
l a muertecel ul ar
(Si es, 1985) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
NADPH-. _
Xenobi ti co <
~~
~~02-
Ci cl o
REDOX
Radi cal
NADP+
Xenobi ti co
Fi gura 2. Ci cl o redox
dexenobi ti cos (Mart nez- Cayuel a,
1998)
1. 4. Ox genoy
radi cal es l i bres del ox geno
En l os organi smos vi vos,
el
02
es el pri nci pal
responsabl e de l a
formaci n de radi cal es
l i bres (Tabl a 1) . La mol cul a
de ox geno puede ser
cal i fi cada de
bi rradi cal puesto que ti ene
dos el ectrones desapareados,
cada
uno de el l os l ocal i zado
en un orbi tal anti enl azante
n* . Si n embargo, l a
reacti vi dadde
esta mol cul aes ms baj ade
l a quecabr a esperar debi do
aque
l os espi nes de estos
dos el ectrones presentan
di recci ones paral el as
(Mal mstrm,
1982; Webster y Nunn, 1988;
Cadenas, 1989; J ami eson,
1989) .
Segnel pri nci pi o de excl usi n
dePaul i , l os el ectrones
deunorbi tal atmi coo
mol ecul ar nuncati enen
di recci ones paral el as. Por l o
tanto, paraque el ox geno
mol ecul ar
aumente su reacti vi dad ti ene
que i nverti rse el esp n de uno
de l os
el ectrones
de sus orbi tal es externos,
o bi en, sufri r unareducci n
secuenci a) y
uni val ente
produci endoradi cal es l i bres de
ox geno.
Tabl a 1.
Radi cal es l i bres del oxi genomsi mportantes
.
I ntroducci n
Nombre
s mbol o
natural eza
ox geno
si ngl ete 102
noradi cal
ozono
Os noradi cal
xi do n tri co
NO
radi cal
di xi do de ni trgeno
N02
radi cal
perxi dode hi drgeno
H202 noradi cal
hi droxi l o
OH'
radi cal
superxi do
02
. -
radi cal
hi droperxi do HOO'
radi cal
al coxi l o
RO' radi cal
peroxi l o
ROO'
radi cal
aci cl oxi l o R- COO'
radi cal
aci l peroxi l o
R- CO- COO'
radi cal
ari l oxi l o Aro'
radi cal
ari l peroxi l o
Ar00'
radi cal
ci dohi pocl oroso
HC10 noradi cal
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I ntroducci n
El ox geno se puede reduci r por
dos v as para transf ormarse tras vari os
pasos, en una mol cul a de agua ( H20) . La v a
tetraval ente es
el
pri nci pal
mecani smo de reducci n del 02 ( Naqui et al . , 1986) , ypor l a v a uni val ente
( Fi gura 3) se produce una pequea pero
conti nua reducci n del ox geno, que
es
l l evada a cabo en l a cadena
respi ratori a mi tocondri al medi ante l a NADH
oxi dasa yl a ubi qui nona ( Hal l i wel l et al . , 1992) .
Fi gura
3. V auni val ente
dereducci ndel ox geno ( Mart nez- Cayuel a,
1995)
.
1
. 5. Propi edades del os radi cal es l i bres del ox geno
Ox geno si ngl ete ( 1
02) .
Es el estado
exci tado i nf eri or del ox geno
mol ecul ar que se puede generar
en reacci ones enzi mti cas y f otl i si s del
ox geno
a
al tas
presi ones, a parti r de perxi do de hi drgeno ( Cadenas,
1989) .
Este
ox geno si ngl ete puede i nteracci onar con
otras mol cul as, bi en
combi nndose qu mi camente con el l as o
transf i ri endo su energ a, yvol vi endo
a su estado basal . La
f ormaci n de este radi cal en l os
si stemas bi ol gi cos,
puede estar medi ada por di f erentes
reacci ones enzi mti cas o por
qui mi oexci taci n durantel a
peroxi daci n l i p di ca ( Cadet et al . ,
1994) .
Ox geno 302
102 Oxi geno
si ngl ete
1 e-
Radi cal Superxi do
02
-
e-
I n
perxi do
2H+
H202
Perxi do de hi drgeno
02
2-
1 e-
2H+
023
H
2
0
Agua
H
I n oxeno
O- -
0
OH' Radi cal
hi droxi l o
1 e-
2H+
I n xi do
02- H2O Agua
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I ntroducci n
Radi cal
superxi do ( 02
'
- )
. En l as reacci ones de este radi cal predomi nan
l os procesos de transf erenci a
el ectrni ca, como su parti ci paci n en l a
reducci ndel i n f rri co
( Fe3+) af erroso ( Fe2+) , quees converti do a
perxi do de
hi drgeno
por el enzi masuperxi do di smutasa
( SOD) . Por otro l ado, carece
de
l a reacti vi dad suf i ci ente
para atacar a macromol cul as aunque
exi sten
numerosos bl ancos bi ol gi cos
sensi bl es al 02' _ ( McCordy
Fri dovi ch, 1969) .
El
02'
-
se ori gi na en muchas
reacci ones enzi mti cas pero su pri nci pal
f uente deproducci n
sonl as mi tocondri as, transf ormndose
en di cho radi cal
el 1%del
02
consumi dopor l as mi smas
( Ri chter, 1988) .
Radi cal hi droxi l o ( OH' ) .
Es el radi cal ms reacti vo en l os
si stemas
bi ol gi cos. Se
ori gi na enprocesos deradi l i si s,
f otl i si s del aguaydel perxi do
de hi drgeno. En l os seres vi vos,
una de l as pri nci pal es f uentes de radi cal
hi droxi l o es l a reacci n de
"Fenton" ( reacci n 1) :
( 1) Fe2+ +
H202
- > Fe3+ +OH- +OH'
( 2)
02 +
Fe3+ 402
+Fe2+
( 3) H202 + Fe2+
OH' + OH-
+Fes+
( 4)
02' - +
H202
02 + OH' +OH-
La reacci n
de Fenton ti ene l ugar
en condi ci ones f i si ol gi cas,
dependi endo
de l a presenci a de
superxi do o de otros agentes
capaces de
reduci r al hi erro. Lareacci n
se produce del a si gui ente
manera: enl a pri mera
etapa
( reacci n 2) el superxi do reacci ona
con trazas de metal , como
por
ej empl o Fe3+ o Cu2+
presente en l as cl ul as, ori gi nando l as especi es
reacti vas
de di chos i ones
as como perxi do de
hi drgeno. Aconti nuaci n, l a mol cul a
de perxi do de hi drgeno
es reduci da por di chos cati ones
produci endo el
radi cal
hi droxi l o ( reacci n 3) . Esta l ti ma
especi e, al ser def i ci ente en e- ,
se
convi erte en un poderoso
oxi dante y por tanto i ni ci ador de reacci ones
en
cadenamedi adas por
radi cal es l i bres ( Hal l i wel l yGutteri dge,
1984) .
El OH' es
consi deradounode l os radi cal es ms
dai nos paral a cl ul a,
ya que di f unde rpi damente del
l ugar donde se f orm a l as cl ul as
de
al rededor, se une a
mol cul as i nsaturadas, i ntervi ene en
procesos de
transf erenci a
de e- , yse encuentra i mpl i cado
en procesos bi ol gi cos de gran
i mportanci acomo
l af agoci tosi s ( Zi gl er et al . , 1985) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I ntroducci n
Perxi do de Hi drgeno
( H2021.
No es unverdaderoradi cal l i bre, ya que
posee dos e- apareados, pero el enl ace entre l os tomos del ox genoes si mpl e
por l o que resul ta bastante i nestabl e y f ci l
de romper
.
Esta mol cul a se
descompone espontnea yrpi damente baj o l a acci n de l a l uz vi si bl e. El
radi cal
superxi do acta comoprecursor cuandoes di smutadopor el enzi ma
superxi do di smutasaque catal i zal a si gui ente reacci n:
02.
+02* +
2 H+ ~
H202 +02
El H202 entra a f ormar parte de l a reacci n de Fenton, yes uti l i zado
como agente bacteri ci da por l os peroxi somas,
en reacci ones de f agoci tosi s
( Boveri s
et
al . ,
1972) .
1. 6. Ni trgenoyradi cal es l i bres del
ni trgeno
El ni trgeno
consti tuye aproxi madamente el 78, 03%del vol umen del
ai re. Al i gual queocurre conel ox geno, desempeauni mportante papel
enl os
si stemas bi ol gi cos al f ormar parte
de casi todas l a bi omol cul as.
Las i ntensas
i nvesti gaci ones sobre el xi do n tri co y rel aci onados
radi cal es
l i bres hanl l evado acrear l os nuevos trmi nos de
especi es reacti vas
de ni trgeno" ( ERN) y"radi cal es l i bres del
ni trgeno" ( Tabl a 2) ( Moncadaet al . ,
1991;
Rubboet al . , 1996; McAndrewet al . , 1997) .
Tabl a2. Radi cal es l i bres del Ni trgenoms i mportantes
Nombre s mbol o natural eza
xi don tri co
NO' radi cal
di xi dode ni trgeno N02' radi cal
ci doni troso
HN02 noradi cal
tri xi do de di ni trgeno
N202 noradi cal
tetraxi do de di ni trgeno
N204 noradi cal
peroxi ni tri to
ONOO- radi cal
al qui l - peroxi ni tri tos LOONO
noradi cal
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I ntroducci n
1 . 7. Propi edades
del os radi cal es l i bres deni trgeno
Oxi do n tri co (NO) . Es
consi derado como uno de l os radi cal es ms
i mportantes del N2;
se forma medi ante l a transformaci n de L- argi ni na
en
L- ci trul i na
por el enzi ma oxi do n tri co si ntetasa
(NOS), del a que se conocen
dos i soformas funci onal es: l aconsti tuti vayl a
i nduci bl e(Pal mer et al . , 1988) .
El NOes un pequea
mol cul a l i pof l i ca que di funde a l as
cl ul as
adyacentes, penetra en el ci tosol y
desempeaun i mportante papel en l a
seal i zaci n cel ul ar
y en l a i nducci n de di ferentes
procesos patol gi cos
(Moncada et al . , 1991) . Segnl as
condi ci ones que rodean l a producci n de
NO, ste puede ser
transformado enotras especi es reacti vas
i ntermedi as del
N2
como son el cati n ni troso (NO+), el ani n
ni troxi l o (NO- ) oel peroxi ni tri to
(ONOO- ) (Staml er et al . , 1992) .
La ci totoxi ci dad del
NOse atri buye al a i nhi bi ci n de l a
respi raci n
mi tocondri al por
uni nal compl ej o I Vmi tocondri al
(Bol aos et al . , 1994) yal
compl ej o I provocando as l a
i nhi bi ci n de l a fosfori l aci n oxi dati va y
un
descenso enl a
s ntesi s del ATP(Cl ementi et
al . , 1998) . Otro procesoci totxi co
es el efecto i nhi bi dor
queej erce sobreel enzi magl utati onperoxi dasa
(Asahi et
al . , 1995) .
An n ni troxi l o (NO- ) . Ti ene
efectos di rectos sobredi versas mol cul as
o
si stemas bi ol gi cos
(Li pton et al . , 1993) . Seformapor
reducci ndel NOpara
dar
el ani n ni troxi l o en una reacci n
catal i zada por el enzi ma superxi do
di smutasa (Murphy y S es, 1991),
o tambi n por l a uni n del NOal
Fe2+
perteneci ente aun
grupohemo del ci tocromo c (Sharpey
Cooper, 1998) ode
l a
hemogl obi na(GowyStaml er. ,
1998) .
Cati n ni troso
(NO+) . Es l a forma oxi dadadel NO,
rel aci onadaconel
ci do ni troso, yespeci ecl ave en
el procesode"ni trosaci n", en el cual el NO+
es transferi do a
un centro nucl eof l i co, normal mente de azufre o
ni trgeno
(Wi l l i ams, 1988) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I ntroducci n
Peroxi ni tri to (ONOO- ) . Lapri nci pal v ade formaci n
de
peroxi ni tri to es
l a reacci ndel NOconel
radi cal 02'
-
(Beckmanet al . , 1990) . El peroxi ni tri toes
capaz de reacci onar con l a mayor a de l as bi omol cul as, mostrando
ci totoxi ci dad aunque noestnpresentes enel medi o
el NOoel superxi do.
Estaci totoxi ci dad se debe asurpi dadegradaci na
radi cal OH' . Posee una
vi da medi a muy corta de aproxi madamente un segundo en condi ci ones
fi si ol gi cas (Beckmanet al . , 1990) .
En l a Fi gura 4, se muestran l as
pri nci pal es v as de producci n de
radi cal es
l i bres .
Metabol i smocel ul ar
Mi tocondri a
Si stemas enzi mti cos
Catecol ami nas
Uni val ente
NAD(P) Hoxi dasa,
XO, LPO, COX,
Ci tocromoP450.
02* - - )
OH* ,
Acti vaci n
l eucoci tari a
Xenobi ti cos, herbi ci das,
contami nantes, frmacos,
Expl osi nrespi ratori a
I rradi aci ones, al cohol . . .
Fi gura4. Ori gende l os radi cal es l i bres
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I ntroducci n
2. Si stemas dedefensa
cel ul ar
Los organi smos
superi ores han desarrol l ado, con el
transcurso del a
evol uci n, unefi caz si stemadedefensa
anti oxi dante(FreemanyGrapo, 1982) .
Hay
si stemas enzi mti cos detoxi fi cadores
y mecani smos "captadores",
denomi nados "defensas
pri mari as o preventi vas", que son capaces
de
di smi nui r l a
concentraci n de radi cal es l i bres, mi entras
quel as defensas
"secundari as", tambi n l l amadas
"rompe- cadenas", actan atrapando o
captando a
l os radi cal es l i bres ya formados, deteni endo
as l os efectos
perj udi ci al es en l as pri mares fases
del as reacci ones (Armstronget al . , 1984;
Cotgreave et al . ,
1988; Cadenas, 1989; Hal l i wel l
et
al . ,
1992; Tosaki et
a1
. , 1993) .
2. 1. Anti oxi dantes
pri mari os
Superxi do di smutasa (SOD) . Forma parte
de una fami l i a de
metal oenzi mas con di ferentes grupos
prostti cos, del ocal i zaci n i ntracel ul ar
vari abl e y gran
heterogenei dad ti sul ar . El i soenzi ma predomi nante
es l a
superxi do
di smutasa dependi ente de
zi nc y cobre (CuZnSOD), que se
encuentra en l a
mayor a de cl ul as eucari otas (Fi sher y
Kaufman, 1973,
Fri dovi ch,
1975; McCord yFri dovi ch, 1977;
Fri dovi ch, 1983) . Tambi nseha
encontrado unaSOD
de al to peso mol ecul ar extracel ul ar (Markl und
et al . ,
1982; Markl und,
1984) . Estos enzi mas catal i zan
l a transformaci n de
superxi do enperxi do dehi drgeno
yox geno.
SOD
02
. -
H202+02
Catal asa
(CAT
) .
Este enzi matransformael perxi do de
hi drgeno del as
cl ul as en H2Oy02
cuando
l a
concentraci n es el evada. Sel ocal i za en l os
peroxi somas
. Al i gual queSOD, CATseencuentraampl i amente
di stri bui daen
todos l os tej i dos. (Proctor yReynol ds, 1984;
Turrens et al . , 1984; Bl akeet al . ,
1987) .
H202
CAT
10
H20+1/ 2 02
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I ntroducci n
Gl utati on
peroxi dasa ( GPx) . Catal i za l a reducci n del peroxi do de
hi drgeno
yde l os perxi dos orgni cos l i bres. Requi ere gl utati on
reduci do
( GSH) comosustrato.
acti vi dad catal ti ca.
cl ul as eucari otas,
Proctor yReynol ds,
1984; Cotgreave et al . , 1988; Spal l hol z y
Boyl an. , 1991;
Harri s, 1992) .
Gl utati on
reductasa. Es un enzi ma ci tosl i co que
reduce el gl utati on
oxi dado uti l i zando NADPHgenerado
por otros si stemas ( Proctor yReynol ds,
1984; Cotgreave
et al . , 1988) .
Otras prote nas. Los metal es de transi ci n
tambi n estni mpl i cados en
l a generaci n de radi cal es
l i bres por medi o de l a reacci n de Fenton
. Si n
embargo, cuando
estos metal es se encuentran uni dos aprote nas
no pueden
catal i zar
l a reacci n.
La concentraci n de hi erro l i bre
en el organi smo est control ada
en
condi ci ones f i si ol gi cas,
al encontrarse el hi erro uni do a
gl ucoprote nas como
transf erri na o l actof erri na, que
l o transportan en l a ci rcul aci n o bi en
a l a
f erri ti na, quel o
al macena. Por l o tanto, estas prote nas regul an
i ndi rectamente
l as
reacci ones de oxi do- reducci n ( Hal l i wel l y
Gutteri dge. , 1984,
1985;
Thurnham, 1990) .
La al bmi na
yl a cerul opl asmi na son l as
encargadas de transportar el
cobre
en el pl asma, aunque no i mpi den l a
i nteracci n entre el cobre yl os
radi cal es l i bres ( Bl ake et al . , 1987; Thurnham,
1990) .
GPx
H202+
2 GSH
GSSG+2H20
GPx
ROOH
+
2
GSH
10 GSSG+ROH
+
H20
Adi f erenci a de l a catal asa, l a GPx posee una al ta af i ni dad
por el
sustrato, ya que posee 4tomos de sel eni o, que
son l os responsabl es de l a
Se encuentra mayori tari amente
en el ci topl asma de l as
parti cul armente en l as mi tocondri as ( Wendel ,
1980;
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I ntroducci n
Pequeas mol cul as
. Exi stendi f erentes mol cul as
depequeo tamao
como gl utati on, ci do
ascrbi co, ci do ri co y tauri na,
di stri bui das
ampl i amenteentodos
l os si stemas bi ol gi cos,
quesoncapaces deel i mi nar l os
radi cal es
l i bres medi ante reacci ones
noenzi mti cas (Freemany
Grapo. , 1982;
Bast et al . , 1984; Cadenas,
1989) .
2. 2. Anti oxi dantes
secundari os
Exi sten
di f erentes gl utati on
transf erasas con acti vi dad
peroxi dasa,
dependi entes del
gl utati on, que actan como
anti oxi dantes f rente a l a
peroxi daci n
l i p di ca. Estos enzi mas
metabol i zan l os hi droperxi dos
de baj o
peso mol ecul ar pero nunca el
H202, y
adems
necesi tan l a acti vi dad de l a
f osf ol i pasa A2
paraser f unci onal es
(Ketterer, 1986; Ketterer y
Meyer. , 1989;
Davi es et al . , 1990) .
Exi sten
otros enzi mas como l as que
reparan el ADN, es deci r
ADN
pol i merasa
I yADNl i gasa,
tambi n consi deradas
mecani smos de def ensa
f rente al daocausado
por el estrs oxi dati vo,
yaquecuandosef ormaun
si ti o
apur ni coo
api ri mi d ni co por l esi noxi dati va
sedeti enel a repl i caci n
del ADN
y actan estos enzi mas reparando
l a rotura. Tambi n l as
endonucl easas
ti enen un i mportante
papel protector
del ADNcel ul ar y del f l uj o
de
i nf ormaci n
genti ca (Cotgreave
et al . , 1988; Davi es et al . ,
1990) . I gual que
sucede con l a
al teraci n de l as
prote nas, cuando el dao al ADN
es
i rreparabl e, l as cl ul as son
el i mi nadas.
Adems de estos
enzi mas, exi sten otras
mol cul as que actan como
anti oxi dantes
secundari os. El a- tocof erol
o vi tami na E(VE) es el pri nci pal
anti oxi dante l i posol ubl e,
presente en todas l as membranas
cel ul ares, que
actaprotegi endo
f rente al aperoxi daci n l i p di ca. Para
el l o reacci onaconl os
radi cal es
l i bres donandouni nhi drgenoy
convi rti ndol os en f ormas menos
reacti vas. El radi cal tocof eri l o
tambi npuedeser reduci do
di rectamentepor el
tndemgl utati on- ci do ascrbi co,
descri to a conti nuaci n (Burton et
al . ,
1982; Cadenas, 1989) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I ntroducci n
Otro anti oxi dante
secundari o es el / 3- caroteno ( el anti oxi dante ms
ef i caz para el i mi nar el ox geno si ngl ete) , que
poseeunaacci nsi nrgi ca conl a
VE
( Burtone I ngol d, 1984; Machl i nyBendi ch. , 1987; Bendi chyOl son, 1989) .
2. 3. ci do ascrbi cooVi tami na C
La vi tami na C( VC) acta como un ef i caz anti oxi dante hi drosol ubl e en
l os f l ui dos bi ol gi cos ( Fre et al . , 1989, 1990) el i mi nando f i si ol gi camente
radi cal es l i bres tanto del ox geno
como del ni trgeno ( Hal l i wel l , 1996) . Entre
estos se i ncl uyen hi droxi l os, peroxi l os, ani n superxi do, y di xi do de
ni trgeno, as como especi es no radi cal es como
ci do hi pocl oroso, ozono,
ox geno si ngl ete, ni trxi do yperoxi ni tri to
.
Adems
de el i mi nar radi cal es l i bres, l a VCpuede regenerar otras
mol cul as anti oxi dantes como a- tocof erol , gl utati on, uratos
yR- caroteno, a
parti r de sus
respecti vas f ormas radi cal es ( Hal l i wel l , 1996) .
La hi ptesi s de que l a VCes
un ef i caz anti oxi dante se apoya en
numerosos datos,
entre l os cual es destaca que tanto el ascorbato como su
radi cal l i bre ( f ormado por l a prdi da de 1 e- de l a
mol cul a del ascorbato)
ti enen baj os potenci al es de reacci n
( Buettner, 1993) , por l o que pueden
reacci onar conotros radi cal es y
oxi dantes presentes enl os si stemas bi ol gi cos
( Buettner y
J urki ewi ck. , 1996) . Adems, el ascorbato puede ser regenerado a
parti r de radi cal l i bre ascorbato y
ci do dehi droascrbi co tanto por v as
enzi mti cas como
noenzi mti cas. El radi cal l i bre ascorbatoes reduci do por
l a
semi dehi droascorbato reductasa dependi ente de NADH
( Wel l s yJ ung, 1997) y
por l a ti oredoxi n
reductasadependi ente de NADPH( Mayet al . , 1998) ( Fi gura
5) .
HOCH2
HOCH
Ascorbato
HOCH2
HOCH2
HOCH n HOC
Radi cal
ci do
Ascorbato
dehi droascorbi co
Fi gura 5. Metabol i smoredox del
ci doascrbi co.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I ntroducci n
2. 4. N- aceti l - ci ste na
El anti oxi dante
ti l i co N- aceti l - ci ste na (NAC) puede
actuar a travs de
di versos mecani smos, ya sea
como precursor del gl utati on, que
es un
anti oxi dante
pri mari o (De Vri es y De Fl ora, 1993),
o bi en esti mul ando l a
acti vi dad de al gunos enzi mas ci tosl i cos
i mpl i cados enel ci cl o del
gl utati on
(De Fl ora et al . ,
1991), comopor ej empl ol a gl utati on
reductasa, que aumenta
l a tasa
de regeneraci n del gl utati on. As ,
NACpuedeaumentar l os ni vel es
de
gl utati onreduci do enl a cl ul a
proporci onando unaf uente de
ci ste na para su
bi os ntesi s,
con el consi gui ente aumento de l a acti vi dad
gl utati on reductasa
(Col ton
et al . , 1995) .
Unsegundomecani smo
de acci nes por reacci n di recta
del grupoti ol
reduci do
de NACconradi cal es l i bres (Auroma
et al . , 1989) .
Tambi nes probabl e
unef ecto esti mul ante de NACsobre
el proceso de
f osf ori l aci n
oxi dati va mi tocondri al (Mi quel
et al . , 1995), al proteger
a l as
prote nas que i ntervi enenen
di choprocesof rente aposi bl es
daos produci dos
por el
estrs oxi dati vo. De este modo, se
manti enenen estado reduci do l os
grupos sul f hi dri l os de esas
prote nas, que son esenci al es para l a
acti vi dad
enzi mti ca (Haugaard
et al . , 1969; Yagi yHatef i . , 1984) .
Otro
mecani smo de acci nde NACes su acci n
protectora f rente a l a
peroxi daci n l i p di ca en l as
membranas mi tocondri al es, con l a
resul tante
preservaci nde l a acti vi dadde l os compl ej os respi ratori os
(Zhang et al . , 1990) .
Adems de estos ef ectos
protectores de NAC, un nmero
creci ente de
observaci ones apoyan su papel enl a i nhi bi ci n de
apoptosi s en cl ul as f i j as
postmi tti cas y ol i godendroci tos
(Mayer y Nobl e, 1994; Ferrari et
al . , 1995;
Yany
Greene, 1998) . As , l a admi ni straci nde NACposee
ef ectos benef i ci osos
f rente a apoptosi s en enf ermedades
neurodegenerati vas rel aci onadas con el
envej eci mi ento (Ratan
et
al . ,
1994 a, b), adems de i nhi bi r l a expresi n
de l os
genes
c- f os ycj uni mpl i cados enl a apoptosi s. Enl a
Fi gura 6, se resumenl os
ef ectos de NAC.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I ntroducci n
Ci stei na
S ntesi s de GSH
El i mi nador
I nhi bi ci n apoptosi s
Fosf ori l aci n oxi dati va
Regeneraci nGSH
de radi cal es l i bres
Ras, c- f os,
c- i um
mi tocondri al .
Compl ej os I , I V, V
1
Aumento
Di smi nuci ndel dao
di sponi bi l i dad del ATP
cel ul ar
Supervi venci a
Cel ul ar
Fi gura
6.
Ef ectos deN- aceti l - ci ste na
(Banacl ocha, 2001 )
2. 5. ci do ti azol i d n carbox l i co o ti oprol i na
El ci do
ti azol i d n carbox l i co o ti oprol i na (TP) es un ami noci do
sul f hi dr l i co c cl i co, quees uti l i zado cl ni camente como
hepatoprotector . Acta
como donador de grupos sul f hi dri l o i ntracel ul ares y
como captador de
radi cal es l i bres
(Dansette et al . , 1 990) yha mostrado ef ectos
benef i ci osos
sobre l a l ongevi dad yf unci ones f i si ol gi cas del ratn ydel
i nsecto Drosophi l a
mel anogaster (Mi quel yEconomos,
1 979) .
Tambi n se ha observado que l a admi ni straci n de
anti oxi dantes
ti l i cos, como gl utati on o TP, previ ene l a
prdi da de coordi naci n
neuromuscul ar de l os ratones, segn se demuestra
medi ante el test de l a
cuerda ti rante (Mi quel yBl asco. , 1 978
;
Pal l ard
et al . , 1 998) .
Otro ef ecto
de l a admi ni straci n de estos anti oxi dantes, observado en
mi tocondri as de
h gado y cerebro de ratn, es un aumento del
coci ente
GSSG/GSHyuna di smi nuci n en el dao, produci do
por el estrs oxi dati vo
l i gado al
envej eci mi ento, en el ADNmi tocondri al (Garc a de l a Asunci n
et al ,
1 996; Sastre et al . , 2000a) . Otros estudi os real i zados con
ratones envej eci dos
cuya di eta estaba supl ementada con TP,
demostraron un ef ecto de TP
esti mul ante
sobre l as cl ul as del si stema i nmunol gi co
(De l a Fuente et
al . , 1 998a) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
ERO
ANTX
ERO
Condi ci ones
Estrs
f i si ol gi cas
oxi dati vo
I ntroducci n
3. Daocel ul ar por estrs oxi dati vo
El
estrs causadopor l os radi cal es l i bres, estrs oxi dati vo,
aparece
cuando
l a
producci n de estos radi cal es supera l a capaci dad
del as def ensas
anti oxi dantes, bi enpor undescenso
enl os ni vel es deestas def ensas opor
un
aumentoen
l a producci ndel os radi cal es (Fi gura 7) . El estrs
oxi dati vopuede
l esi onar vari os ti pos de cl ul as di ana, y
l a l esi n de stas puedea su vez
ori gi nar l a
l esi ndeotras (Hal l i wel l , 1991) .
ANTX
Fi gura 7. Equi l i bri oy
desequi l i bri oestrs oxi dati vo/ def ensasanti oxi dantes
La
presenci a deradi cal es l i bres es necesari a para
l as cl ul as. Dehecho,
estos radi cal es
son produci dos conti nuamente en l os organi smos
y son
esenci al es
enprocesos f i si ol gi cos tan i mportantes
como l a f agoci tosi s yl a
comuni caci n i ntercel ul ar .
Si n embargo, cuando aumenta excesi vamente l a
producci n de
radi cal es l i bres o se han deteri orado
l os mecani smos
anti oxi dantes, aparece el dao cel ul ar
(Fri dovi ch, 1983; Armstrong et al . ,
1984; Pryor, 1986; Hal l i wel l
yGutteri dge, 1989; Val enzuel ayVi del a,
1989) .
La
mayor a del os radi cal es l i bres dei nters
bi ol gi co suel en ser muy
reacti vos, i nestabl es yde vi da medi a muy
corta, quese puedemedi r i ncl uso
en f racci ones de
mi crosegundo por l o cual suel en causar l esi ones
cerca del
l ugar desuproducci n.
Noobstante, cuandounradi cal l i brereacci ona con
un
compuesto
no radi cal puedenf ormarseotros radi cal es l i bres,
demanera que
es posi bl equesecreenreacci ones en
cadena ydenl ugar a ef ectos bi ol gi cos
l ej os del si stema
que
ori gi n
el pri mer radi cal . Uni mportanteej empl o de
este
mecani smodesorgani zador es l a peroxi daci n l i p di ca (Pryor,
1986; Bl ake et
al . , 1987; SouthornyPowi s, 1988; Webster
yNunn1988; Harman, 1992) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I ntroducci n
3. 1. Daoaprote nas
Las prote nas sonal teradas por l os radi cal es
l i bres de ox geno atravs
de vari os mecani smos. Por
ej empl o, pueden reacci onar di rectamente conel
l i gando
metl i co de muchas metal oprote nas, yas se hacomprobado que
el
hi erro de l a oxi hemogl obi napuede reacci onar con
el radi cal superxi do o el
perxi do de hi drgeno para
f ormar metahemogl obi na (Freeman y Grapo,
1982) . Por otra
parte, l ahemoprote naci topl asmti cacatal asaes i nhi bi dapor
el
02
. - , que
l a
convi erte en sus f ormas i nacti vas f erroxi yf erri l o
(Kono et al . ,
1982) .
Debi do a l a reacci n de l os radi cal es l i bres
con l as mol cul as
i nsaturadas o que conti enen
azuf re, l as prote nas conproporci ones el evadas
de tri ptf ano,
ti rosi na, f eni l al ani na, hi sti di na, meti oni na y
ci ste na pueden
suf ri r modi f i caci ones acausade
di chos radi cal es. As se haobservado que
enzi mas tal es como l a
papa na, l a gl i ceral deh do- 3- f osf ato deshi drogenasa
e
i ncl uso l a superxi do
di smutasa, quedependende estos ami noci dos
paraser
acti vas,
soni nhi bi das enpresenci ade radi cal es
l i bres. Las reacci ones de l os
radi cal es con
l os ami noci dos menci onados, dan l ugar
a cambi os
estructural es
en l as prote nas, con f ormaci n
de puentes di sul f uro i ntra e
i ntermol ecul ares que
desencadenan procesos de agregaci n mol ecul ar
(Freemany
Grapo, 1982; Wol ppet al . , 1986
;
Webster
yNunn, 1988; Stadtman
1992; Gebi cki yGebi cki , 1993) .
Los enl aces
pept di cos tambi nsonsuscepti bl es de ser atacados
por l os
radi cal es
l i bres. Estos enl aces puedenromperse tras l a
oxi daci nde resi duos
de
prol i na
por radi cal es hi droxi l oo
superxi do (Wol f f yDean, 1986) .
Fi nal mente, l a
reacci ndel os radi cal es l i bres conl as prote nas
tambi n
puede
generar subproductos que aumentan
el dao i ni ci al . Un ej empl o l o
consti tuye l a oxi daci n del
tri ptf ano hastaH202 yN- f ormi l ki nureni na,
un
compuesto
que, al reacci onar con grupos ami no,
provocaentrecruzami entos
entre prote nas yl pi dos (Ni el sen,
1981 ; FreemanyGrapo, 1982;
Stadtman,
1992) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I ntroducci n
3. 2. Dao
al pi dos
Los radi cal es l i bres
hi droxi l o e hi droperoxi l o y el ox geno
si ngl ete
pueden
reacci onar con l os ci dos grasos
de l os f osf ol pi dos y otros
componentes l i p di cos de l as membranas
paraf ormar hi droperxi dos
l i p di cos
( Hal l i wel l yGutteri dge, 1984;
HortonyFai rhurst, 1987; Gutteri dge
yHal l i wel l
1990; Ai kens et al . ,
1991; Ni ki et al . , 1991; Schai ch,
1992; Choe et al . 1995;
Choi y
Yu, 1996) .
Este proceso
de peroxi daci n l i p di ca
comi enzacuando el radi cal l i bre
qui tauntomode hi drgeno de
unode l os carbonos meti l enos
de l acadena
carbonada para f ormar un
radi cal l i bre l i p di co ( L' ) .
Los ci dos grasos
pol i i nsaturados
de l as membranas son
especi al mente suscepti bl es
de ser
atacados por radi cal es
l i bres, puesto que conti enen
grupos meti l enos
separados por
dobl es enl aces que debi l i tanel enl ace C-H
meti l eno. Cuando
se
ori gi na el
radi cal l i p di co, suf re
i nmedi atamente un reaj uste
mol ecul ar,
produci ndose undi enoconj ugado
quepuedereacci onar
conel 02yf ormar un
radi cal
hi droperoxi l o. Este radi cal l i bre
dal ugar a endoperxi dos,
o bi en,
puede tomar untomo
de hi drgeno de un carbono
meti l eno de otro ci do
graso
pol i i nsaturadoadyacente paraf ormar,
de nuevo, unradi cal
l i bre l i p di co
yunhi droperxi do( Fi gura8) .
El radi cal l i p di co se
combi naconotramol cul a
de ox geno
y as se establ ece una
cadena de propagaci n del
dao
peroxi dati vo. Por suparte, el
hi droperxi do, que es uncompuesto
establ e, si
entraen contacto con
i ones metl i cos de transi ci n,
produci r ms radi cal es
l i bres que
i ni ci arn y propagarn otras
reacci ones en cadena. As , l as
membranas resul tan seri amente
daadas y, por tanto, se af ecta
sucapaci dad
f unci onal
( Ri ceevans yBurdan, 1993) .
Los productos f i nal es del
procesode peroxi daci nl i p di ca,
entre l os que
se i ncl uyen hi drocarburos
vol ti l es, al cohol es o
al deh dos, pueden di f undi r
l ej os del l ugar donde se ori gi naron
ycausar edemacel ul ar e
i nf l ui r en l a
permeabi l i dad
vascul ar, i nf l amaci n y qui mi otaxi s
( Bl ake et al . , 1987;
SouthornyPowi s, 1988; Pryor yChurch,
1991) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I ntroducci n
Estos productos puedenal terar l a
acti vi daddel a f osf ol i pasa ei nduci r l a
l i beraci n de ci do araqui dni co y l a subsecuente f ormaci n
de
prostagl andi nas ydi sti ntos endoperxi dos (Sevani an
et al . , 1981) .
- H'
- Oa 1 1 +O2H'
L'
+
/
\- /\- /
Hi droperxi do
l i pi di co
Al deh dosyproductos
de
pol i meri zaci n
Li pi do pol i i nsaturado
Radi cal l i brel i p di co
Di eno conj ugado
Radi cal
peroxi
Fi gura 8. Esquemadel a peroxi daci n
l i p di ca (Mart nez- Cayuel a, 1998)
El mal ondi al deh do (MDA), uno
de l os productos f i nal es de l a
peroxi daci n de ci dos grasos con
tres o ms dobl es enl aces, puede
causar
entrecruzami entos y
pol i meri zaci n dedi sti ntos componentes
del a membrana
(Ni el sen, 1981; Val enzuel a, 1991) . As
pues, l as propi edades de l as
membranas anresul tan
ms al teradas.
3. 3. Dao al ADN
Los ci dos nucl ei cos
tambi n pueden ser atacados por radi cal es
l i bres,
cuya ci totoxi ci dad
se debeengranpartea l as aberraci ones
cromosmi cas que
suf ren
l as bases yl os azcares del ADN
al reacci onar con di chos radi cal es,
especi al mente con el OH' (Dempl e y
Li nn, 1982; Webster y Nunn. ,
1988;
Harman, 1992;
Randerathet al . , 1992; Hi guchi yLi nn, 1994) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I ntroducci n
Di chas
modi f i caci ones qu mi cas de l os nucl eti dos
provocanenmuchos
casos l a ruptura
de l as cadenas de ADN. Si el dao que se
ori gi na es tan
grande
que nopuede ser reparado, se produce una
mutaci n, obi en l a cl ul a
muere (Hof f manet al . , 1984;
Cerutti , 1985; I ml ayyLi nn, 1988; Davi es
et al . ,
1990, Dempl e 1990) .
As , por ej empl o, se hacomprobado
que l a rupturadel
ADNque
ocurre durante el estal l i do respi ratori o,
produci dopor l os neutrf i l os
en
respuesta adetermi nados
est mul os, puede causar l a muerte de
estas
cl ul as e i nduci r procesos
autoi nmunes (Bi rnboi my
Kanabus-Kami nska,
1985;
Mart nez-Cayuel a, 1995) .
3. 4. Daoa
carbohi dratos
Los carbohi dratos son
daados por l os radi cal es l i bres
en menor
proporci n que otras
mol cul as. Azcares tal es como l a
gl ucosa, el mani tol o
ci ertos desoxi azcares
pueden reacci onar con el radi cal
OH' para produci r
sustanci as
reacti vas (Si es, 1985;
Bl ake et al . , 1987) . De i gual
f orma, l os
pol i sacri dos pueden
suf ri r el ataque de l os radi cal es
l i bres, f ragmentndose
enuni dades
ms pequeas. As , se ha
demostrado que el ci dohi al urni co
se
despol i meri zaenpresenci a
de concentraci ones el evadas
de radi cal es hi droxi l o,
provocando
un descenso de l a vi scosi dad
del l qui do si novi al
de l as
arti cul aci ones (Greenwal dy
Moy, 1980) .
4.
Apoptosi s i nduci dapor estrs
oxi dati vo
Lacaracter sti ca ms
sobresal i ente de l a apoptosi s es que
se trata de
unmecani smof i si ol gi co
de muerte cel ul ar quees necesari opara
mantener l a
pl asti ci dad de l os
tej i dos yel i mi nar l as pobl aci ones
cel ul ares excesi vas o que
se
tornanpel i grosas. Laapoptosi s
no causadaosecundari o ni
i nf l amaci n,
pues l a cl ul a
apoptti ca se separa de l as cl ul as
veci nas y acaba
f ragmentndose en cuerpos apoptti cos
que son el i mi nados por f agoci tos
del
entorno. Lanecrosi s,
por el contrari o, es unmecani smo
patol gi col i gado aun
dao
cel ul ar extremo, que af ecta a
l as cl ul as col i ndantes y
provoca
i nf l amaci n(Duke,
1992; SongyStel l er, 1999) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I ntroducci n
La apoptosi s es
un
mecani smo fundamental para
mantener l a
i ntegri dad de l os organi smos pl uri cel ul ares y su ej ecuci n evi ta procesos
patol gi cos como el cncer, o l as enfermedades autoi nmunes (Cohen, 1997;
Campi si ,
2000) . Por el contrari o, en ci ertas si tuaci ones patol gi cas se ha
demostrado que l a apoptosi s supone una prdi da i rreparabl e de cl ul as
necesari as para mantener
una funci n pti ma. As se ha demostrado que
muchas de l as neuronas que desaparecenenl as enfermedades de Al zhei mer y
Parki nson (Nagy y
Esi ri , 1997
;
Wul l ner et al . , 1999) y en l os acci dentes
cerebrovascul ares (Bennett et al . , 1998) l o hacen por apoptosi s. Las
consecuenci as patol gi cas de l a apoptosi s ti enen l ugar, aunque exi sten
mecani smos de protecci n que confi erena
l as
cl ul as
fi j as postmi tti cas (como
l as neuronas y l as cl ul as del mi ocardi o) su mayor resi stenci a a sufri r
apoptosi s. Esta resi stenci a est modul ada por prote nas i mpl i cadas
en
si stemas de transducci n
de
seal es
como Bcl - 2, y otras como Bax y Bad
(Tamatani et al . , 1998; Li uyZhu, 1999) .
4. 1. Regul aci nde
l os mecani smos de muerte cel ul ar
Dado el papel fundamental que desempeanl os procesos de
apoptosi s y
necrosi s en l a muerte neuronal que ti ene
l ugar en l as enfermedades
neurodegenerati vas y
en el envej eci mi ento fi si ol gi co (Nagy y Esi ri , 1997;
Bennett
et
al . ,
1998; Wul l ner et al . , 1999), parece oportuno consi derar al gunos
de l os mecani smos que regul an ambos procesos y
que se encuentran, en gran
medi da, l i gados
a
l a
capaci dad de autodupl i caci n de l as cl ul as (Mei krantz y
Schl egel , 1995) . De acuerdo con este concepto,
Campi si (1997) y Ly et al .
(2000), i nvesti garon l a
funci n de unel evado nmero de genes l l egando al as
si gui entes concl usi ones : pri mera, que al menos un 1%
de todos l os genes
exami nados sufren
al teraci ones dependi entes de l a edad, y segunda, que
muchos
de l os genes al terados ti enen rel aci n conl aprol i feraci n cel ul ar
.
No todas l as cl ul as de unorgani smo ti enen l a capaci dad
de prol i ferar
i ndefi ni damente.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I ntroducci n
Adems, l a prol i f eraci n
cel ul ar es un proceso
c cl i co que i ncl uye
di f erentes
etapas yquerequi erel a expresi n
coordi nada deuna seri e de
genes
quecodi f i can, por una parte
ci cl i nas y qui nasas
dependi entes del as ci cl i nas
(cdk regul adas
por di versos f actores
decreci mi ento) , y por
otro, determi nados
productos gni cos que
actan di recta o i ndi rectamente
sobre l os f actores
antes menci onados
bi en para desencadenar l a
prol i f eraci n (protooncogenes) ,
o para
i nhi bi rl a (genes supresores
detumores) (Schaf er, 1998)
.
Entre l os protooncogenes
destaca el responsabl e
de l a prote na ras,
rel aci onado con
l os mecani smos de
transducci n de seal es, as
como l os
genes myc,
f osyj un, quecodi f i can
f actores detranscri pci n,
rel aci onados con
l a acti vaci n del ci cl o
cel ul ar en respuesta a f actores
decreci mi entoycon
el
desencadenami ento
de cambi os
en el patrn deexpresi n
gni ca responsabl e
del control de l os
compl ej os ci cl i nas- cdk
(Reddel , 1998) . Entre
l os genes
supresores de tumores destacan
el gen Rb, responsabl e
de l a prote na del
reti nobl astoma, que
acta bl oqueando determi nados
f actores decreci mi ento
e
i mpi dequel a
cl ul a pase del a etapa
Gl a l a etapa S
(Kobayashi , 1996) yl a
f osf oprote na p53, que
acta corno f actor de
transcri pci n sobre el gen
responsabl e de l a
prote na p21, l a cual , a
su vez, i nacti va l a f ormaci n
del
compl ej o
ci cl i na G1- cdk. Adems,
l a prote na p53 ti ene una
probada
capaci dad apoptti ca
(Choi sy- Rossi et al . , 1998;
Meri no y Cordero- Campana,
1998) . De
hecho, tanto l os
protooncogenes como l os
genes supresores de
tumores, j unto a
otros responsabl es de l a
puesta en marcha del propi o
proceso dedestrucci n cel ul ar, son
l os encargados deregul ar l os
mecani smos
apoptti cos.
De
f orma muygeneral , yen rel aci n
conl o anteri or, parece
queexi sten
dos
grupos degenes responsabl es
del a acti vaci n o represi n
del programa de
apoptosi s: l os genes
promuertey l os genes provi da
. Los pri meros codi f i can
productos
queacti van l a apoptosi s a travs del a
i nducci n deunconj unto
de
genes como bax, bad, bak, bi k,
nucl , mec y el si stema
f as/ f asL (Meri no y
Cordero- Campana,
1998; J acotot et al . ,
1999; Nagata, 1999) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I ntroducci n
Concretamente,
se ti ene evi denci a de l a actuaci n de fas en l os
mecani smos de desencadenami ento de muerte cel ul ar presentes en di ferentes
si tuaci ones,
como
en l os procesos neuro- degenerati vos ( Nagata, 1999) . Entre
l os genes promuerte tambi n se encuentran l os
responsabl es de
l as
endonucl easas ( que
rompen el ADN) o l os de l as caspasas, ci stei nprotei nasas
cal ci odependi entes encargadas de l a ej ecuci n di recta del programa de
destrucci n cel ul ar ( Marks yBerg,
1999) .
Tambi n
sei ncl uye dentro del grupo
de
l os
genes promuerte,
a l os genes supresores de tumores, que i nhi ben l a
prol i feraci n cel ul ar.
Los genes provi da son responsabl es de
i nacti var l os si stemas de
destrucci n cel ul ar,
bi en a travs de l a i nhi bi ci n de caspasas y
endonucl easas, o bi en acti vando si stemas de
protecci n cel ul ar ( por ej empl o
l os enzi mas
anti oxi dantes) . Tambi n a l os protooncogenes myc, fos yj un,
i nductores de prol i feraci n cel ul ar, se l es consi dera
genes provi da. Adems,
genes como survi vi n,
nai p ( Marks yBerg, 1999), yotros comoced- 9, bcl - 2 ybcl -
xL
son di rectamente responsabl es de i nhi bi r el
programa de muerte cel ul ar
( J acotot et al . , 1999) .
Desde el punto de vi sta
del presente trabaj o, l o esenci al es que l a
apoptosi s
es i nduci da por unagran vari edad de est mul os de entre l os que
destaca el estrs oxi dati vo( Cl utton, 1997) .
Cuando aumentan l as concentra
ci ones de radi cal es
l i bres, l a cl ul a sufre di ferentes cambi os bi oqu mi cos y
fi si ol gi cos como l a l i beraci n de coenzi mas yun
agotami ento o i nacti vaci n
de l os si stemas
de defensa y reparaci n cel ul ar . Si no se restabl ece el
adecuado equi l i bri o estrs oxi dati vo/ anti oxi dantes,
se desencadenan di fe-
rentes procesos
patol gi cos como peroxi daci n l i p di ca, prdi da de l a
homeostasi s del
cal ci o yunaal teraci n de l as rutas metabl i cas quel l evan a
l a el i mi naci ndel a cl ul a por apoptosi s ( Sl ater, 1987)
.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I ntroducci n
5. Patol og as
rel aci onadas conel estrs oxi dati vo
Durante l os
l ti mos 40aos se ha i mpl i cado
el estrs oxi dati vo en un
gran nmero de enf ermedades,
que, como se i ndi ca en l a Tabl a
3, i ncl uyen
patol og as que af ectan
al si stema nervi oso central ,
ateroscl erosi s, y una
vari edad de procesos i nf l amatori os
y enf ermedades
degenerati vas asoci adas
conel envej eci mi ento
o f actores noci vos del esti l o
devi da, como el tabaqui smo
yel consumo
excesi vo deal cohol
(Marti nez- Cayuel a, 1995) .
S ndromesdegenerati vos
queaf ectanal S. N. C.
* Al zhei mer.
*
Parki nson
Procesos
i nf l amatori os.
* Artri ti s
reumatoi de
* Asma
* Col i ti s ul cerosa
* Gl omerul onef ri ti s
* Gota
* Pancreati ti s
* Uve ti s
Al teraci ones causadas por
si ones anormal es de
ox geno
*Congel aci n
*
Estrs por ej erci ci o f si co
* Fi bropl asi aretrol ental
* Hi poxi a
*
I squemi a- reperf usi n
* Ul ceraduodenal
ten-
Al teraci ones
cromosmi cas y
cncer.
* Def i ci enci as
enzi mti cas
* Enf ermedadde
Batten
* Mel anoma
*
Xerodermapi gmentosa
Otros
s ndromes
Ateroescl erosi s
I nmunodef i ci enci a
Enf ermedades autoi nmunes
Cataratas
Escl erosi s
ml ti pl e
Hi pertensi n
S ndrome hemol ti co del prematuro
Traumati smo
cerebral
Reacci ones txi cas
* Toxi ci dad deanti bi ti cos
*
Toxi ci dad del tabaco y el al cohol
*Toxi ci dad de
xenobi ti cos
Tabl a3. Patol og as rel ac onadas conel
estrs ox dati vo.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I ntroducci n
6. El envej eci mi ento
bi ol gi co
Probabl emente, el fenmenobi ol gi co msi mportante l i gado al estrs
oxi dati vo es el envej eci mi ento, un
proceso que si empre ha provocado gran
i nters
ydado
l ugar
aespecul aci ones sobre sus causasysobre l os factores
quefi j an unl mi te al a duraci n del a vi da de l os ani mal esydel oshumanos.
Conel progreso de l as ci enci as experi mental es en el
si gl o
XI X
empi ezan a
surgi r teor as gerontol gi cas, con
expl i caci ones parci al es de l os mecani smos
del envej eci mi ento, centrndoseencadapocaenl os aspectos
de este proceso
rel aci onados conl os avancescontemporneos
de l a bi ol og a yde l a medi ci na.
As , l a
i nvesti gaci n se hai do despl azando desde el anl i si s estad sti co
de
l a
l ongevi dad de l as pobl aci ones yl a prdi da
de rendi mi ento de l os si stemas
fi si ol gi cos al presente
auge de l a gerontol og a cel ul ar ymol ecul ar (Mi quel
et
al . , 1984, Mi quel yFl emi ng1986, Mi quel , 1990) .
Puesto que el envej eci mi ento ti ene l ugar
a todos l os ni vel es de
organi zaci n bi ol gi ca,
no puede sorprender que se hayan
propuesto
numerosasteor as gerontol gi cas para expl i car l as
al teraci ones que ocurrena
l os di versos ni vel es estudi ados. Segn
Medvedev (1990), se han publ i cado
cerca
de 300 teor as
del
envej eci mi ento, aunquel a mayor a ya sl o
ti enen
un
val or hi stri co, e i ncl uso conceptos tan popul ares
hace unos aos, como l a
teor a del
"error catastrfi co" enl a s ntesi s de prote nas,
de
Orgel
(1963), yl a
del "rel oj mi tti co' ; de Hayfl i ck (1965),
pi erden apoyo. Por el contrari o, l as
i deas sobre
el
papel
que j uegan en el envej eci mi ento l os radi cal es l i bres
(Harman,
1956) yl a "toxi ci dadresi dual " del ox geno
(Gerschman, 1962) ganan
un grado creci ente de aceptaci n, aunque por
s sol as son i ncapaces de
expl i car todos l os efectos
del envej eci mi ento y no ofrecen una expl i caci n
razonabl ede por qu apareci eronenl a bi osfera l os
organi smosqueenvej ecen,
encontrasteconl a aparentei nmortal i daddeformas
devi da mssi mpl e, como
l asbacteri as
yl as amebas.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I ntroducci n
Por el l o, l a opi ni nms
preval ente entre l os gerontl ogos es
que
an
no
se di spone
de todasl aspi ezas del puzzl e del
envej eci mi ento yque ste nose
resol verhasta que aparezca una teor a
general que i ntegre todosl os cambi os
que ocurrenal envej ecer enl os
di versosti pos cel ul ares que forman
l ostej i dos.
Aeste fi n puede
contri bui r una mej or comprensi n
de l os factores que
contri buyena desencadenar el
procesode envej eci mi ento, como etapafi nal
no
di rectamente programada
pero i nevi tabl e del desarrol l o
. Aconti nuaci n
resumi remos al gunos de estos factores y
l uego revi saremos brevemente
l as
teor asgerontol gi cas
que ahora encuentranuna
mayor aceptaci n.
6. 1 . Factores
que i nfl uyenenel
envej eci mi ento.
Factores de creci mi ento
. La fal ta o escasez de
determi nados factores de
creci mi ento puede
desencadenar l a apoptosi s
l i gada a l a senescenci a.
Adems, en
el caso de l as neuronas se
ha comprobado que aquel l as que
no
l l egana al canzar susdi anas
o noreal i zanl os contactos
adecuadosdurante el
desarrol l o se
"sui ci dan" por apoptosi s
(Cl arke et al . , 1998) .
Posi bl emente l a
fal ta de adecuada
modul aci n por parte
de determi nados factores
de
creci mi ento
(compl ej os ci cl i nas)
es uno de l os moti vos que
conducen a l a
apoptosi s. Adems l a fal ta de
i nducci n de l os genes
responsabl es de l a
s ntesi s de determi nados
enzi masanti estrs
(Tagl i al atel a et al . , 1998) as
como
de
prote nas secuestradoras de
cal ci o (como l a cal bi ndi na)
pueden tambi n
contri bui r al desarrol l o de
l a apoptosi s (Li pton y
Kal i l , 1995) . Ytambi n, l a
fal ta de repres n
de l a maqui nar a de
muerte cel ul ar por parte de
determi nados factores trfi cos puede
ser otro de l os mecani smos
i mpl i cados,
dado que l os pri nci pal es
enzi mas que
desencadenanl a apoptosi s (caspasasy
endonucl easas), suscepti bl es
de acti vaci n, estn
presentes en todas l as
cl ul as (Boonman
yI sacson, 1999) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I ntroducci n
Def i ci enci as en el
sumi ni stro energti co. Se sabe que el requeri mi ento
de energ a de l as neuronas est muypor enci ma del
de l as otras cl ul as del
organi smo. Esto se debe a l a necesi dadde
sumi ni strar ATPconti nuamente a
l a bomba
de
Na+/ K+
de su membrana cel ul ar, de cuyo f unci onami ento
dependentanto l a homeostasi s i ni ca comoel manteni mi entodel potenci al de
membrana, de sumai mportanci a enl a f i si ol og a neuronal
.
De hecho, ms
del 40%del ATPproduci do por l as mi tocondri as
neuronal es se desti na aeste f i n (Ames et al . , 1992) y
por el l o l os sustratos del
proceso de f osf ori l aci n
oxi dati va (sobre todo l a gl ucosa yel ox geno) deben
estar di sponi bl es en abundanci a en l a
neurona. Por otra parte, l a
hi pogl ucemi a e hi poxi a asoci adas
a l os procesos de i squemi a causan una
i nhi bi ci n
de
l a
f osf ori l aci noxi dati va y, por l o tanto de l a
producci nde ATP,
con l a consi gui ente prdi da de f unci n que a
menudo acompaa al
envej eci mi ento cerebral (Davi s et al . ,
1997) .
Exci totoxi ci dad
. Son ml ti pl es l os f actores que
desencadenan l a
exci totoxi ci dad. As , se ha observadoque l a
prdi da de homeostasi s i ni ca, l a
depl eci n de reservas
energti cas, l a i squemi a, l as l esi ones traumti cas, l as
enf ermedades
neurodegenerati vas y el envej eci mi ento
provocan despol a-
ri zaci n exci tatori a y, por
consi gui ente, l i beraci n de neurotransmi sores
exci tadores (Ol ney, 1994) .
Se ha i mpl i cado
f undamental mente al receptor NMDA como
responsabl e de l a puesta enmarcha
del procesode toxi ci dad que puede tener
l ugar medi ante
dos v as di f erentes (Auer, 1991) . La
pri mera de el l as,
denomi nada de "exci totoxi ci dad aguda",
es provocada por agoni stas
gl utami nrgi cos yse asoci a conl a
entrada masi va de Na+, Cl yH2O en l a
cl ul a, con
aumento del vol umencel ul ar yconsi gui ente
edema exci totxi co
que puede l l evar a l a l i si s
osmti ca de l a cl ul a y, por consi gui ente, a
su
muerte
necrti ca.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I ntroducci n
Por otra parte, en l a
"exci totoxi ci dad retardada"
i ntervi ene una
el evaci ndel cal ci o ci tosl i co, que asuvez
acti va unaseri e de enzi mas cal ci o-
dependi entes como l a f osf ol i pasa
A2, protei nasas, endonucl easas
y xi do
n tri co si ntasa
( Strol i n- Benedetti et al . , 1993)
. Todos estos enzi mas
desencadenan
cascadas metabl i cas que
enl ti ma i nstanci a pueden
provocar
daoymuerte cel ul ar por apoptosi s
. Adems, se haobservado
que el proceso
da l ugar a una i nhi bi ci n
de l a s ntesi s de prote nas
yque tambi n ti ene un
i mportante
papel patogenti co el aumento
enl a producci nde
radi cal es l i bres
( Schonei ch, 1999) .
Prdi da de l a homeostasi s del
cal ci o. El aumentode
l os ni vel es de Cae+
ci tosl i co acti va determi nados
enzi mas Cae+- dependi entes
como son l as
f osf ol i pasas,
proteasas, endonucl easas yl a
propi a NOS, todas el l as
i mpl i cadas
en l a
ej ecuci n del programa de
muerte cel ul ar
( Chakraborti et al . , 1999) .
Estos aumentos
de l a concentraci n de
Cae+- ci tosl i co son causados
por el
i nf l uj o de
Cae+- extraci tosl i co que
penetra por canal es acopl ados
areceptores
regul ados por vol taj e o agoni stas,
como es el caso de
l os receptores de
gl utamato o NMDA
( Mi l l s, 1996) . Cuando
el Cae+ penetra en l a
cl ul a, sta
ti ende a estabi l i zar el aumento
de su concentraci n
medi ante captura
y
al macenami ento
del cati n en depsi tos
i ntracel ul ares tal es como
el ret cul o
endopl asmti co yl os cal ci osomas,
uti l i zando para el l o
bombas metabl i cas
dependi entes de
ATP. La mi tocondri a
tambi n al macena cal ci o
medi ante un
transportador
de ti po uni portador
dependi ente de vol taj e,
de sumembrana. No
obstante, l a captura de
cal ci o por l os orgnul os
ci tados puede ser i nef i caz si
persi stenl os
mecani smos que causansuentrada
enl a cl ul a.
Por
ej empl o, l a uni nde gl utamato
aNMDAno sl oprovoca
l a apertura
de canal es de cal ci o si no
que tambi nacti va l a
f osf ol i pasa Cde l a membrana,
que
transf orma el i nosi tol bi f osf ato
en i nosi tol tri f osf ato y
di aci l gl i cerol . El
i nosi tol tri f osf ato
abre canal es de cal ci o en el
ret cul o endopl asmti co, y
el
mi smo
aumento de l os ni vel es de cal ci o
causa l a apertura de
canal es enl os
cal ci osomas, l o que l l eva
a unf l uj o masi vo del cati n
desde sus depsi tos
i ntracel ul ares
haci a el ci tosol ( Pascheny
Douthei l , 1999) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I ntroducci n
Si nembargo, no son stas l as ni cas causas del os al tos ni vel es de Cae+
ci tosl i co, pues l a mi tocondri a tambi n desempea un i mportante
papel
(Rusti n et al . , 2000) . As ,
como ya se hamenci onado, este orgnul o tambi n
acta como reservori o de Cae+, el cual penetra en su i nteri or graci as aun
transportador que f unci ona i ni nterrumpi damente mi entras se mantenga
el
potenci al de membrana
mi tocondri al . El Cae+ ci tosl i co que entra en l a
mi tocondri a es uti l i zado para l a acti vaci n de determi nadas
deshi drogenasas
del ci cl o de l os ci dos
tri carbox l i cos, y su sal i da de l a mi tocondri a (por un
mecani smo de cotransporte) permi te l a i ntroducci n si mul tnea
de Na+desde
el ci topl asma al i nteri or del orgnul o. A
su vez, el Na+mi tocondri al es
cotransportado de
nuevo al hi al opl asma permi ti endo l a entrada de H+al
i nteri or mi tocondri al . Se crea, de esta f orma, un gradi ente
de H+que resul ta
i mpresci ndi bl e para que puedan darse l as condi ci ones
que permi ten el
desarrol l o del proceso de
f osf ori l aci n oxi dati va, es deci r, de generaci n de
energ a en f ormade ATP. As , es sabi do que una
ATPsi ntasa presente en l a
membrana mi tocondri al
uti l i za l a energ a al macenada en el gradi ente de
protones para generar ATP. Si n embargo, todo
este si stema puede
desequi l i brarse como
consecuenci a de l a prdi da del potenci al de l a
membrana
mi tocondri al (Sugrue et al . , 1999) . Laprdi dade potenci al
se debe
f undamental mente a que en l a mi tocondri a puede abri rse,
de f orma ms o
menos transi tori a, un poro que
de f orma i nespec f i ca dej a sal i r haci a el
hi al opl asma i ones y
mol cul as i nf eri ores a 1, 5 kDa, entre el l os Cae+. De esta
f orma se di si pa el potenci al de membranay l a
ATPsi ntetasa puede tambi n
f unci onar a l a i nversa
consumi endo ATPen l ugar de generarl o. Aunque
se
especul a con l as causas queprovocan l a apertura del
poro mi tocondri al , se
conocenyaal gunos aspectos
i ndi cati vos del posi bl emecani smoregul ador. As ,
se ha
comprobado que el i nmunosupresor ci cl ospori na A i mpi de
l a apertura
del poro, mi entras que, por el contrari o,
el evadas concentraci ones de Cae+
ci tosl i co as como
el estrs oxi dati vo acti van su apertura (Cassari no y
Bennett,
1999) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
6. 2. Teor as sobrel as causas
del envej eci mi ento normal .
I ntroducci n
Laprdi da del potenci al
de membranami tocondri al no sl o i mpl i caun
descenso enl a producci n de energ a, si no
quetambi nl l eva al a prdi da de
l a homeostasi s cel ul ar, aumento
de l os ni vel es de Cae- 1 ci tosl i co y
resul tante
autoal i mentaci n
de un proceso que acaba
provocando l a muerte cel ul ar
necrti ca
oapoptti ca.
6. 2
. 1
.
Teor a del "envenenami ento" por l os
radi cal es l i bres de
ox geno
Esta teor a f ue
propuesta por Rebeca
Gerschman, a parti r de sus
estudi os
sobre el aumento de l os
ef ectos l esi vos de l a radi aci n
i oni zante en
presenci ade ox genoy
el ef ecto protector del a
f al tadeestegas (Gerschman
et
al . , 1954) .
Segnestaautora,
el ef ecto si nerg sti co de l a radi aci n
yel ox geno
(oxygen ef f ect) se debe
al aumento que causa este
gas en l a concentraci n
i ntracel ul ar de
radi cal es de ox geno,
quesesumanal os radi cal es
quederi van
de l a
i rradi aci n. Esto l a l l ev
a deduci r que el ox geno
que se usaen el
metabol i smo desencadena
el envej eci mi ento
(a travs de l a f ormaci n de
radi cal es l i bres)
debi do a quel as def ensas
anti oxi dantes del organi smo
no
contrarrestantotal menteel
ataque oxi dati vo.
6
. 2. 2. Teor adel os radi cal es
l i bres
Segnesta teor a,
muysi mi l ar a l a de
Gerschman, l os radi cal es l i bres
no sl o pueden
causar el
envej eci mi ento si no tambi n otros
procesos
degenerati vos, como
el cncer, l a arteri oscl erosi s,
l a ami l oi dosi s y l a
i nmunodef i ci enci a
. Segn Harman (1956),
tanto l a senescenci a como estos
s ndromes
sonel resul tado de l as reacci ones
noci vas de l os radi cal es
l i bres
queti enenl ugar conti nuamente
atravs del as cl ul asydel os
tej i dos.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I ntroducci n
Adems, Harman i mpl i ca en su teor a
de
l os radi cal es l i bres al a
evol uci n
afi rmando que "segn parece, l a vi da surgi como resul tado de
reacci ones de radi cal es l i bres, sel ecci on reacci ones
de
radi cal es l i bres para
j ugar un
papel
en
el metabol i smo yasegur l a evol uci n empl eando estas
reacci ones paracausar mutaci ones yl amuerte" (Harman, 1981) .
6
. 2 . 3
. Teor a
mi tocondri al del envej eci mi ento
Estateor a propuestapor Mi quel et al . en 1980 l i ga el envej eci mi ento
cel ul ar aun dao produci do por
el estrs oxi dati vo a
ni vel
mi tocondri al . Se
tratadeun
concepto dedao genti co/ bi oenergti co, pues l os autores afi rman
que "el envej eci mi ento cel ul ar est l i gado al esi ones del genomami tocondri al
por radi cal es l i bres queseforman en l a
cadenarespi ratori ade
l a
membrana
mi tocondri al
i nterna"
.
Acausa del entorno mutagni co yrel ati va fal ta de
protecci n del genomami tocondri al
(quecarecedehi stonas ydemecani smos
de
reparaci n por esci si n yrecombi naci n), l as mi tocondri as pi erden
con el
paso del ti empo su i nformaci n,
l o que i mpi de l a renovaci n de estas
organel as yl l evaasu
desorgani zaci n yconsi gui ente prdi dade s ntesi s de
ATPydefunci n
fi si ol gi ca(Mi quel , 1992) .
Esta teor a es compati bl e con
l os ms i mportantes conceptos
gerontol gi cos como el del
"desgaste de l as cl ul as somti cas como efecto
secundari o
de su trabaj o fi si ol gi co" (Wei ssman, 1891), l a teor a
del
envej eci mi ento como "efecto secundari o"
de l a di ferenci aci n cel ul ar (Mi not,
1907), as
como el concepto antes resumi do de l a "toxi ci dad
resi dual del
ox geno" (Gerschman, 1954) y
l a teor a de Harman (1956), que defi ne al
envej eci mi ento
como
l a
"sumade l as l esi ones resul tantes del ataque de l os
radi cal es l i bres queseproducenconstantemente
enel organi smo".
Segn Mi quel (1992), al
centrarse en el genoma mi tocondri al de l as
cl ul as di ferenci adas
como l a di ana ms probabl e de l os oxi - radi cal es
deri vados del metabol i smoaerobi o, l ateor ami tocondri al puede
i ntegrar todos
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I ntroducci n
l os conceptos
gerontol gi cos menci onados
anteri ormente, para of recer una
expl i caci nrazonabl e de l as
al teraci ones queocurrenal envej ecer a
todos
l os
ni vel es de organi zaci n
bi ol gi ca.
6. 2. 4. Teor ade l amutaci n
genti ca
Lacl ave para
entender el proceso de
envej eci mi ento desde el punto de
vi sta
de
l a
evol uci nbi ol gi ca es
encontrar unmecani smoqueexpl i que
cmo
y por qu se desencadenan
l os cambi os
f unci onal es, estructural es y
mol ecul ares que
ocurrenconel pasodel
ti empo.
Deacuerdoconl ateor adel
" rel oj mi tti co" ( Hayf l i ck, 1965) , el
l mi te de
l a
capaci dad de di vi si n cel ul ar
podr a depender
de l a prdi da de genes que
i nducen, como
f os, l a prol i f eraci n
de l as cl ul as ( protooncogenes)
. Segn
trabaj os ms reci entes,
se consi dera que l a
senescenci a deri va de un
programa
genti coqueempi ezaaej ecutarse
conel desarrol l o ( etapa
prenatal y
postnatal ) , conti naen
el adul toycul mi na, tras l a
etapareproductora, en l a
vej ez ( Fi nch,
1990; J ohnson, 1997; Ki rkwood,
1998) . De hecho,
empi ezan
a
conocerse toda unaseri e
de genes, denomi nados
gerontogenes, queguardan
unaestrecha
rel aci n con l a esperanza de vi da de
l os ani mal es de di sti ntas
especi es ( Rose,
1994; Longo, 1999;
Gems, 1999) . Laacti vi dad de estos
genes
estmodul adapor numerosos
est mul os, tanto externos
comoi nternos, que
actan
desencadenando cascadas de
seal es que i nducen o repri men su
expresi n yque, por
consi gui ente, acel eran o retrasan
el desarrol l o del
programa genti co
que conduce al a
senescenci a. Se ti ene conoci mi ento de
que al gunos de estos genes
estn mutados enl os mi embros
de f ami l i as que
padecen
enf ermedades degenerati vas
como l a progeri a yl a enf ermedad de
Al zhei mer ( Wel sh, 1975; Mi l l s y
Wei l l s, 1990; Prez- Tur, 2000) . Si nembargo,
aunque es evi dente
que l os genes estn i mpl i cados
en patol og as
neurodegenerati vas,
an se desconocen
sus mecani smos espec f i cos de
actuaci n
conrespecto al envej eci mi ento
.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I ntroducci n
Por otra parte, exi ste
una seri e de genes con una i mpl i caci n ms
di recta en l os fenmenos de senescenci a normal
ymuerte cel ul ar, cuya
funci n yl a de
l os productos que codi fi can se conocen mej or graci as a l os
estudi os real i zados en model os de envej eci mi ento
ms senci l l os como l a
l evadura Saccharomyces
cerevi si ae ( J azwi nski , 1999) , el pequeo nematodo
del suel o Caenorhabdi ti s
el egans ( J ohnson, 1997; Vanfl eteren y
Braeckman,
1999) yl a mosca de l a fruta Drosophi l a
mel anogaster ( Rose, 1999) . Son
especi al mente i nteresantes
l os trabaj os del grupo de Horvi tz ( 1999) en
Caenorhabdi ti s el egans
. Esta i nvesti gaci n demostr que durante el
desarrol l o
de este organi smo "se sui ci dan" 132 cl ul as
de l as 1090que ti ene l a l arva, y
que el mecani smo de
muerte apoptti ca tambi nopera en
procesos fi si ol gi cos
de recambi o cel ul ar en l os tej i dos del
organi smo adul to. Como ya hemos
i ndi cado, esta muerte
cel ul ar apoptti ca estgenti camente
programada, a
di ferenci a
de l a muerte cel ul ar necrti ca,
no programada o patol gi ca
( Horvi tz, 1999) .
6. 2. 5. Rel evanci a
gerontol gi ca de l os estudi os sobre
tel meros y
tel omerasa
Desde l os aos 30se sabe
quel os cromosomas nose fusi onan conotros
cromosomas rotos
graci as a l a presenci a de una estructura protectora
al fi nal
del cromosoma, denomi nada tel mero
( Mul l er, 1938) . Ms reci entemente se
demostr que l os tel meros
son repeti ci ones en tndem
de l a secuenci a
TTAGGGyprote nas asoci adas
( Bl ackburn, 1991) , as como el si ti o de uni n
de l as prote nas que reparan
roturas de l a dobl e hl i ce del ADN( Bi anchi
yDe
Lange, 1999; Hsuet al . ,
1999) .
Aunque
l a rel evanci a gerontol gi ca
de l os datos sobre tel meros y
tel omerasa es obj eto de controversi a,
l o ms rel evante desde el punto
de vi sta
de l a senescenci a
es que cada vez que l a cl ul a
se di vi de, l os tel meros se
acortan con l a consi gui ente prdi da de i nformaci n
genti ca que resi de en l as
secuenci as repeti das
TTAGGG( Gems, 1999) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I ntroducci n
Este acortami ento
noocurre encl ul as comol as germi nal es o l as cancerosas
graci as a
l a presenci a enl as mi smas de l a tel omerasa,
unenzi maque repone
l as zonas
de ADNperdi das y
conf i ere
a
l as cl ul as que l a poseenun
f enoti po
i nmortal (Campi si , 1997) .
El envej eci mi ento tampocoti ene
l ugar enespeci es
en l as que
todas l as cl ul as de sus suj etos
adul tos (i ncl ui das l as neuronas)
estn someti das a
unconti nuo recambi o, como ocurre por
ej empl o en l as
hi dras de
aguadul ce (Mart nez,
1998) .
La prdi da de
f unci n tel omri ca que ocurre en
l as cl ul as de l os
humanos, al
envej ecer, puede deberse a
una di smi nuci n progresi va de
l as
secuenci as repeti das TTAGGG
con el creci ente nmerode mi tosi s
que ti enen
l ugar conel paso del
ti empo, o auna prdi da o mutaci n
de l as prote nas
tel omri cas (Van
Steensel et al . , 1998;
Samper et al . , 2000; Goyti sol o et
al . ,
2001) . En
todo caso l a prdi da
de f unci n tel omri ca da l ugar
a f usi ones
cromosmi cas
(Bl asco et al . , 1997; Hande
et
al
. , 1999), di f i cul tades en l a
separaci n de l os puentes de
anaf ase e i nestabi l i dad cromosmi ca
general ,
que causal a prdi da
de vi abi l i dad cel ul ar y
puede tener uni mportante
papel
no
sl oenel envej eci mi ento
si notambi nenel cncer y
reparaci nde l esi ones
del ADN(Autexi er y
Grei der. , 1996; Goyti sol o et al . ,
2000; Wonget al . , 2000) .
A
pesar del i ndudabl e i nters de l os
mecani smos rel aci onados
con l os
tel meros yl a tel omerasa
enl os procesos de mi tosi s y
di f erenci aci n cel ul ar,
Carl sonyRi l ey
(1998) af i rmanquenoest
demostradoqueestos mecani smos
(i nvesti gados
encl ul as manteni das
encul ti vos cel ul ares) se rel aci onen
conel
envej eci mi ento del
organi smo, ya que i ncl uso en
tej i dos con al to
reaprovechami ento,
l os mam f eros
no muerencomoresul tado del
agotami ento
de su potenci al mi tti co.
De f orma si mi l ar, opi nan Goyns
yLavery (2000) :
"Muchas
cl ul as del cuerpo no
prol i f eran regul armente ypor tanto
no se
acortansus tel meros
. . . Nohaycorrel aci n entre l a l ongi tud de
l os tel meros
yl a l ongevi dad
mxi made l as di sti ntas especi es
. . . Paraval orar l a i mportanci a
de l a
tel omerasa en el proceso del
envej eci mi ento es esenci al consi derar
l a
si tuaci n i nvi vo, que es ms
rel evante que cul ti var f i brobl astos
enpl acas de
Petri ".
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I ntroducci n
Deacuerdo con estas opi ni ones, l os estudi os sobrel os tel meros yl a
tel omerasa no restan
rel evanci a a l as opi ni ones de Mi not (1907) sobre l a
estrecha
rel aci n entredi f erenci aci n yenvej eci mi ento. Por el l o actual mente
se presta gran atenci n al estudi o de l os mecani smos del envej eci mi ento i n
vi vo, especi al menteel del as neuronas yotras cl ul as di f erenci adas
(Mi quel ,
2002)
.
6. 3. Conceptoi ntegrador deteor as cl si cas ymodernas
Segn hemos resumi do,
se
han
propuesto di versas teor as di ri gi das
f undamental mente a expl i car l os cambi os que ocurren al envej ecer a
l os
di sti ntos ni vel es de
organi zaci n (de l a pobl aci n a l as macromol cul as) .
Puesto queni ngunateor a proporci ona unaexpl i caci n sati sf actori a de
todos
l os aspectos del envej eci mi ento, l a tendenci a actual es l a de
uni f i car l os
conceptos cl ave en una teor a i ntegradora
. Como ya hemos comentado,
numerosos
datos apoyanel concepto dequeel agentepatogni co es el ataque
por l os radi cal es deox geno yl a pri nci pal di anadedi cho ataque
es el genoma
mi tocondri al del as cl ul as di f erenci adas
. SegnMi quel (1998), esteconcepto
ayuda ai ntegrar
l as teor as cl si cas (quedestacaban el papel desorgani zador
del metabol i smo aerobi o yl a
prdi da decapaci dad regenerati va l i gada a l a
di f erenci aci n
cel ul ar) con el presente i nters en el estrs oxi dati vo y sus
ef ectos sobrel ai nf ormaci ngenti canucl ear y
mi tocondri al .
En resumen,
desde el punto de vi sta de l a evol uci n bi ol gi ca
el
envej eci mi ento
es el resul tadodel a apari ci ndecl ul as, comol a
neurona, que
por su al to consumo deox geno ynul a
o escasa capaci dad deregeneraci n
i ntegral (a travs de l a
mi tosi s) acumul an l esi ones peroxi dati vas que
di smi nuyen
su capaci dadbi oenergti cayf unci onal ysonpor
el l o el "tal n de
Aqui l es" de l os organi smos
mul ti cel ul ares ( . . . ) Las cl ul as que f orman
l os
tej i dos somti cos
pri mero si guen un programa genti co de
di f erenci aci n y
l uego se desorgani zan como resul tado
del desgaste por estrs oxi dati vo
mi tocondri al y de su
prdi da de mecani smos de regeneraci n
subcel ul ar
i ntegral "
(Mi quel , 1998) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I ntroducci n
7.
Pri nci pal es
enfermedades ocul ares asoci adas con el estrs
oxi dati voyel
envej eci mi ento
El envej eci mi ento
ocul ar se acompaa de
numerosos cambi os ani vel
funci onal , bi oqu mi co
yestructural . Entre
l os msrel evantes seencuentran
l a
prdi da
de transparenci a del
cri stal i no, con el consi gui ente
descenso de l a
capaci dad de acomodaci n
yde l a agudeza
vi sual , proceso denomi nado
cataratas ( Fal t,
1992) . Durante vari as
dcadas se han estudi ado
l as
al teraci ones bi oqu mi cas
rel aci onadas con l a formaci n
decataratas l l egando
a l a
concl usi n de que l a
cataratognesi s es un
proceso compl i cado y
mul ti factori al , nti mamente
l i gado al envej eci mi entoy
probabl ementei ni ci ado
por el estrs oxi dati vo
( Berman, 1991) .
Tambi nparecej ugar un papel
muy
i mportante el estrs oxi dati vo
en l a patognesi s
de l a degeneraci n macul ar
asoci ada al aedad ( DMAE)
. Enel presente estudi o
nos hemos centrado en
un
model o rel evante
paraacl arar
mecani smos patogenti cos l i gados al a
DMAE,
un
proceso de grani mportanci a
por l a amenazaque
suponeal a cal i dad de
vi dadel as personas
mayores, del que
trataremos brevementea
conti nuaci n.
7. 1.
Degeneraci nmacul ar
asoci adaal a edad
( DMAE)
Puesto que
el rea macul ar del a
reti na es responsabl e
del a vi si n de
al ta
resol uci n ydel a
vi si n en col or, l as al teraci ones
de este rea l l evan
a
i mportantes prdi das
devi si nei ncl uso
al a ceguerapermanente.
Las
l esi ones degenerati vas
de l a mcul a que ocurren
al envej ecer
pueden ser secundari as
a cambi os vascul ares
. El proceso bsi co
probabl emente est
rel aci onado con
al teraci ones en l a permeabi l i dad
de l os
capi l ares
reti ni anos o de
l os cori ocapi l ares ( debi do
a una exudaci n
subreti ni ana e
i ntrareti ni ana), o conuna estenosi s
de l os pequeos vasos
adyacentes
al a mcul a, con
resul tanteatrofi adel EPR
( Kl ei n et al . , 1989) .
Por otra
parte, l a degeneraci n
progresi va del EPRes el factor
ms
deci si vo
en el desarrol l o del a
enfermedad, debi do asuestrecha
rel aci n con
l os fotorreceptores, tanto
enel aspectonutri ci onal como
metabl i co, por l o que
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I ntroducci n
l a al teraci n progresi va del EPRtrae consi go l a degeneraci n progresi va de
conos ybastones, esenci al es para l a f unci nvi sual
( Zarbi n, 1998;
Adl er
et al . ,
1999) .
8. Model os para el
estudi odel estrs oxi dati voyenvej eci mi ento.
8. 1. Epi tel i o pi gmentari o de reti na como model o de envej eci mi ento
cel ul ar i nvi tro
Los estudi os i nvi vo sobre l a DMAEsondi f i ci l es
ycostosos,
porque
l os
ni cos ani mal es de
experi mentaci nque poseenuna mcul a bi endesarrol l ada
son
l os pri mates . Por el l o, l os cul ti vos i n vi tro de EPRson consi derados
i dneos para l a real i zaci nde i nvesti gaci ones para
l a i nducci nde
dao
por
estrs oxi dati voyenvej eci mi ento
cel ul ar ( Matsunaga et al . , 1999; Ca et al . ,
2000) . Para l a
mej or comprensi nde este model oi nvi tro, quees el que hemos
uti l i zado en el presente estudi o, resumi mos
a conti nuaci n sus pri nci pal es
caracter sti cas hi stol gi cas y
f i si ol gi cas .
El EPRes una monocapa de cl ul as epi tel i al es
conf orma hexagonal de
ori gen neuroectodrmi co
que se encuentra si tuadoentre l a redcori ocapi l ar y
l a capa de f otorreceptores de l a reti na. La presenci a
de esta capa cel ul ar enel
oj oj uega unpapel cruci al en l a
preservaci n del si stema vi sual ( Berman,
1991), ya que consti tuye
j untoconl os capi l ares de l a reti na neural , l a barrera
hemato- reti ni ana
( Wi sl ocki yLadman, 1955), desempeando un
i mportante
papel enel control del i ntercambi o
de materi al entre l os capi l ares yl a reti na
neural .
Entre
l as pri nci pal es f unci ones del EPR,
adems del manteni mi entode
l a barrera hemato- reti ni ana
ydel espaci osubreti nal ( Shaki b yCunha- Vaz,
1966), se puede destacar l a necesi dad de
su presenci a durante l a
embri ognesi s para el
ef i caz desarrol l o de l os f otorreceptores ( Mund y
Rodri gues, 1979), el
reconoci mi entoyf agoci tosi s de l os segmentos externos
de
l os
f otorreceptores que hansi dodesechados ( Bok y
Young. , 1979) yel trans-
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I ntroducci n
porte acti vo
de metabol i tos desde yhaci a l a reti na ( Stei nberg y
Mi l l er . 1979) .
Otra f unci n i mportante
del EPRes l a recepci n, transporte, al macenami ento
ymetabol i smo
de l a vi tami na A( RappyAnderson, 1983) .
Se trata de untej i do que
constantemente suf re estrs oxi dati vo ya
que
en l l os radi cal es
l i bres songenerados por di f erentes causas,
entre el l as una
el evada
concentraci n de
02
en l a zona de l a mcul a
( Al der et al . , 1985) as
como
el estar someti do a una i ntensa
i l umi naci n ( Dorey et al . , 1989),
que
desencadena el proceso
de peroxi daci n l i p di ca ( Tate et al . ,
1995) . Por otra
parte, est
expuesto a un adi ci onal estrs
oxi dati vo debi do a su f unci n
de
f agoci tosi s de l os
segmentos externos de l os f otorreceptores
( Kennedy et al . ,
1994) .
Por todo el l o, el EPRpuede
consi derarse como un excel ente
model o
para
el estudi o del papel
de l os radi cal es l i bres de ox geno
enel envej eci mi ento
cel ul ar .
Un i ndi ci o de que l a
DMAEl i gada al estrs
oxi dati vo ef ecti vamente
ocurre en el EPR
es l a acumul aci n del pi gmento
del envej eci mi ento
l i pof usci na, como
resul tado de l a degradaci n
de l os componentes
de l os
segmentos
externos de l os
f otorreceptores que han
si do f agoci tados.
La
l i pof usci na
puede estar rel aci onada con
una di smi nuci n de l a
acti vi dad
l i sosomal
del epi tel i o, as
como con l a degradaci n de
l as membranas
mi tocondri al es
quehansuf ri do l as
consecuenci as crni cas del estrs
oxi dati vo
( Hol z
et al . , 1999) .
8. 2. Ratones OF1
comomodel ode envej eci mi ento
i n vi vo
La mayor a de l os estudi os
de gerontol og a experi mental
i n vi vo se han
real i zado en ratas y
ratones de l aboratori o. Estos
estudi os son ti l es
para
poner a
prueba l as hi ptesi s sobre l os
mecani smos f i si ol gi cos, genti cos
y
bi oqu mi cos asoci ados al envej eci mi ento
normal y patol gi co, as
como l a
ef i caci a de l as
i ntervenci ones experi mental es que se
real i zan para aumentar
l a
l ongevi dad.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I ntroducci n
Uni mportante resul tadode estos estudi os en roedores es queexi sten
di f erenci as
si gni f i cati vas entre l a "edad bi ol gi ca" y l a "cronol gi ca" de
ani mal es de l a mi sma cohorte yque, segnCol l i er yCol eman( 1991) , "estas
di f erenci as aportan datos ti l es para
estudi ar l os mecani smos del
envej eci mi ento normal , envej eci mi ento pti mo, envej eci mi ento anormal y
enf ermedad". Estos autores tambi n comentan
que al gunos roedores vi ej os
muestranval ores de ci ertos parmetros prcti camente i dnti cos a
l os de
l os
ani mal es j venes y, por el l o concl uyen que "l a edad
cronol gi ca, como ni co
parmetro,
es
una
medi dai nsuf i ci ente yamenudoequ voca".
Lo anteri or j usti f i ca l os estudi os en ratones para i nvesti gar l as
rel aci ones entre radi cal es l i bres, edadcronol gi ca y
edadbi ol gi ca, as como
l a uti l i dad
de l os tratami entos para consegui r que un mayor nmero de
ani mal es
muestrenun"envej eci mi entopti mo".
De acuerdo
con estos conceptos, en l as dcadas de l os setenta yl os
ochenta el l aboratori o de Mi quel estandari z mtodos
de val oraci n de l os
tratami entos anti oxi dantes en
ratones macho de l a cepa C57BL/6J . Estos
mtodos
de determi naci n de "edad bi ol gi ca" se basanen
el
anl i si s
de l as
curvas de l ongevi dad, de l os val ores de l a coordi naci n
muscul ar y vi gor
( determi nados medi ante l a
prueba de l a cuerda ti rante; Mi quel yBl asco,
1978) ,
de l a di stri buci nde l os val ores del peso corporal ( Mi guel y
Economos,
1979) ydel consumo di ari o de pi enso ( Mi guel , 1983) .
Mtodos si mi l ares,
adecuados para i nvesti gar l a
reacci n de ratones devari as cepas al a i ngesta
de supl ementos anti oxi dantes, hansi do uti l i zados por
De J uan ( 1994) , De l a
Fuente et al . ( 1998b) ,
Pal l ardet al . ( 1998) , I ngramyJ ucker. ( 1999) y
Correa
et al . ( 1999) .
Una tcni ca compl ementari a
de val oraci n conductual ( estandari zada
enratones hembra
OF1) permi tecl asi f i car al os ratones en"rpi dos"
o"l entos"
segn su acti vi dad expl oratori a espontnea cuando
se l es col oca en un
l aberi ntoenf ormadeT( DeJ uan, 1994) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I ntroducci n
Como sucede con otros
parmetros de rendi mi ento f i si ol gi co,
l a
acti vi dad expl oratori a
espontnea (val orada por
el test del l aberi nto en T)
di smi nuye con l a edad. As ,
mi entras que l a mayor ade l os ratones
j venes
(de. 3- 4 meses de
edad) son capaces de compl etar
l a expl oraci n del
pri mer
brazo
del l aberi nto en f orma
de T en menos de 20 segundos,
con el
envej eci mi ento un nmero
creci ente de ratones
expl oran el l aberi nto muy
l entamente
o i ncl uso permanecen i nmvi l es,
mostrando l a denomi nada
`f reezi ng
behavi or" por Gi l adyGi l ad
(1995) . Estaconductaref l ej a
unaf unci n
def i ci ente de l as reas del
si stema nervi oso central
(SNC) que determi nan l a
respuestaa
f actores ambi ental es estresantes
.
Larespuestai nadecuada
aestos f actores (asoci adacon
un al to gradode
ansi edad) es una
de l as caracter sti cas ms
t pi cas del envej eci mi ento
(Stei n-
Behrens ySpol sky,
1992) , por l o cual se puede
concl ui r quel os ratones que
no pasanl a
prueba del l aberi nto en
20segundos ("ratones l entos"; RL)
estn
prematuramente envej eci dos
enl o querespectaaci ertas f unci ones
de suSNC
en comparaci n con
l os ani mal es de l a mi sma
edad quepasanl a prueba
con
xi to
("ratones rpi dos"; RR) .
Deacuerdo coneste concepto, el
porcentaj e de
supervi venci a aedades
avanzadas es mayor para
l os RR(que compl etan l a
pruebaenun
ti empo menor de20segundos)
queparal os RL.
Segn De l a Fuente
et al . (1998b) , l os RL ti enen
un si stema
i nmunol gi co
menos competente (en cuanto a
l a f unci n de l i nf oci tos
y
macrf agos) quel os RR.
Por otraparte, segn Correa
et al . (1999) , este ef ecto
del
envej eci mi ento sobre l os RL puede
ser contrarrestado por l a
admi ni straci n del anti oxi dante
TP, que no sl o esti mul a l a
acti vi dad
f agoc ti ca si no que
neutral i za el estrs oxi dati vo, a
j uzgar por el descenso en el
ni vel
deradi cal superxi do
extracel ul ar enl as suspensi ones demacrf agos
.
Otro estudi o sobre
l a rel aci n entre envej eci mi ento y
conducta en
ratones BALB/c y
OFI muestra que tanto l os machos
como l as hembras
rpi dos de estas dos cepas ti enen
una mayor l ongevi dad que l os
RL
(Guayerbas et al . ,
2002) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I ntroducci n
Adems, l os RRen l a prueba del l aberi nto ( de l as dos cepas y
sexos)
tambi n
muestran un mej or rendi mi ento que l os RL en l a otra prueba
psi comotora, es deci r l a queval ora l a coordi naci n neuromuscul ar y
el vi gor
enl a
cuerda ti rante segnel mtodo de Mi quel yBl asco ( 1978) .
Msreci entemente, l a uti l i dad del a prueba del l aberi nto en forma
deT
para estudi os gerontol gi cos
ha si do confi rmada por Guayerbas et al . ( 2002) ,
pues segn
l os datos obteni dos enunestudi o l ongi tudi nal del envej eci mi ento
deratones hembra OF1 el baj o rendi mi ento en el l aberi nto del os
RLno sl o
predi ceuna menor l ongevi dad
( en comparaci n con l a de l os RR) si no quese
acompaa
de una menor competenci a de sus si stemas i nmuni tari o
y
neuromuscul ar ydeal teraci ones neuroqu mi cas
enci ertas reas del SNC.
Los estudi os
de Ferrandi z et al . ( 1994) y de Mi quel et al . ( 1995)
confi rman quel os ratones hembra OF1 son
tambi n muyadecuados para
i nvesti gar l os efectos del
envej eci mi ento y de l a i ngesta de anti oxi dantes
ti l i cos
sobrel a bi oqu mi ca mi tocondri al .
De acuerdo con
todas l as i nvesti gaci ones resumi das aqu , hemos
escogi do el ratn
hembra OF1 como el model o msadecuado para profundi zar
enl os estudi os
sobreel efecto protector de l os anti oxi dantes ti l i cos
NACyTP
contra el estrs oxi dati vo i n vi vo as como
contra el deteri oro del a conducta y
de otros parmetros
fi si ol gi cos ybi oqu mi cos que ti ene l ugar al
envej ecer.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Volver al ndice/Tornar a l'ndex
OBJ ETI VOS
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
El obj et i vo
pri nci pal de est e t rabaj o es
est udi ar l os ef ect os ant i oxi dant es
de NACy
TPsobre cl ul as
de EPRb yrat ones OF1. Apart i r
de est e est udi o se
podr a pl ant ear
l a posi bi l i dad de ut i l i zarl os
como supl ement o
ant i oxi dant e
apropi ado
para f ut uros ensayos
cl ni cos en humanos
con s nt omas de
envej eci mi ent o premat uro
l i gados a al t os ni vel es
de est rs oxi dat i vo, para
l o
cual se proponen
dos model os de est udi o
sobre el envej eci mi ent o, uno
i n vi t ro
yot ro i n vi vo
con l os si gui ent es
obj et i vos concret os:
Model o de envej eci mi ent o
cel ul ar i n vi t ro
1.
I nvest i gar el dao cel ul ar
produci do por l a exposi ci n del
EPRb al
peroxi ni t ri t o ya ot ros agent es
qu mi cos i nduct ores
de est rs oxi dat i vo,
as
como por i ncubaci n
del EPRb en at msf eras
conbaj o cont eni do de ox geno
.
2. Est udi ar si l a
supl ement aci n del medi o con
vi t ami na C, NACo
TP
prot ege cont ra l os
menci onados procesos
pat ol gi cos .
Model o
de envej eci mi ent o i n vi vo
1.
Est udi o de di f erent es
parmet ros f i si ol gi cos y
bi oqu mi cos
i ndi cadores de envej eci mi ent o
.
2. I nvest i gar
si l a admi ni st raci n de
ant i oxi dant es, NACyTPa
part i r de
una
edad avanzada prot ege
cont ra el descenso
l i gado al a senescenci a.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Volver al ndice/Tornar a l'ndex
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
MATERIAL
YMTODOS
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
1. Materi al
Materi al yMtodos
1. 1. Materi al
bi ol gi co
Paral os estudi os i n vi tro
se uti l i zaron cul ti vos pri mari os en
monocapa
de l as cl ul as del epi tel i o
pi gmentari o de reti na bovi na
(EPRb), para el
ai sl ami ento
del as cl ul as del EPRbseuti l i zaron
de 6a8 oj os dereses
bovi nas
reci n sacri f i cadas en el
matadero muni ci pal de Al i cante,
que f ueron
transportados al l aboratori o
en un f rasco devi dri o estri l
sumergi doen hi el o.
Para
l os estudi os i n vi vo se uti l i zaron ratones
hembrasv rgenes del a
cepa
OF1 (al bi no ti po Swi ss) .
Fueron manteni dos en el
estabul ari o de l a
Uni versi dad de
Murci a en j aul as para ratones, 7
por j aul a, en condi ci ones
ambi ental es establ es deci cl ode
i l umi naci n de 12 horas de
l uz y12 horasde
oscuri dad y temperatura
ambi ente de 231C. Los
ani mal es f ueron
al i mentados con pi ensodemanteni mi ento
pararata/ ratn
deB&Kyagua ad
l i bi tum.
1. 2. Reacti vos
PBS, DMEM,
Suero Fetal Bovi no, Tri psi na
f ueron sumi ni strados
por Gi bco
BRL(Li f eTechnol ogi es,
Pai sl ey, Scotl and) .
Vi tami na
C, N- aceti l - ci ste na, ci doti azol i d n
carbox l i coo
ti oprol i na, Hoechst
33258, Ol i gomi ci na, ci do
i odoacti co, Ci anuro
sdi co f ueron sumi ni strados
por Si gma
(St. Loui s, MO. EE. UU. ) .
El Peroxi ni tri to f ue
sumi ni strado por Al exi s
Bi ochemi cal s (San Di ego,
EE. UU
. ) .
Todo
el materi al de cul ti vos
f ue sumi ni strado por
Costar (Cambri dge,
I ngl aterra) .
El Ki t de ATP
"ATP bi ol umi ni scence Assayki t HS
I I " f ue sumi ni strado por
Boehri nger Mannhei m.
El restode materi al y
reacti vos cuyoori gen nose ha
especi f i cado f ueron de
gradoanal ti cosumi ni stradospor Si gma,
Merck oPanreac.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Materi al yMtodos
1. 3. Aparataj e
- Bal anza( Sal ter el ectroscal e KE- 120) .
- Bal anzade
preci si nSal ter - ADNmodel oER- 60.
- Centr f ugade sobremesa( Eppendorf model o
5403) .
- Centr f ugaref ri gerada( BeckmanJ - 21 conunrotor J A- 20) .
- El ectrodode ox geno deCl ark( SYS203, I NSTECH) .
- Espectrof otmetroHi tachi ,
model oU- 2. 000.
- Homogeni zador
"Potter S" deTef l on- vi dri o( Braun, Bi otech I nternati onal ) .
- I ncubador BB16( Heraeus, Al emani a) .
- Lumi nmeteoLKBWal l ac, model o
1250Lumi nometer.
- Mi croscopi oi nverti do
def l uorescenci a ( Lei ca DMI RB/ E) .
- Si stema de el ectrof oresi s en gel de agarosa,
Mi ni cel l EC370M, EC 135 ( E- C
Apparatus Corporati on,
St
.
Petersburg, Fl ori da) .
- Vi deo- camaraSonymodel oDCR- PC- 1.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
2. Mtodosde
l osestudi osi n vi tro
2. 1 . Cul ti vode
cl ul asdel epi tel i opi gmentari o
dereti na
Materi al yMtodos
2. 1 . 1. Cul ti vospri mari os
El peri ododesde
l a obtenci ndel os oj os de
resesbovi nasdel matadero
muni ci pal
deAl i cante, hasta el comi enzo
del procedi mi entode
ai sl ami entoen
l auni daddecul ti vosf ue
i nf eri or adoshoras.
Las cl ul as
se ai sl aron si gui endoel
mtodopropuestopor Luoet al .
( 1996) con al gunas modi f i caci ones. En
pri mer l ugar se l avaron l os
oj os con
tampn f osf ato sal i no
de Dul beco ( PBS) estri l queconten a
50 Ng/mL de
gentami ci na y
2, 5
Fi g/mL
deamf oteri ci na
B. Sedi secci on el oj oy
l a porci n
anteri or f uedesechada,
permaneci endoi ntactal a copa
pti ca. Seel i mi naronel
cri stal i no,
el humor v treoy l a reti na. El
EPRbserecogi conpi nzas, cortando
por dondenosedesprend a
yse col oc enunapl aca
Petri estri l . Sei ncub
durante20mi n
a 37C con 5mLdeuna
sol uci n estri l detri psi na-EDTA
( tri psi na
1 : 250 0, 05%yEDTA
0, 54 mM) para as poder
di soci ar l as cl ul as.
Transcurri doel per odo
dei ncubaci n, se detuvo
l a acci n de l a tri psi na con
10 mL de medi o
de cul ti vo consi stente en una
mezcl a de medi oDul beco
modi f i cado
por Eagl e ( DMEM) ,
supl ementado con 3, 2 g/L
de bi carbonato
sdi co, 50
l gg/mL
degentami ci na, 2, 5
!i g/mL
deamf oteri ci na B, a pH7, 2,
compl etadocon suerof etal bovi no
( SFB) al 10%( desacti vado previ amente
a
56C durante
45 mi nutos) . A conti nuaci n se
trasvasaron l as muestras a
tubos
de centr f uga y se centri f ugaron
a 1000xg durante 10 mi n
en una
centr f uga de sobremesa
el i mi nando l os sobrenadantes.
Las cl ul as se
resuspendi eron en medi ode cul ti vo y l a
suspensi n cel ul ar resul tante se
sembrenbotel l asT-125del amarca
"Costar" ( Fi gura9) .
Los cul ti vos de
EPRbsemantuvi eron en uni ncubador
a37C, enuna
atmsf era
hmeda queconten a un 5%de
C02, hasta que al canzaron l a
conf l uenci a( 7-10d as) . Durante
esteti emposecambi el medi ocadadosd as
.
La conf l uenci a sedef i necomoel momentoaparti r
del cul l as cl ul as
cubrentodal a superf i ci edel a
botel l a, cambi andosumorf ol og a.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Materi al y
Mtodos
Extraer cri stal i no,
v treoyreti na
I ncubar con Tri psi na
L
Suspensi n
cel ul ar
15 mi n. a 37C
1000xg 10
mi n
Resuspender el preci pi tado en
medi ode cul ti vo
Sembrar en botel l as T-125
Subcul ti vos
Cambi odemedi ocada 2
d as
Fi gura 9. PROTOCOLODE
CULTI VOPRI MARI ODE CLULAS DE EPI TELI O
PI GMENTARI ODERETI NABOVI NA.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Materi al yMtodos
2. 1. 2. Subcul ti vos
Una vez
que l as cl ul as de EPRb
al canzaron l a conf l uenci a, se
real i zaron
subcul ti vos. Parael l o, pri mero
sel avarondosvecesl as
botel l as de
l os cul ti vos con10 mL
de
PBS
estri l yposteri ormente
se i ncubaroncon10
mLde
una sol uci n de tri psi na-EDTA a 37C
y5%de C02durante 15
mi nutos. Transcurri do este
ti empo se detuvo l a acci n
de l a tri psi na
aadi endo10
mLdemedi odecul ti vo.
Las cl ul as f ueron despegadas
de l as botel l as con
l a ayuda de un
raspador estri l de
pl sti co, serecogi eronen
untubocni codecentr f ugayse
centri f ugaron
a1. 000xgdurante 10
mi nutosenunacentr f ugade
sobremesa.
El
preci pi tadocel ul ar resul tante
seresuspendi en15mL
demedi odecul ti vo
yse vol vi a
centri f ugar enl as mi smas
condi ci ones conel f i n de el i mi nar
restos
de tri psi na. El preci pi tadocel ul ar
obteni do de cadabotel l a
enesta
segundacentri f ugaci n
se resuspendi en 10 mLde
medi ode cul ti voyse
sembr en
dosbotel l asT-125. Estas
cl ul asf ueronmanteni das
enl as mi smas
condi ci ones de
i ncubaci nycambi o de medi o
que l as cl ul as del cul ti vo
pri mari o
hastaqueal canzaronl aconf l uenci a
( entre7y10 d as) .
Decada cul ti vo pri mari o
se real i zarontres
subcul ti vos consecuti vos,
conl a suspensi n
cel ul ar f i nal obteni datras
el tercer subcul ti vo, se
real i zaron
l as
di l uci ones adecuadas para
sembrar en pl acas de 6
poci l l os ( 150. 000
200. 000
cl ul as/
poci l l o)
o
en
botel l as
de
75
cm2
( 1, 5-2
mi l l ones
de
cl ul as/ botel l a)
.
El recuentodel nmero
decl ul asyel porcentaj e
de vi abi l i dadde l as
mi smas se determi n
tomando una
al cuota de l a suspensi n f i nal
y
mezcl ndol aai gual vol umen
conunasol uci n al 0, 04%
deazul tri pn. El
rendi mi entomedi o
del as preparaci ones cel ul ares
obteni das aparti r de 6-8
oj os
deresesbovi nasf uede4a5mi l l ones
decl ul as.
Entodos l os experi mentos
seuti l i zaroncul ti vos
cel ul ares conf l uentes
trasuncuartopase
detri psi na.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Materi al yMtodos
2. 2. I nducci ndel daocel ul ar por estrs
oxi dati vo
Se uti l i zaron di f erentes condi ci ones experi mental es con el f i n de
exponer l as cl ul as de EPRbaunestrs oxi dati vo.
Seuti l i zaronl as cl ul as de
EPRbquehab ansi do sembradas
enpl acas de6poci l l os.
2. 2. 1. I nducci ndeestrs oxi dati vopor agentesqu mi cos
Enpri mer l ugar se reti r el medi odecul ti vodel os
poci l l os ysel avaron
l as cl ul as dos veces con tampn
sal i no (NaCI 145mM, KCl 5mM, CaC12
1,5MM, MgC12 1mM, NaH2PO41mM, gl ucosa5,6mM, Hepes
20mM, aj ustadoa
pH7,4) .
Se prei ncubaron l as cl ul as durante 10 mi nutos
en tampnsal i no a
temperatura ambi ente en l a campanade
f l uj o l ami nar. Aconti nuaci n, se
aadi eron al os
poci l l os correspondi entes l os di f erentes agentes qu mi cos
causantes del dao oxi dati vo y se i ncubaron a 37C, en una atmsf era
hmedaqueconten aun5%deC02, durante
15mi nutos.
Para l a i nducci n del
daocel ul ar se uti l i zaron l os si gui entes agentes
qu mi cos (Regoet al . , 1997) :
- Ascorbato 5mMe i nf erroso
100uMaunpHl i geramente ci do (pH
6,5) , provocandoas unaperoxi daci nl i p di ca.
- Ol i gomi ci na (OL) 5pg/mL,
i nhi bi dor del enzi ma ATP si ntetasa, y
Ci anuro sdi co
(CN) 2 mM, i nhi bi dor del enzi ma ci tocromo
c oxi dasa,
provocandoas unahi poxi aqu mi ca.
ci do i odoacti co (I AA) 0,5 mM,
i nhi bi dor de l a enzi ma gl ucol ti co
gl i ceral dehi do si ntetasa,
o bi en 2- desoxi gl ucosa (DG) 2mM,
que puede
susti tui r al agl ucosaenl a gl ucl i si s, cuandoseuti l i z
DG, seuti l i z tampn
sal i nosi ngl ucosa, dandol ugar
acondi ci ones de hi pogl ucemi a.
- La combi naci n de l os i nductores de hi poxi a
qu mi ca conunode l os
i nductores dehi pogl ucemi a, seuti l i z
parai nduci r unai squemi acel ul ar.
Transcurri do el ti empo dei ncubaci n
propuesto, 15mi n, l as pl acas se
col ocaron en un bao de hi el o (10
mi n) para i nterrumpi r l os di f erentes
est mul os.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Materi al yMtodos
A
conti nuaci n se col ectaron l os medi os
dei ncubaci n, con el f i n de
medi r posteri ormente l a acti vi dad
LDHsecretadapor l as cl ul as al
medi ode
cul ti vosi gui endoel protocol o
descri toen el apartado
2. 4 . 1 . Trasl arecol ecci n
de l os
medi os, l as cl ul as de cada
poci l l o f ueron despegadas medi ante
tri psi ni zaci n,
aadi endoa
cadapoci l l o 0, 5 mLdeunasol uci n
detri psi na-
EDTAei ncubandodurante
15 mi nutosa37Cy5%
deC02. Transcurri doeste
ti empo, se
detuvo l a acci n de l a tri psi na
aadi endo 1 mLde medi o
de
cul ti vo,
l as cl ul asf ueron
recogi dasdel ospoci l l osconl a
ayudadeunapi peta
Pasteur de
pl sti coestri l , yse
traspasaronauntubocni code
centr f uga. Se
centri f ugaron a1 . 000xg
durante10 mi nutosen
unacentr f ugadesobremesa,
descartndose
el sobrenadanteyresuspendi endo
el preci pi tadocel ul ar
en 0, 5
mLde PBS
. Se cont el
numero de cl ul as en una
cmara Neubauer,
manteni ndol as a4C
para real i zar posteri ormente
l as determi naci ones
de
acti vi dad
LDHcel ul ar yATP
descri tas en l os apartados
2. 4. 1 y2. 4. 2,
respecti vamente
.
Enl as pl acas desti nadas
al a detecci n de
apoptosi smedi anteti nci n
con
bi s- benzami da, l aacci nde
l osest mul osf uedeteni da
medi antel aadi ci n
de1 mLdeparaf ormal deh do
al 4%entampn
f osf ato0, 1 M( PFA) , f i j ando
as
l as cl ul asy
manteni ndol as
aCChastael momentodereal i zar l a
ti nci n.
2. 2. 2. I ncubaci n
de cl ul as de EPRb
en unaatmsf eracon
baj a
concentraci n
deox geno
Seuti l i zaron cl ul as
sembradas en l as pl acas de
6 poci l l os yen l as
botel l asde75cm2.
En
pri mer l ugar sereti r el medi o
decul ti voysel avaronl as
cl ul ascon
PBS, aconti nuaci n, se
l es aadi 2 mLdemedi o
decul ti vo si n SFByse
i ncubaron
durante72 horasa37C, en
unaatmsf erahmeda
queconten a
un 5%de C02 yun 3%
de ox geno en un
i ncubador BB16: Heraeus.
Paral el amente,
comocontrol se
i ncubaron cl ul as conysi n SFB
al 21%de
ox geno.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Materi al yMtodos
Unavez transcurri doel peri ododei ncubaci nl as cl ul as del as pl acas
de6poci l l os se col ocaronenunbaode
hi el o durante
10
mi n, y
se
procedi
como
enel apartado2. 2. 1.
Las cl ul as que se hab ansembrado enl as botel l as de 75 cm2 se
uti l i zaron paral a cuanti f i caci ndeADNsi gui endo
el
protocol odescri to
en
el
apartado
2. 4
. 3
. b.
2. 2. 3. I nducci n
de
estrs
oxi dati vopor peroxi ni tri to
Enpri mer l ugar se reti r el medi odecul ti vo, l as cl ul assel avarondos
vecesconPBSyposteri ormente
se
aadi
el agenteoxi dante(PN50 o100J uM)
preparadoenmedi ode
cul ti vosi nSFB. Lascl ul as se i ncubaronduranteuna
hora a37C
en
una
atmsf era hmedaque conten a un5%de C02
.
Las
condi ci onescontrol consi st anenl a i ncubaci ndecl ul as enmedi o
de
cul ti vo
si n SFB. Laacci ndel PNse i nterrumpi
col ocandol as pl acas enbaode
hi el o(10 mi n) yaconti nuaci nsesi gui el protocol odescri toanteri ormente
.
2. 3. Tratami entoconanti oxi dantes
Seestudi , por separado, el ef ectodel aVC
300
pLM,
NAC10
mM
yTP3
mM. Parai nvesti gar este ef ecto, l as cl ul as
se i ncubaronconel anti oxi dante
sel ecci onadodurante2horas
antes de i nduci r el daocel ul ar. Transcurri do
esteperi odode
i ncubaci n, se aadi eronl os est mul os correspondi entes para
produci r el estrs oxi dati vo acadapoci l l omanteni endol a
concentraci nde
anti oxi danteduranteel
peri ododei ncubaci nconel est mul o.
2. 4. Determi naci n
del daocel ul ar
2. 4. 1. Determi naci n
del aacti vi dadl actatodeshi drogenasa
Se determi n tantol a acti vi dad LDHsecretadaal
medi oextracel ul ar
(medi odei ncubaci ndel ascl ul as) como
l aacti vi dadLDHentracel ul ar.
La medi das de
l a acti vi dad enzi mti ca de LDHse real i zaron
espectrof otomtri camente
si gui endol a conversi ndeNADH
a
NAD+a
340nm
segnel mtododescri topor Bergmeyer yBrent
(1974) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Materi al yMtodos
Laacti vi dad de l a LDH
secretadaal medi oextracel ul ar se val or
en
l as
muestrasdemedi oquehab ansi do recogi das
di rectamentedel os poci l l os.
La absorbanci a
se determi n tras aadi r auna cubeta: 100
uL de
muestra, 0, 5
mL de pi ruvato 9, 72 mM,
i ni ci ando l a reacci n con2, 5 mL de
NADH
0, 24 mM. Todas l as medi das
se real i zaron f rente aunbl anco que
conten atampnLDH( Tri s
81, 3 mM, NaCI 0, 2 MpH7, 2)envez
depi ruvato.
La
di smi nuci n de absorbanci a ( debi do al a
oxi daci n del NADH) f ue
segui da a
i nterval os de 30 s durante 3
mi n, a30 Cenunespectrof otmetro
model oU-2. 000 Hi tachi .
Para
determi nar l os ni vel es
de LDHi ntracel ul ar, l as cl ul as
f ueron
l i sadas conTri tnX-100
0, 1%preparadoenTampnLDH, y
centri f ugadas a
7. 000xg durante
15 mi na 4C, uti l i zando
el sobrenadante para
medi r l a
acti vi dad de LDH, si gui endo
el mi smoprocedi mi ento
descri to anteri ormente
paradetermi nar
l a acti vi dadLDHextracel ul ar
.
La concentraci nde l a
acti vi dad catal ti ca ( [ AC] ) del enzi ma
LDHse
cal cul medi antel a si gui ente
ecuaci n:
[ AC] =DA/At
x
V/ 1
. 000 x Ex dx v
( mol x s
-1x
1-1)
donde
DA/At se ref i ere al
cambi o de absorbanci a, E es
el coef i ci ente de
absorci nl i neal del
NADH( 0, 63 Lx mmol -1
x mm-1), des el pasode l uz de
cubeta
( 10mm), Ves vol umenf i nal
del ensayo( 3, 1 mL) yv
es vol umende
muestraaadi do
( 0, 1 mL) .
Los resul tados se expresaron
como %de LDH
secretado al medi o
respecto a l a acti vi dad
LDHtotal , si endo
el LDHtotal l a suma de LDH
i ntracel ul ar y
deLDHpresente enel medi o
decul ti vo.
2. 4. 2. Determi naci ndel a
concentraci ni ntracel ul ar deATP
El ATPi ntracel ul ar
se determi n por bi ol umi ni scenci a
uti l i zandounki t
comerci al : "ATP
bi ol umi ni scenceAssayki t
HSI I " deBoehri nger Mannhei m.
El protocol o
experi mental f ue el si gui ente:
una vez f i nal i zado el
experi mento, l as cl ul as se recogi eron
medi ante tri psi ni zaci n, yse
cont el
nmerodecl ul as.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Materi al yMtodos
Se tomuna
al cuota correspondi ente a25. 000 cl ul as que se l i saron,
contampndel i si s sumi ni stradoconel ki t,
durante5mi nutos atemperatura
ambi ente.
Parareal i zar l as determi naci onesdeATPsepi petearon
100 l i L del i sado
cel ul ar y100j i L de l uci f erasa,
e i nmedi atamente se real i z l a l ectura enun
l umi nmetro
(LKBWal l ac model o1250 Lumi nometer) durante
10 s.
La concentraci n de
ATP i ntracel ul ar se cal cul extrapol ando l as
l ecturas obteni das
aunarectade concentraci ndesde 1mMhasta 1 nMde
ATP. Los
resul tados seexpresaroncomo[ ATP] (^/25
. 000 cl ul as.
2. 4. 3 . Detecci ndel a apoptosi s
Exi stendi f erentes
parmetros quei ndi canquesehaproduci doundao
cel ul ar e
i ncl uso i ndi cadores de muerte cel ul ar
comosonl a ti nci nconazul
tri pn, connaranj a de
acri di na- bromuro de eti di o o con
Hoechst 33258. La
vi sual i zaci nconunsol omtodonoes suf i ci ente
por l o quehayquereal i zar
otrosmtodos compl ementari os
comoel ectrof oresi s engel deagarosa, TUNEL,
etc. (Sastryet a1
. , 2000) . Eneste estudi osehauti l i zado
l a Ti nci nconHoechst
33258
compl ementadapor l ael ectrof oresi s en
gel deagarosa.
a) Ti nci nconHoechst
33258(bi s- benzami da)
Esta tcni ca permi te
vi sual i zar deunamanera senci l l a
yrpi da si l as
cl ul as han
suf ri do apoptosi s. La tcni ca
se basa enl a uti l i zaci n de bi s-
benzami da (Hoechst 33258),
compuestoque se uneespec f i camente
al surco
menor del
ADN, noes i ntercal ante ycompi te
conl a prote na hi stni ca
Hl .
Esta mol cul a cuando
se exci ta conl uz ul tra vi ol eta
(360 nm) emi te una
f l uorescenci a
quese reci be conunal ongi tud
deondaquese correspondecon
el col or azul (emi si nde490- 500nm) .
Conestatcni caseproduceunmarcaj e
espec f i codel ncl eocel ul ar en
el casode cl ul asapoptti cas
vi sual i zndosel a
`f ragmentaci n" del
ncl eo.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Materi al y
Mtodos
Unavez f i nal i zado el peri odo
de i ncubaci n, se real i z l a ti nci ncon
Hoechst 33258
(bi s- benzami da) para determi nar si se
hab a i nduci do
apoptosi s, sesi gui el mtododescri to por
Boi xet al . (1997) .
Enpri mer l ugar
l as cl ul as se f i j aronaadi endoa
cada poci l l o de l a
pl acay
sobreel mi smomedi odei ncubaci n,
1 mLde paraf ormal dehi do al 4%
en tampnf osf ato 0, l M
(PFA), i ncubando l uego durante
10 mi nutos. A
conti nuaci nse
reti r el medi oyse vol vi aaadi r
1 mLde PFA, i ncubando
durante
15mi n. Posteri ormente,
se
l avaron
l as cl ul as tres veces con
PBS, se
aadi 1 mLde Hoechst
33258 (0, 05 l i g/ mL) y
se i ncub enoscuri dad
durante 25 mi n
atemperatura ambi ente.
Unavez termi nado el peri odo de
i ncubaci n,
sereti r el Hoechst, y
l as cl ul as se l avarontres vecescon
PBS. A
conti nuaci n, acada
poci l l ose l e aadi eron
25
pL
de medi oPBS- gl i cerol en
l a
proporci n
2: 1 y se l e col oc
uncubre para as poder
proceder a su
observaci ncon
l uz ul tra- vi ol eta enun
mi croscopi oi nverti dode f l uorescenci a
(Lei ca
DMI RB/ E) .
b) El ectrof oresi s en
gel deagarosa
Paral a
real i zaci ndel ael ectrof oresi s
engel deagarosaseuti l i zaronl as
cl ul as
de EPRb que f ueron
sembradas en botel l as de
75 cm2 (1, 5- 2x106
cl ul as/ botel l a) . Antes
de l a el ectrof oresi s se
real i z l a extracci nde ADN
si gui endo el
protocol o descri to por
Ramachandra et al . (1995) con
al gunas
modi f i caci ones.
Tras tri psi ni zar l as cl ul as,
se contaronenunacmara
Neubauer yse
centri f ugarona 7. 000xg
durante 10 mi n a
temperatura ambi ente enuna
centr f uga de
sobremesa. El sobrenadante
se descart yel preci pi tado
se
resuspendi
con250pLdetampn
de di gesti n(CI Na 100
mM, Tri s 10mM,
EDTA 25 mM,
SDS 0, 2%, protei nasa K0, 2 mg/ mL,
aj ustado a pH8) . Se
i ncubaronl as
cl ul as del preci pi tado en
unbaoa50Cdurante 12 horas.
Transcurri doeste peri odode
i ncubaci n, seaadi acadamuestra
250pLde
una sol uci nde
f enol / cl orof ormo/ i soami l al cohol (25
: 24: 1) yse centri f ug a
7. 000xg
durante5mi na4C.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Materi al yMtodos
La
f ase acuosa( f ase superi or) sepi pete ysecol oc en otro vi al .
Esta
operaci n se repi ti dos
veces. Al l ti mosobrenadanterecogi dose l e aadi
unvol umen
deacetatoamni co7, 5 Mi gual al ami taddel vol umen
recogi do.
Seagi t el tubo
duranteunos segundosyse aadi eron dos vol menesde
etanol
absol uto, i ncubando durante dos horas a 4C. Posteri ormente,
se
centri f ug a 7. 000xg durante 30 mi n a temperatura
ambi ente. El
sobrenadante se decant
yel preci pi tado se resuspendi con el mi smo
vol umenf i nal
( Vol f i nal = vol f aseacuosa+ 1/ 2 vol acetatoamni co7, 5
M+2 vol
etanol ) deetanol al 70%, centri f ugndosedenuevoenl asmi smas
condi ci ones.
Trasl al ti macentri f ugaci n,
seaspi r el sobrenadantecui dandode
no
tocar el
preci pi tadoysedej secar enl a campanadef l uj ol ami nar
durante1
hora. El preci pi tadose
di l uy entampnTri s 1 0mM, EDTA5 mM, aj ustado
a
pH8( Tampn
TE) ysetom unaal cuotaparadetermi nar l a
concentraci nde
ADN
espectrof otomtri camentea
260/ 280nm( Fi gura1 0) .
Unavez
determi nada l a concentraci n de ADN, se pas a
real i zar l a
el ectrof oresi s
en gel deagarosa, paral ocual se real i zaron
di f erentes pruebas
deconcentraci n deagarosa,
0, 8%, 1 %, 1 , 2%, 2%. Laconcentraci n i dneade
agarosaf ue del 1 %y
se prepar en tampn TBE0, 5x ( Tri s 0, 089
M, ci do
bri co 0, 089
M, EDTA 0, 002 MpH8) . Sereal i z unadi l uci n
del ADNpara
obtener
unaconcentraci nf i nal
de
ADN
de1 0 pg, quesei ncub con
0, 1 Ude
ARNasa
durante 1 horaa37C, conel f i n dedi geri r
posi bl es restos deARN.
Transcurri doel ti empodei ncubaci n
seaadi eron2 pL debromof enol - xi l en-
ci anol acada
muestrayse deposi taron en l os poci l l os del gel
de agarosa
real i zando
l a el ectrof oresi s en TBE0, 5x a64V y
45mA. Unavez queel ADN
hubotermi nadosu recorri do
enel gel ( aproxi madamente 4horas) ,
se real i z
unaf otograf i adel
gel conunavi deo- camaraSonymodel oDCR- PC- 1
.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
I nducci ndeapoptosi s
t
Recoger l as cl ul as
1
Aadi r 250pul de Fenol / cl orof ormo/
i soami l al cohol
I ncubar con
250
j i 1
tampnde di gesti n
1 2h. a50C
c~
7
. 000xg, 5mi n, 4C
Recoger f ase
acuosaymedi r vol umen
Repeti r dos
l ti mos pasos 2veces
Aadi r
1 / 2 del vol umenrecogi do de acetato
amni co7, 5M
agi tar conl os dedos suavemente
unos segundos
Aadi r 2vol menes deetanol absol uto
2h. a4C
7. 000xg, 30
mi n, Ta ambi ente.
Decantar sobrenadante
yaadi r mi smovol umende
etanol al 70%
Reti rar
sobrenadarteysecar preci pi tado
encampanadef l uj o l ami nar
Medi r
[ ADN]
t
t
Di l ui r en
tampnTE
Fi gura 1 0. PROTOCOLODE
EXTRACCI NDEADN
Materi al yMtodos
t 7. 000xg, 30mi n, T' ambi ente.
El ectrof oresi s engel deagarosa
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
3. Mtodosdel
model odeenvej eci mi entoi nvi vo
Materi al yMtodos
3. 1 . Ani mal esde
experi mentaci n
Comoyahemosi ndi cado, l os ratones
hembra OFl f ueronestabul ados
en el ani mal ari o enj aul as para roedores
en un nmerode 7 por j aul a.
Duranteel estudi o
f ueronmanteni dosencondi ci ones
ambi ental esestabl es de
ci cl o de
i l umi naci n de 12 horas de l uz y
12 horas de oscuri dad,
y
temperaturaambi ente
de 231 C. Losani mal esf ueron
al i mentadosadl i bi tum
con
pi ensode manteni mi entopararata- ratn
de B. K .
El estudi ose
i ni ci conunapobl aci nde 96ani mal es
de 32 semanas
de edad,
exentosdepatol og as macroscpi cas,
yal aedadde 40
semanas, f ue
di vi di da al azar entres
grupos: ungrupocontrol ydosgrupos
cuyadi eta f ue
supl ementadacon
anti oxi dantes. Losgrupos quereci bi eron
l os anti oxi dantes
en l a di eta
f ueron al i mentados ad l i bi tum
con el mi smo
pi enso de
manteni mi entopara rata- ratn
B&Ksupl ementadoconuno
de l os si gui entes
f rmacos anti oxi dantes
: TPal 0, 2%oNACal
0, 2%(peso/ peso) en
el pi enso
seco, mi entras que el grupo
control se si gui al i mentando
conel pi ensode
manteni mi ento
para rata- ratn de B&K. El
pi enso supl ementado
con l os
f rmacos
anti oxi dantes (preparado
por el Centrode
I nstrumentaci nCi ent f i ca
de l a Uni versi dad
de Granada) f ue consumi do
dentrodel pl azomarcado
para
evi tar
su al teraci n. Entodosl os
casosel sumi ni strode agua
si empref ue ad
l i bi tum.
Desde el
momentoenel que se i ni ci el
tratami entoconl os
f rmacos
anti oxi dantes (a l a edadde
40 semanas) hasta que
termi n el estudi o se
real i zaron dos
sacri f i ci os de ani mal es (para obtener
muestras de h gadoy
ri onesparal os estudi os bi oqu mi cos)
al as edadesde48y
62 semanas.
Esteestudi o
f ue real i zadodeacuerdoconl as
di rectri ces del Consej o
de
Europa y
el R. D. 223/ 88 sobre l a
uti l i zaci n ycui dado
de ani mal es de
l aboratori oyf ue
aprobadopor l a Uni versi dadde Al i cante
.
Ladi f erenci ade edadentre l os ratonesdel a
pobl aci ni ni ci al era
de1
semana.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Materi al yMtodos
3. 2. Estudi ode
parmetros noi nvasi vos f i si ol gi cos yconductual es
Para l os estudi os l ongi tudi nal es, f i si ol gi cos yde conducta, se
i denti f i c
cada ani mal perteneci ente a cada j aul a de 7ani mal es,
marcandol os ratones
enel pel omedi ante
ci do
p cri co
di l ui do. Lacabeza, l a porci nsuperi or de l as
patas
del anteras o traseras, el extremo caudal del l omo ysu
combi naci n
f ueron l os l ugares marcados. Este procedi mi ento
de i denti f i caci n, por su
i nocui dadpara
el ani mal , susenci l l ez ypor supermanenci a sobre el
pel o del
ani mal , es adecuado para estudi os de
medi a yl arga duraci n enroedores
al bi nos.
3. 2. 1 . Consumopromedi ode al i mento
Parti endode 600g de
pi enso seco i ni ci al por j aul a, l a i ngesta medi ase
determi n
para cada grupo de estudi o medi ante l a determi naci n
qui ncenal ,
por j aul a, del peso del al i mento que quedaba enel
comparti mento. Los datos
se expresancomogramos
de pi ensoconsumi dopor ratnypor d a.
3. 2. 2. Determi naci ndel pesocorporal
El peso corporal
es
uno
de l os parmetros f i si ol gi cos ms f ci l es
de
determi nar,
y adems proporci ona gran i nf ormaci n
acerca del estado de
sal uddel ani mal . Eneste estudi o
el peso corporal se mi di qui ncenal mente
para cada ratn,
perteneci ente a l os di sti ntos grupos: control , NACy TP,
durante todo el
peri odo de estudi o. Para el l o se uti l i z una bal anza
( Sal ter
el ectroscal e KE-120) conunapreci si nde 0, 1
g.
3. 2. 3.
Parmetros de conducta
Todas l as eval uaci ones de l as pruebas
de vi tal i dad y conducta se
l l evaron acabo en
el mi smoreci nto donde estabanestabul ados l os
ratones,
conel f i n de evi tar posi bl es i nterf erenci as
en l as respuestas de l os ani mal es
debi do al transporte a otro l ugar
de ensayo. Los ensayos se real i zaron
entre
l as 10y14horas
parami ni mi zar posi bl es vari aci ones enri tmos
ci rcadi anos.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Materi al
yMtodos
a) Medi da deparmetros devi gor ycoordi naci n neuromuscul ar
La eval uaci n de l a vi tal i dad de l os ani mal es se real i z uti l i zando l a
tcni ca de l a cuerda ti rante descri ta por Mi quel yBl asco ( 1978) . La correcta
resol uci n del test requi ere que l os ratones superenenunmi nuto o menos l a
prueba de l l egar
a
uno
de l os dos extremos de unacuerdatensadayf i na, tras
suf ri r unasi tuaci nde estrs comoes, al comi enzo de cada prueba, suj etarl os
del rabo conl os dedos, col gando cabeza abaj oydej ndol es coger, ai nterval os,
l a cuerdaconl as patas anteri ores
.
Cuando se col ocan ani mal es j venes en l a cuerda, son
capaces de
agarrarseael l a conl as cuatro
patas ei ncl uso el raboydespl azarse enmenos
de un mi nuto hasta al canzar uno de l os dos extremos.
Los ratones vi ej os
suel en permanecer i nmvi l es
col gados con l as patas del anteras o bi en, al ser
i ncapaces de suj etarseconl as cuatropatas, caenal l echo de
vi rutas.
Se empl e una cuerda de
camo de 1 mmde di metro atada a dos
soportes f i rmemente
ancl ados y di stantes entre s 60 cm, quedando a una
al tura de 40 cmsobre un l echo de vi rutas. La cuerda
se tens al mxi mo
( Fi gura 11) .
Fi gura 11. Procedi mi entode eval uaci ndevi gor ycoordi naci n neuromuscul ar
medi ante l a tcni ca de l a cuerda ti rante. Ratncapaz de al canzar unextremode
l a cuerdaenmenosde 1 mi nuto
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Materi al yMtodos
El procedi mi ento f ue el si gui ente: se reti r al ratn de su j aul a de
procedenci acogi ndol o cui dadosamente de l a col a, yse mantuvosuspendi do
de l a col a durante 10 s aunadi stanci aaproxi mada de 1 cmde l a cuerda en
su parte central . Se l e permi ti durante estos 10 s l l egar atocar yagarrarse
con suspatas del anteras al a cuerdaen tres ocasi ones. Fi nal mente, se dej al
ani mal agarrado de el l a sol amente con l as
patas anteri ores yse comenz a
cronometrar el ti empo quetardabael ratn en al canzar unode l os extremos
del a cuerda.
Si
el ani mal
ca aal l echodevi rutas ono
l l egaba
aunodel os dos
extremos de l a cuerda, i nmedi atamente se repet a por segunda vez l a
mani pul aci n anteri or pero manteni ndol oen esta ocasi n suspendi do de l a
col a tan sol o 5s.
La correcta resol uci n del test requi ere que l os ratones l l eguen en un
mi nutocomomxi moa cual qui era de l os dos extremos de l a cuerdatensada,
bi enen el pri mero, bi en en
el segundoi ntento. Una
vez
f i nal i zada estaprueba,
se reti ra al ani mal ya conti nuaci n se l e somete al si gui ente test, en el
l aberi nto
en
f orma
deT.
b) Medi dadel aacti vi dadexpl oratori aespontnea.
Se uti l i z unl aberi ntoen Ten base adi seos empl eados
paraestudi os
de memori aespaci al por DeanyKey( 1981) . El
l aberi ntoconsta de tres brazos
en f orma
deT, con paredes de congl omeradode maderacuyas caras i nternas
estnrecubi ertas con chapapl sti ca negra. Lamedi dade cada brazodel a T
es
25 cmde l argo, 10 cmde ancho y20 cmde al to ( todas l as medi das
ref eri das soni nternas) . El suel odel l aberi ntoconsi ste en vari l l as ci l ndri cas
de
al umi ni ode
3mmdegrosor, col ocadasaunadi stanci ade 5mmunadeotra y
si tuadas transversal mente al as paredes l ateral es decadabrazo( Fi gura 12) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Materi al yMtodos
Fi gura 12. Laberi nto en Tpara l a eval uaci n de l a acti vi dad expl oratori a
espontnea. Ratnsal i endodel pri mer brazo.
El l aberi nto
se col oc en el suel odel reci ntodel estabul ari osobrepapel
bl anco de f i l tro. En todos l os ensayos f ue si tuado en l a mi smaposi ci n baj o
i l umi naci n di recta de dos tubos f l uorescentes de 25w, en el techo,
a
una
di stanci a de 2, 5 m. La i l umi naci n sobre l os tres brazos del l aberi nto eral a
mi sma.
El mtodo para medi r l a acti vi dadexpl oratori a espontnea se bas en
el ti empoempl eadopor unratn enrecorrer el brazodesal i da del l aberi nto en
f orma
de Tsi n sumi ni strode cual qui er ti po deref uerzo posi ti vo o aversi vo. Se
tom al ratn por l a col a cui dadosamente y se col oc j unto a l a pared del
brazo de sal i da de l a T, en l a zona di stal de l a convergenci a de
l os brazos. Se
comenz aregi strar el ti empo ( en segundos) conun cronmetro hastaque el
ani mal sal i compl etamente ( con l as cuatro patas) del brazo i ni ci al . Se l e
concedi unasegunda oportuni dadal ratnque no consi gui sal i r del brazo
del l aberi nto enmenosdeunmi nuto( DeJ uan, 1994) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Materi al yMtodos
3. 3. Obtenci ndemuestrasdetej i dospara
estudi osbi oqu mi cos
El sacri f i ci o de l os ani mal es se real i z por decapi taci n previ a l i gera
anestesi aenatmsf eradeter .
I nmedi atamente se pasal a extracci n del os
rganos ( h gado y ri ones) de cada ani mal .
Todo el procedi mi ento de
ai sl ami ento se real i z a 4C.
Cada rgano de cada ani mal f ue procesado
i ndi vi dual mente parael ai sl ami entodemi tocondri as.
3. 4. Ai sl ami entode mi tocondri as de h gadoyde ri nydetermi naci n
deprote nami tocondraal
El
ai sl ami ento de mi tocondri as de h gado y ri n se l l ev a cabo
si gui endo el protocol o descri to por Ri ckwood et al . ( 1987)
con al gunas
modi f i caci ones
.
Tanto el h gado como l os ri ones
se l avaron 3 veces con tampn
compuestopor sacarosa
250mM, al bmi na sri ca bovi na ( BSA) 0,5 mg/mL,
Tri s/Cl H
10 mM, EDTA-K1 mMa pH7,2 aj ustado con KOH
( tampn de
ai sl ami ento) con l a f i nal i dad de el i mi nar restos
de sangre. Se col ocaron l as
muestras de tej i do en
unpapel def i l tro paraabsorber el excesodetampn, y
se
pesaronenunabal anzaSal ter-ADNmodel oER-60conuna
preci si nde 0,1
mg
.
Se trocearon y homogeni zaron en tampn
de ai sl ami ento a l a
concentraci n de
30%para h gado y 10%para ri n ( p/v) . Se real i z
una
homogeni zaci n en unhomogeni zador ti po "potter"
de Tef l on-vi dri o ( Braun)
medi ante 10
pases de mbol o ( 0,1 mmdehol gura) a1000 rpmeni nterval os
de
10s.
Unavez obteni doel homogeni zado,
f uel l evadoaunvol umenf i nal de15
ml , se centri f ug
a 1000xg durante 10 mi nutos a4Cen unacentr f uga
de
sobremesa, se desechel preci pi tado f ormadodencl eos
cel ul ares ymateri al
cel ul ar pesado. El sobrenadante
recogi do se centri f ug a8700xg durante 10
mi na4Cen
unacentr f ugaref ri gerada( BeckmanJ -21 conunrotor J A-20)
.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Materi al y
Mtodos
Unavez f i nal i zada
l a centri f ugaci n, el sobrenadante
se desech, y
el
preci pi tado,
es deci r l a f racci n
mi tocondri al , se resuspendi
en20 mLde
tampn
de ai sl ami entoyse
vol vi acentri f ugar enl as condi ci ones
anteri ores
para l avar yel i mi nar
restos cel ul ares. Se desech
el sobrenadante y el
preci pi tado f ue
resuspendi do enel m ni mo
vol umenposi bl e ( 1- 1, 5
mL) de
tampn
deensayo, tampn
consti tui dopor mani tol 220
mM, sacarosa70mM,
al bmi na
sri ca bovi nal i bredeci dos
grasos ( BSAFFA) 0, 5 mg/mL,
Tri s/CI H
50 mM, Cl K30mMapH7, 4
aj ustadoconKOH, manteni doa
4 Chasta su
uti l i zaci n ( Fi gura
13) . Se tom una al cuota
para l a determi naci n
del
consumo
deox genopor l as mi tocondri as
de h gadoyri n.
El homogeni zado
mi tocondri al restante f ue al i cuotado
ycongel ado a
- 80Chastael
d a enque se real i zaronl as
determi naci ones enzi mti cas
que
sedescri bena
conti nuaci n.
Ladetermi naci n
deprote nas se real i z uti l i zando
el mtododeLowry
( Lowryet al . ,
1951) . El rendi mi ento
obteni do durante este proceso
f ue del
ordende 10, 633, 5 y
3, 78 1, 18 mg de prote na por
gramode h gado y
ri nrespecti vamente
.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Materi al yMtodos
Anestesi a
conter
Decapi taci n
Extraer rganos (h gadoyri ones)
17
Lavar rganos
Pesar rganos
Homogeni zar rganosen Potter tef l n- vi dri o"Braun"
1 . 000 rpm, 10 pases cada l Os
Centri f ugar
homogenado 1 . 000xg 10 mi n4C
Preci pi tado
Sobrenadante
Sobrenadante
Resuspender
en20mLdeTampnAi sl ami ento
C~s 8700x
g
10
mi n
4C, rotor J A20
Preci pi tado
Ct2 8. 700xg10mi n4C, rotor J A
20
Preci pi tado
W
Resuspender en. 0, 5 mLdeTampnEnsayo
Determi naci ones
Sobrenadante
FIGURA13. Protocol odeai sl ami entodemi tocondri as
de h gadoyri n.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Materi al yMtodos
3. 5. Determi naci ndel
consumodeox geno
La
acti vi dad respi ratori a de l as
mi tocondri as f ue determi nada con
un
el ectrodode ox genode Cl ark
(SYS203, I NSTECH), si gui endoel
mtododesci to
por Pal mero(1992)
.
La
cal i braci n del el ectrodo se
real i z mi di endo el porcentaj e de
saturaci nde ox genoen 600pLde
tampnde respi raci n (tampnde
ensayo
supl ementadocon
rotenona 4pi M, acetato l OmM,
f osf ato 0, 2mM, C12Mg3mM,
EDTA-K
0, 5mM) en l a cmara
bi en sel l ada. Lal ectura del
0%de ox geno se
obtuvomedi ante
l a adi ci nal medi ode
di ti oni to sdi co.
Tanto l a cal i braci n del el ectrodo
de ox geno como el
estudi o de l a
respi raci n mi tocondri al
se l l ev a cabo a 25C,
manteni endo si empre l a
sol uci n
enagi taci nsuaveyconstante
.
Ladetermi naci ndel
consumodeox genose i ni ci
unavez estabi l i zada
l a l ectura del el ectrodo
de ox geno con tampn
de respi raci n. En ese
momentose aadi
l a suspensi nde
mi tocondri as (40 pL de
mi tocondri as de
h gadoyri n), ysedej que
se estabi l i zara el regi stro
(al rededor de30s) .
La respi raci n
mi tocondri al
se i ni ci con l a adi ci n
al medi o de
succi nato 5 mM, sustrato
de l a cadena de transporte
el ectrni co, (estado4
o
de reposo de l a
respi raci n), regi strando
l ecturas cada 30
s. Una vez
estabi l i zado el consumode
ox geno, se aadi al medi o
ADP0, 4mM, pasando
al estado3o
estadoacti vo de l a respi raci n
mi tocondri al , regi strando
l ecturas
cada30
s.
El ndi ce de respi raci n
mi tocondri al seobtuvoa
parti r del a rel aci n de
l os estados 3 y4 de
l a respi raci n, consti tuyendo
sta una medi da del
acopl ami ento
entre f osf ori l aci n
oxi dati va yrespi raci n
mi tocondri al . Los
resul tados se
expresancomopmol es de
02consumi do/mgprote nax mi n.
Para comprobar l a i ntegri dad
de l a membrana mi tocondri al ,
tras l a
adi ci ndeADPse
aadi al medi ouni nhi bi dor
del transporte denucl eti dos,
atracti l si do
60 j i M, sedej estabi l i zar al
si stemayse vol vi aaadi r
ADP.
Tanto mi tocondri as
como sustratos se
aadi eron uti l i zando
mi croj eri ngas,
evi tandol a f ormaci nde burbuj as en
l a cmara.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Materi al y
Mtodos
3. 6. Determi naci onesenzi mti cas
Se real i z l a
determi naci n de l a acti vi dad de dos enzi mas
mi tocondri al es, ci trato
si ntetasayci tocromocoxi dasa.
3. 6. 1 . Acti vi dadci tocromocoxi dasa
Seescogi este enzi ma
para el presente estudi oporque es el compl ej o
respi ratori o mi tocondri al msal teradopor el envej eci mi ento
( Ferrandi z et
al . ,
1994) .
La acti vi dad ci tocromo c oxi dasa ( EC 1
. 9. 3
. 1),
se determi n
espectrof otomtri camente, segnel mtodo
descri to por WhartonyTzagal of f
( 1967) . Semi di l atasade oxi daci n
del ci tocromocreduci doaoxi dadopor el
enzi ma
ci tocromocoxi dasa, uti l i zandocomobl anco f erri ci ani da
potsi ca.
Enpri mer l ugar, se reduj o el ci tocromo
c al 1%: Parael l o, se pes el
ci tocromo
c yse di sol vi en el 60%del vol umenf i nal contampnf osf ato
potsi co ( KP) 10 mM, al que se l e aadi
una punti ta de esptul a de
borohi druro de sodi o ( NaBH4)
para reduci rl oyse i ncub durante
30
mi na
4C
. Transcurri do el peri odo de i ncubaci n,
ypara el i mi nar el exceso de
NaBH4, se aj ust el pHde
l a di sol uci n a7, 1 conci do cl orh dri co 1Nyse
l l ev al vol umen
f i nal .
La
absorbanci a se determi n aadi endo a
una cubeta 0, 1 mL de
tampn KP 10 mM, 0, 07 mL
de ci tocromo c reduci do y0, 81 mL de agua
desti l ada.
Para i ni ci ar l a reacci n se aadi si mul tneamente
20pL de l a
muestraal a cubeta del amuestray20 , uL
def erri ci ani da potsi ca 0, 1Mal a
cubetadel bl anco.
La
di smi nuci n de absorbanci a a 550
ni n ( debi da al a oxi daci n del
ci tocromo
creduci do), sedetermi n ai nterval os de 10s durante
1 mi na37C
en
unespectrof otmetroHi tachi U-2. 000.
La acti vi dad espec f i ca del enzi ma
( S. A. ) se cal cul uti l i zando l a
si gui entereacci n:
S. A. = ( k x
[ ci t . C] )/ [ enz]
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Materi al yMtodos
donde k =
2, 3
x
l og (A0/ Al mi n), expresada en mi n- 1, [ c i t . C] es l a
c onc entrac i nde c i toc romoc
quese aadi (0, 08M), y[ enz] es l ac anti dadde
enzi ma
quese aadi expresadaenmgdeprote na.
Los resul tados se expresaronc omoac ti vi dad
espec f i c a (S. A. ) por mg de
prote na.
3. 6
. 2.
Ac ti vi dad
c i tratosi ntetasa
El enzi ma c i trato si ntetasa se enc uentra en
l a matri z mi toc ondri al ,
pertenec eal c i c l o deKrebsy
es c odi f i c adopor el ADNnuc l ear. Seuti l i za
c omo
marc ador
de l a c apac i dad metabl i c a de l as
mi toc ondri as y tambi n es
uti l i zado c omo i ndi c ador de l a c anti dad
de mi toc ondri as presentes en
l a
preparac i n(Morel
et al . , 1995) .
Sedetermi n mi di endol a
f ormac i nde c i trato aparti r de
ac eti l - CoA y
oxal ac etato(OAA),
segnel mtododesc ri topor Wi l l i ams
et al . (1998) .
La absorbanc i a se mi di aadi endo
a una c ubeta: 0, 1 mL de
Tri s
0, 75mM, O, 1mLdeDTNB1mM,
0, 1 mLdeTri tnX- 100 1%,
0, 05 mLdeAc eti l
CoA8, 6mM, 0, 58 mL
deaguadesti l aday0, 02 mL
demuestra. Lareac c i nse
i ni c i
c onl a adi c i nde 0, 05 mLde OAA
l OmMal a c ubeta de l a
muestra,
f rente aunbl anc oquenol l evaba
OAA.
El
aumentode absorbanc i a a412 nm
se si gui a i nterval os de
30s
durante3mi nutosya30C,
enunespec trof otmetro
Hi tac hi U- 2. 000.
La c onc entrac i n de ac ti vi dad
c atal ti c a ([ A. C. ] ) del enzi ma
c i trato
si ntetasasec al c ul segnl a
si gui entereac c i n:
[ A. C
. ] =AbsxVxc emxdxtxv
dondeAbs es l a absorbanc i aati empo
3 mi nmenosl aabsorbanc i aati empo
0,
Ves el vol umenf i nal del
ensayo (1 mL), c emes el c oef i c i ente de
exti nc i n
mol ar
dec i tratosi ntetasa, des el pasodel uz de
c ubeta (1Omm), t es el ti empo
enmi nutosyves el vol umendemuestraaadi do
.
Losresul tados
seexpresaronc omoc onc entrac i n
deac ti vi dadc atal ti c a
por mgdeprote na.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Materi al yMtodos
3. 6. 3. Determi naci n
demal ondi al deh do
El mtodo del ci doti obarbi tri co (TBA) se
uti l i za para anal i zar l a
peroxi daci n l i p di ca. En este proceso,
unode l os productos f i nal es es el
mal ondi al deh do (MDA), un compuesto que reacci ona con
el ci do
ti obarbi tri co (TBA) dandol ugar al a
f ormaci ndeuncompuestocol oreado
rosayqueti ene unmxi mode absorci na535 nm. Se
si gui el protocol o
descri topor Fragaet al . (1988) .
A50pLdehomegeni zado
(deh gadoori n) sel eaadi 75pLdeci do
f osf ri coy25 pL deTBAy5 pL dehi droxi tol uenobuti l ado
(BHT), se agi t en
vrtex , se i ncub durante60mi nutosa
95C, transcurri doese peri odode
ti empose aadi 150uL
desol uci nneutral i zante (compuestapor 500 AL de
NaOH 1Ny4, 5
mL de acetoni tri l o), se centr f ugo ensegui da
a 13000 rpm
durante 5 mi nutos a4 Cyse i nyectaron
20 pL del sobrenadante en el
cromatgraf ol qui dodeal taresol uci n (HPLC) (el f l uj o
f uede0, 4 mL/mi nyel
detector UNa535nm) .
Laconcentraci nde
MDAdel as muestrassecal cul por
extrapol aci n
a
unacurva patrn de concentraci ones
conoci das de MDA(desde 10 mM,
hasta0, 1 mM) . Laf asemvi l uti l i zadaf ue
acetoni tri l o/agua(70/30%)
Enl a f ase
de i ncubaci n durante60mi nutos a95
C se producel a
hi drl i si s del osl i poperxi dos, conl a consi gui entel i beraci n
deMDA. Este se
conj ugacon 2 mol cul as de TBA, por l o
que se determi nael aductoMDA
(TBA) 2 como
ndi ce deperoxi daci nl i p di ca. Si nembargo,
este aductoes muy
i nestabl e, por l o quehayquecentri f ugar y
pi nchar en el HPLC
rpi damente
tras aadi r
l a sol uci n neutral i zante. Por otra
parte, l a adi ci n del
anti oxi dante BHTprevi enel a autooxi daci n
de
l a
muestradurantel af ase de
cal entami ento, evi tandoobtener val ores
deMDAarti f i ci al menteal tos.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Materi al
yMtodos
4. Mtodos
estadi sti cos
Seha
real i zadounaestad sti ca descri pti va
de cadaunade l as vari abl es
cal cul ando l os
parmetros caracter sti cos: medi a, desvi aci n
t pi ca. La
comparaci n
de l as medi as normal es
entre dos grupos experi mental es se
ef ectu por el test del at de
Student ( de dobl ecol a) .
Para l os
resul tados expresados comoporcentaj es
se real i z el estudi o
estad sti co medi ante l a comparaci n
de dos proporci ones, apl i cando l a
Correcci ndeYates
.
Se consi der
que no hab a si gni f i caci n
cuando el val or de l a
probabi l i dad de si gni f i caci n ( p) f ue
mayor de 0, 05. Se di o el val or
de
"si gni f i cati vo" parap<0, 05
.
Todos
l os datos f uerontratados
conl os programas i nf ormti cos
SPSS,
Excel 2000. Todos
l os grf i cos f ueronobteni dos graci as
al os programas
i nf ormti cos
Si gmaPl ot 3. 0 yExcel 2000.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Volver al ndice/Tornar a l'ndex
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
RESULTADOS
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
1.
Model odeenvej eci mi entocel ul ar i nvi tro
El pri nci pal obj eti vo de esta i nvesti gaci n f ue
demostrar el ef ecto
protector de
di f erentes compuestos anti oxi dantes (vi tami naC, N- aceti l - ci ste na
yti oprol i na) sobrecl ul as del EPRb expuestas aestrs oxi dati vo
. Para el l o, se
val or el gradodedaocel ul ar determi nando
l a concentraci ni ntracel ul ar de
ATPyl os ni vel es
de
LDH
l i berados al medi opor estas cl ul as.
1. 1.
I nducci ndedaocel ul ar por peroxi ni tri to
Seobserv
unadi smi nuci ndosi s- dependi entedel a concentraci n
de
ATPcuandosei ncubaronl as cl ul as
conPNcomoseobservaenl a Fi gura 14.
As , cuando
l a concentraci ndeperoxi ni tri touti l i zada f ue50
MMel conteni do
enATPcel ul ar di smi nuy al a mi tadmi entras
quecuandol a concentraci n
f ue100pMl a concentraci ndi smi nuy
un94%yaparti r de250l i Mnovari
l a concentraci n
deATP.
140
-
120-
100
-
Control
50PM
100PM
250pM
500PM
Resul tados
Fi gura 14. EFECTODELPEROXI NI TRI TO
SOBRELOSNI VELESDEATP.
Ef ectodel PNsobrel a concentraci n
i ntracel ul ar deATPencl ul as deEPRb tras 1 h
de i ncubaci n con di sti ntas
concentraci ones de PN. Las cl ul as se recogi eron
medi ante tri psi ni zaci n, y se determi n l a
concentraci n i ntracel ul ar de ATP
medi ante bi ol umi ni scenci a, comose i ndi ca
enl a secci nMateri al yMtodos. Las
barras representanl a medi aD. E. detres experi mentos
real i zados por tri pl i cado((* )
=p <0, 05al comparar conel grupocontrol , si nPN) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Resul tados
Secomparel ef ecto deNAC( anti oxi dante) con
dosi nhi bi dores dePARP
( 3-ami nobenzami da( 3-AB) y
ni coti nami da ( NM) ) uti l i zando PN( 50 y 10011M)
comoi nductor del dao
cel ul ar.
Comoseobservaen l aFi gura15, al uti l i zar PN
50 ^ni NACni 3-AB
y NM0, 1 mMmostraron ef ecto protector
sobre l a concentraci n de ATP
i ntracel ul ares
mi entras que s se observ unaprotecci n
si gni f i cati va en
presenci a de concentraci ones
superi ores ( 1 y 10 mM) , observndose
el
mxi mo
ef ectocon NAC10 mM.
a
Q
NAC
3-AB
NM
Fi gura15. EFECTODELOS
I NHI BI DORESDEPARPYNACSOBREEL
ATP.
Se determi n el ef ecto protector de l os
i nhi bi dores de l a PARPy NACsobre
l a
concentraci n i ntracel ul ar de ATPen
cl ul as de EPRb. Las cl ul as se prei ncubaron
con di f erentes
concentraci ones def rmacosy f ueron
expuestasaPN50 j i Mdurante
1
hora, de acuerdoal protocol odescri to en l a secci n
deMateri al yMtodos. ( ( * ) _
p<0, 05
f renteal grupocontrol si n PN; ( #) =
p<0, 05 f renteaPN50 j i M) .
Si n
embargo, en presenci a de
PN100 MM, NAC1 mMf ue
i nef i caz
mi entras queNAC10 mMs
mostr un ef ecto protector f rente
aPN100 j I M
( Fi gura 16) . Al
prei ncubar l as cl ul as con
3-AB se observ unaacci n
protectora
aambasconcentraci ones 1 y
10 mMmi entras queNMal i gual
que
p
0, 1
MM
120
1
MM
a10MM
100
80 1 1 1
# #
# a Z#
60
0
T
n
t
40 -
r
0
P
1
N
20
5
0
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
NACsl o mostr ef ecto protector al a concentraci n ms al ta, o sea 10 mM.
Como se observa
enl a
Fi gura
16, l os f rmacos al a dosi s ms al tapreservaron
el conteni do de ATPi ntracel ul ar pero nunca
al canzaron l os val ores del control .
120
100
80
a.
60
40
20
0
c
0
n
t
r
O
1
91
Control NAC 3-AB
NM
Resul tados
M1 mM
M10MM
Fi gura 16. EFECTO
PROTECTORDELOSI NHI BI DORESDEPARPYNACSOBRE
LOSNI VELESI NTRACELULARES
DEATPyPN100
Ef ecto protector de l os i nhi bi dores
de PARPyNACsobre l aconcentraci n i ntracel ul ar
de ATPen cl ul as
de
EPRb
. Cl ul as prei ncubadas condi f erentes concentraci ones de
f rmacos e i ncubadas
durante 1 hora con PN100 pM. Las barras representan l a
medi aD. E. de tres experi mentos
real i zados por tri pl i cado. ( ( #) =p<0, 05 al comparar
conel grupo control si n PN; ( * ) =p
<0, 05f rente aPN100pM) .
Al i gual que en
l as anteri ores condi ci ones de estrs oxi dati vo, tambi nse
estudi l a
vi abi l i dad cel ul ar determi nando l a secreci n de LDHal medi o de
cul ti vo. El peroxi ni tri to tambi n reduj o l a vi abi l i dad cel ul ar
de f ormadosi s
dependi ente
i ncrementando si gni f i cati vamente l os ni vel es de LDHsecretado al
medi o conf orme aumental adosi s de PNl l egando aal canzar ni vel es del
100%
de
acti vi dad
LDH
en el medi o de cul ti vo con l a dosi s ms al ta de PN( 5001uM)
como se observaenl a Fi gura17.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Resul tados
80
I
70
60 -
- Uz-
Q
50
b
n
- C 40
10 - .
D 0
1
r
C ontrol
50 PM
100PM
250uM
500
PM
Fi gura17. EFEC TODEPNSOBRELAVI ABI LI DAD C ELULARDELEPRb.
Lavi abi l i dad cel ul arse determi nmi di endol a acti vi dadLDHsecretadaal medi ode
cul ti vo por cl ul as de EPRb i ncubadas durante unahora con concentraci ones
creci entes
de
PN,
segn el protocol o
descri to
en l a
secci n
deMateri al yMtodos. Las
barras representan l amedi aD. E. de tres experi mentos real i zados portri pl i cado, ( * )
=p<0, 05vs control .
Seestudi el ef ectodel osi nhi bi dores dePARPyNAC sobre
l avi abi l i dad
cel ul ar. Para el l o se prei ncubaron l as cl ul as con l a dosi s ms al ta de
estudi ada anteri ormente ( 10 mM) , que mej or
ef ecto present en l a
preservaci n de l os ni vel es de ATP, y posteri ormente sei ncubaron con PN
501i M. Se observ unadi smi nuci n si gni f i cati va
de l a acti vi dad LDHen el
medi odecul ti vo
con l os tres f rmacos si endoNAC el quemsse aproxi m a
l osni vel es control ( Fi gura18) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Resultados
Fi gura
18. EFECTODELOS I NHI BI DORES DEPARP YNACSOBRE
LA
VI ABI LI DAD
CELULAR.
Seestudi el
ef ecto protector delos i nhi bi doresdelaPARP(3-ABy NM) y
del
anti oxi danteNAC
sobrelavi abi li daddelasclulasdeEPRb expuestasaPN
50PM
durante1 hora. El ef ecto
sedetermi n mi di endolaacti vi dadLDHsecretadaal
medi o
por clulas deEPRb si gui endoel
protocolodescri toen la secci n deMateri al y
Mtodos. Lasbarrasrepresentan la
medi a
D
. E
.
detresexperi mentosreali zadospor
tri pli cado,
(* ) =p<0, 05vscontrol; (#) =p<0, 05vs50pMPN.
50
1
45-
40-~
i
35
x
30-
Q
x
25
Q
- 20-
15 -'
-u
.
_.
10
*
#
5
1
El F-1
0
Control 50PM 50uM+3- 50pM+NM 50
AB
uM+NAC
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
ResuI t ados
1. 2. I nducci nde est rs oxi dat i vo por agent es qu mi cos
Cuando el est rs oxi dat i vo f ue i nduci do por ascorbat o e i n f erroso
(Asc/ Fe2+), l a concent raci n de ATPno vari conrespect o a l as condi ci ones
cont rol , al i ncubar l as cl ul as cont ampnHepes (Fi gura
19), mi ent ras que, s
se observ un descenso en l a concent raci n de ATP al i nduci r hi poxi a
medi ant e i ncubaci nconol i gomi ci na yci do i odoact i co (OL- I AA), al i gual que
cuando se i nduj o gl ucopeni a medi ant e exposi ci n
al ci do i odoact i co (I AA) y
cuando
se i ncubaron l as cl ul as en presenci a de compuest os capaces de
i nduci r ef ect os pat ol gi cos (OL- CN- I AA) si mi l ares al os de l a i squemi a.
U
0
0
0
N
W
.
F-
0, 45
0, 40
0, 35 -
0, 30-
0, 25 -
0, 20-
0, 15
0, 10
0, 05 -
0, 00-
Cont rol Asc/ Fe OL- CN I AA OL- CN- I AA
Fi gura 19. NI VELESDEATPENCONDI CI ONES
DEESTRSOXI DATI VO.
Se det ermi nl a concent raci n
i nt racel ul ar de ATPencl ul as del EPRb somet i das a
est rs oxi dat i vo, hi poxi a, gl ucopeni a o i squemi a
si gui endo el prot ocol o experi ment al
descri t o enl a secci nde Mat eri al yMt odos. Las barras
represent anl a medi a D. E.
de t res experi ment os real i zados por t ri pl i cado. (* ) p<
0, 05 con respect o a l os
resul t ados obt eni dos encondi ci ones cont rol .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Todos l os anti oxi dantes
uti l i zados protegi eron al as cl ul as someti das
a
hi poxi a
mi entras que tras l a exposi ci n a gl ucopeni a
sol o NACf ue capaz de
proteger f rente al daocel ul ar, TP
previ no l a di smi nuci n de l a concentraci n
de ATPsi n al canzar l os ni vel es
del control , mi entras que VCf ue
i nef i caz .
Por
otra parte, en el model o
de
i squemi a
qu mi ca, VCy NAC
protegi eron si gni f i cati vamente
l a di smi nuci n de l os ni vel es
de ATP si n
al canzar l os ni vel es del control ( 0, 32
0, 09) ( Fi gura 20) , mi entras queen este
casoTPf ue i ncapaz de mantener
l os ni vel es de ATP.
0, 7
0, 6
~, 0, 5
U
O
O
0, 4
N
0
El Estrs
a+VC
"+NAC
E+
TP
Control OL-CN I AA
0L-CN-I AA
Resul tados
Fi gura20. EFECTODE LOSANTI OXI DANTES
SOBRE LOSNI VELESDE ATP.
Ef ecto
de l a vi tami na C( 300 pM) , N-aceti l -ci ste na
( 10 mM) yti oprol i na ( 3 mM) sobre
l a concentraci n
i ntracel ul ar de ATP en cl ul as de EPRb
someti das a hi poxi a,
gl ucopeni a o i squemi a
qu mi ca, determi nado segn est descri to
en l a secci n de
Materi al yMtodos
. Las barras representan l a medi a D. E .
de tres experi mentos
real i zados por tri pl i cado. ( #) =
p<0, 05 f rente control , ( * ) =p<
0, 05 comparado con el
estrs correspondi ente.
La vi abi l i dad
cel ul ar se determi n mi di endo
l a secreci n al medi o de
cul ti vo
del enzi ma LDH, i ndi cador
de l a i ntegri dad de l a membrana
pl asmti ca. Como se puede
observar en l a Fi gura 21, se
produj o un marcado
aumento
de l a acti vi dad de este enzi ma en el medi o
de cul ti vo conrespecto a
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Resul tados
l as condi ci ones control cuando l as cl ul as se i ncubaron en condi ci ones
de
hi poxi a
e
i squemi a,
mi entras queen l as dems condi ci ones experi mental es
sl o se observ un l i gero aumentono si gni f i cati vo del al i beraci n al medi ode
esteenzi ma.
x
Q
U
Q
v
0
14-
12 -I
10
$.
6-
4
2
Control Asc/Fe OL-CN I AA
OL-CN-I AA
Fi gura21.
DETERMI NACI NDELAVI ABI LI DADCELULAR.
Sedetermi n
l aacti vi dad LDHsecretadaal medi opor l ascl ul as de EPRb someti das
aestrs oxi dati vo,
hi poxi a, gl ucopeni a e i squemi a, si gui endoel protocol o
descri to en
l a secci n de Materi al y
Mtodos. Las barras representan l a medi aD. E. de tres
experi mentos real i zados por tri pl i cado. ( * ) p<
0, 05 vs control .
Como
yase hamostrado anteri ormente, el i ncremento de
l a secreci n
de LDHal medi o de cul ti vo se produj o
en l as condi ci ones experi mental es de
hi poxi aqu mi ca
e
i squemi a
( Fi gura21) . Cuandol as cl ul as f ueron
pretratadas
con
l os anti oxi dantes durante dos horas antes de ser
someti das ahi poxi a
qu mi ca, se observ
como el i ncremento de LDHen el medi o di smi nuy
si gni f i cati vamente cuando l as cl ul as se trataron
con NACyTP( Fi gura22) ,
si endo TPel
anti oxi dante quemantuvol os ni vel es de secreci n ( 2, 98
0, 59)
msproxi mos al os ni vel es del as condi ci ones control ( 3, 34
1, 27) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Mi entras que en l a condi ci n de gl ucopeni a ( I AA, l a secreci n de LDH
al medi o de cul ti vo no i ncrement si gni f i cati vamente) , al i ncubar l as cl ul as
conVC, di smi nuyeron si gni f i cati vamente l os ni vel es de secreci n al medi o, TP
l os
mantuvo si mi l ares a l as condi ci ones control y de gl ucopeni a y
paradj i camente NACi ncrement si gni f i cati vamente l os ni vel es de secreci nal
medi o de cul ti vo.
Enl a condi ci n
de
i squemi a
se produj o unl i gero aumento si gni f i cati vo
en l os ni vel es del enzi ma secretado al medi o de cul ti vo ( Fi gura 21) que
di smi nuyeron si gni f i cati vamente cuando
l as cl ul as se prei ncubaron conTP,
al canzando l os val ores de l as condi ci ones control ( 3, 46 0, 26 y
3, 34
1, 27
respecti vamente) .
Cuando el anti oxi dante f ue VC se produj o un l i gero
descenso si gni f i cati vo de l a secreci n al medi o de LDHcon
respecto a l os
ni vel es obteni dos en presenci a de OL-CN-I AA
mi entras que NACvol vi a
produci r uni ncremento
del a secreci nal medi o de LDH( Fi gura22) .
C) Cd
18
16
0
Estrs
"VC
"NAC ETP
OL-CN
I AA OL-CN-I AA
Resul tados
Fi gura22. EFECTODELOSANTI OXI DANTESSOBRELAVI ABI LI DADCELULAR.
Se determi n l a concentraci n de LDHsecretado al medi o de cul ti vo en cl ul as
de
EPRbi ncubadas en ausenci a o presenci ade anti oxi dantes: VC300^NACl OmMy
TP3mMy someti das adi f erentes condi ci ones de estrs oxi dati vo hi poxi a, gl ucopeni a
o
i squemi a qu mi ca,
si gui endo el protocol o descri to en l a secci n de Materi al y
Mtodos. ( * ) =p<0, 05vs control ( 3, 34 1, 27) y ( #) =p<0, 05f rente al a condi ci n de
estrs oxi dati vo
.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Resul tados
1. 3. I nducci ndedaocel ul ar por
hi poxi a
Una
exposi ci n prol ongada abaj as concentraci ones
de ox geno i nduce
un
dao en l as cl ul as del EPRb. Como
se puedeobservar en l a Fi gura
18,
cuando l as cl ul as se i ncubaron
durante 72 horas en medi o
de cul ti vo si n
suero
f etal bovi no ycon unaconcentraci n
de 02 al 3 %se produj o
una
di smi nuci n si gni f i cati va
en l a concentraci ni ntracel ul ar
de ATPcomparada
conl os
ni vel es obteni dos enl as condi ci ones control
( 21 %de02) .
Como se observa en
l a Fi gura 23 l a concentraci n
de ATP f ue
preservada sol amente cuando
l as cl ul as f ueron tratadas
con ti oprol i na que
mantuvo l a
concentraci n de ATPprxi ma
a l a observada en condi ci ones
control .
v
0
C
N
3
1
a
h
Q
`"
u, , : >
0
T
T
T
21%02 3%02
3%02+ 3%02+
3%02+
VC NAC
TP
Fi gura23.
EFECTODELOSANTI OXI DANTES
SOBRELOSNI VELESDEATP.
Conteni do
i ntracel ul ar deATPen cl ul as de EPRb
i ncubadas durante 72 horas
en
normoxi aehi poxi a, yen
presenci ade anti oxi dantes.
Se si gui el protocol o descri to
en l a secci n de Materi al y
Mtodos. Las barras representan
l a medi aD. E. del
conteni do
i ntracel ul ar deATP, de tres
experi mentos real i zados por tri pl i cado
( ( *) =p<
0, 05comparado
conel control ennormoxi a, ( #) =p<
0, 05vs control hi poxi a)
.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Se determi n l a vi abi l i dad cel ul ar tras someter a
una
hi poxi a
prol ongada
a l as cl ul as mi di endo
l os
ni vel es
de LDHsecretados al medi o de cul ti vo.
Se
observ un i ncremento si gni f i cati vo de l a
acti vi dad LDHen condi ci ones de
hi poxi a al comparar con l os
resul tados obteni dos en condi ci ones control
(Fi gura 24) .
Cuando l as cl ul as
se i ncubaron en hi poxi a yen presenci a
de l os
anti oxi dantes
se observ una reducci nsi gni f i cati va en l a
l i beraci n al medi o
de LDH, si endoel tratami entoconti oprol i na el
ms ef ecti vo.
7
x
Q
3
- o .
U
_ - C
<
2
\
y~ O
0
21%02 3%02
3%02+3%02+3%02+
VC
NAC TP
Resul tados
Fi gura 24. EFECTODELOSANTIOXIDANTES
SOBRELAVIABILIDADCELULAR
Se determi nl a acti vi dadde LDH
secretada al medi ode cul ti vo por cl ul as de EPRb
i ncubadas en normoxi a (21%de
02)
o en hi poxi a
(3%de 02) , yen presenci a de
anti oxi dantes, si gui endoel protocol o experi mental descri to
en l a secci n de Materi al
yMtodos. Las barras representan l a medi a D. E. del
porcentaj e de LDHtotal
secretado
al medi o de cul ti vo en tres experi mentos real i zados por tri pl i cado
. (
(#) =
p<0, 05 vs control normoxi a;
(* ) =p<0, 05 vs control hi poxi a) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Resul tados
1. 4. I nducci ndeapoptosi spor estrs oxi dati vo
1. 4. 1. Detecci nmedi anteti nci nconHoechst
En el presente estudi o se real i z
l a ti nci n Hoechst en l os di f erentes
model os de i nducci n de estrs oxi dati vo en l as cl ul as de EPRb para
comprobar si i nduc anapoptosi s.
Los resul tados se i ndi canen
l as
f i guras
presentadas aconti nuaci n. En
l as cl ul as de EPRbl a apoptosi s no muestra el t pi co patrn de cuerpos
apoptti cos, en el queel ncl eo se di vi de en pequeos f ragmentos dandol a
i mpresi ndequehay
ms de unacl ul a. Cuandol as cl ul as deEPRbsuf ren
apoptosi s muestran una reducci n del tamao del ncl eo y una mayor
f l uorescenci aque
encondi ci ones normal es (Osborneet al . , 1997) .
Enl a Fi gura 25se muestral ati nci ndel os ncl eos de cl ul as que se
i ncubaron en medi o de cul ti vo control . Como
se puede apreci ar, l a
f l uorescenci a es
muytenuey el aspecto de l os ncl eos es el de unacl ul a
normal .
Fi gura25. MORFOLOGADECLULASDEEPRb. TI NCI ONDEHOECHST
.
Aspecto de l as cl ul as de EPRbsi n ni ngn ti po de estrs oxi dati vo
. Las cl ul as se
i ncubaron en medi ode cul ti vo yposteri ormente
se
real i z
l a ti nci n con Hoechst
33258(0, 05pg/ mL) durante25mi n. , si gui endoel
protocol odescri to enl a secci nde
Materi al yMtodos. Barra=20 pm
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Resul tados
Cuando l as cl ul as
se i ncubaron conPN50 MM
se observ apoptosi s
como
se muestraenl a Fi gura
26, mi entras queconl as
concentraci ones ms
al tas estudi adas se produc a
unaumento dosi s
dependi ente de muerte por
necrosi s.
Esta apoptosi s i nduci da por PN
50MMf ue preveni da cuando
l as
cl ul as
sei ncubaronconNACl OmM
comoseobservaenl a Fi gura
27
.
14
. w
~
~
"
%
. " , *
f *
"
1.
" " %
t
a"
s
i
a
11
_%
"
.
i
9. l a
l o
Ma
41h
Fi gura26. INDUCCINDEAPOPTOSIS
CONPN50^
Estudi o morf ol gi co de l a apoptosi s
en cl ul as del EPRb medi ante ti nci n
con
Hoechst.
Las cl ul as del EPRtras 1 horas deexposi ci n
a50 gMPNf ueron tei das
si gui endo
el protocol o descri to en l a secci n
de Materi al y Mtodos. Las f l echas
bl ancas seal an
os ncl eos apoptti cos. Barra=10 um
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Resultados
+
- 9 !
`y*
_
w
0
Al
+
w
l
ya "
` r
_
) 1
o
`
rF
Fi gura27. EFECTOPROTECTORDENACSOBREPNSOPM.
Lasclulasseprei ncubaron durante1 horaconNAClOmMyposteri ormentef ueron
expuestasa
PN
50pMdurante1 horayseti eronsi gui endoel protocolodescri toen
lasecci ndeMateri al yMtodos
. Lasf lechasblancasi ndi canlosnucleosapoptti cos
ylasroj aslosncleosnoapoptti cos
. Barra=1 0
pm
En laFi gura28seobservalaausenci adeapoptosi sen lasclulasde
EPRb expuestas ahi poxi aqu mi ca, glucopeni ay
ai squemi aqu mi ca,
morf olog aquetambi n
seobserv cuandolasclulassei ncubaron con los
anti oxi dantesprevi amentealai ncubaci n conlosdi f erentesagentes
qu mi cos
i nductoresdel estrsoxi dati vo. Comosepuede
observar, lamorf olog acelular
novar acon respectoalade
lasclulascontrol, nosometi dasani ngn ti po
deestrs(Fi gura25) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Resul tados
Fi gura 28. EFECTODELA
HIPOXIAQUMICASOBRELAAPOPTOSIS
.
Las cl ul as se i ncubaron durante
15 mi n con OLCNpara someterl as
a hi poxi a
qu mi cayposteri ormentese f i j aron
con
PFA4%
paratei rl as segnel
protocol ode l a
Ti nci n
conHoechst descri to enMateri al y
Mtodos. Barra=20pm.
Como
se muestra en l a Fi gura 29,
en l as condi ci ones de hi poxi a
ambi ental (3%de
02
durante 72
horas) se observ apoptosi s.
Cuando l as
cl ul as se
prei ncubaroncon l os anti oxi dantes y
posteri ormente se expusi eron
aunahi poxi a
prol ongada (3%de
02
durante 72
horas) se observ que l os
anti oxi dantes proteg an f rente
al dao cel ul ar produci do por l a
exposi ci n a
l as baj as concentraci ones
de ox genocomoseobservaenl aFi gura
30, donde
el
anti oxi dante uti l i zado f ue VC, menos
ef ecti vo que NAC(Fi gura 31) . El
anti oxi dantemsef ecti vo
f ueTP comoseobservaenl a Fi gura
32.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Resultados
"
.
".
'
a
,
:
"
"
a
s,
""
".
"
a
4,
"
""
."
"
a
Z
l
"
a
--w
9
s
a .
!
,
" "
4
la %
o
Fi gura29. INDUCCINDE
APOPTOSISCONHIPOXIAAMBIENTAL.
Seobservalamorf olog adelas
clulasdel EPRbtrashaber si doi ncubadasdurante
72horasenunaatmsf era
al 3%deOz con medi odeculti vosi n SFB. Lasf lechas
sealanlosncleosapoptti cos.
Barra=10li m.
. -
-s
"
. l
9
a
"
4*
4
"
q.
" "
"
0
0
" a
a
"
e
111
.
-0
!
4
a
9
4
0
9
aI
a
a
a"
a
0
.
1P
&
"
"
" .
t !
"
""
"
a
" I
"r
"
a
0
4p
e#
4
"
"
"
"
"
0
Q> 0
#a
"
I
"
a 0
I
.
4b
r
e
41
!
.
o 0 a
%
"
a
"
1
"
0
, 1
"
,
Fi gura30.
EFECTODEVCSOBRELAHIPOXIAAMBIENTAL
.
Lasclulas
del EPRbseprei ncubarondurante2horasconVC
antesdeser expuestas
aunahi poxi a
ambi ental transcurri doel peri ododei ncubaci n
sereali z launci n
con Hoechst.Lasf lechasblancas
i ndi can los ncleosapoptti cosy
laroj aslosno
apoptti cos. Barra=
20Mm
.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
12
Fi gura
31. EFECTODENACSOBRELA
HIPOXIAAMBIENTAL
Lascl ul as se prei ncubaron conNACdurante
2 horas y posteri ormente se i ncubaron
durante 72
horas en unaatmsferaal 3%
02 . Transcurri do l a i ncubaci n se fi j aron
con PFAal 4%y
se real i z l a Ti nci n de Hoechst
si gui endo el protocol o descri to en
Materi al y
Mtodos. Las fl echas bl ancas seal an
l os ncl eos apoptti cos y l as roj as
l os ncl eosno apoptti cos
. Barra=20, um.
Resul tados
Fi gura32. EFECTODETPSOBRE
LAHIPOXIAAMBIENTAL
Las cl ul as se prei ncubaron con
TPdurante 2 horas y posteri ormente
se i ncubaron
durante 72 horas en una. atmsfera
al 3%02 . Posteri ormente se real i zl a Ti nci n
de
Hoechst si gui endo el protocol o
descri to en Materi al y Mtodos. Las fl echas bl ancas
i ndi canl os ncl eos apoptti cosyl as
roj as l os ncl eosno apoptti cos. Barra=20pm.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Resul tados
1 . 4. 2. El ectrof oresi s
engel deagarosa
Despus de determi nar quti po
de estrs i nduce muerte cel ul ar por
apoptosi s medi ante
l ati nci nHoechst, seconf i rmaron
l os resul tados medi ante
real i zaci n
de el ectrof oresi s engel de
agarosa. Seuti l i zaron paraeste estudi o
l ahi poxi aambi ental yel PN
comoi nductores de apoptosi s.
Enl a Fi gura
33 se muestrael ef ecto de
di f erentes concentraci ones de
PN
sobre l as cl ul as del EPRb. La
f ragmentaci n del ADNse muestra como
una "estel a" que va
di smi nuyendo su i ntensi dad conf orme
aumenta l a
concentraci n de PN
si endo l a ms i ntensa l a cal l e 4
correspondi ente al a
concentraci n
de PN50 MM, en l as
cal l es 5 a7, en l as que se
representan
concentraci oes creci entes
de
PN,
di smi nuyel ai ntensi dad de
l aestel ayaquel a
muertees
por necrosi s ynoseproducef ragmentaci n
del ADN.
1
2 3 4 5
6 7 8
a
Fi gura
33. INDUCCINDEAPOPTOSIS
PORPEROXINITRITO.
Sedetect l a muertecel ul ar por
apoptosi s medi ante el ectrof oresi s en gel
de agarosa
si gui endo
el protocol oexperi mental
descri to enl a secci nde Materi al y
Mtodos. La
f ragmentaci n
del ADNse demuestrapor
unaestel a enl ascal l es correspondi entes
. 1
y8: Standard
de ADN(cDX174 DNA/ HAEIII) ;
2
:
Control negati vo (sol ventedel PN) ;
3:
0uMPN; 4,
5, 6y7: 50, 100, 250y500uM
de
PN,
respecti vamente.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Resul tados
Cuando l as cl ul as se i ncubaronconl os i nhi bi dores de l a PARPyel
anti oxi dante NACal a concentraci n10 mM,
se observ comodi smi nu al a
f ragmentaci n
del ADNextra do de estas cl ul as al i gual quesuced aenl a
ti nci ndeHoechst, comose observa
enl a
Fi gura34,
si endo NACy3-ABl os
quemayor
protecci nof recen( cal l es 6y4, respecti vamente) .
1 2 3 4 5 6
7 8
! aMW
Fi gura 34. EFECTODE LOS I NHI BI DORES
DEPARP YNACSOBRELA
APOPTOSI SI NDUCI DAPORPEROXI NI TRI TO.
Se determi n el ef ecto protector de l os i nhi bi dores de l a PARP( 3-ABy NM) yNAC
sobre l a muerte cel ul ar por apoptosi s i nduci da por PN50^segn
el prototocl o
experi mental descri to enMateri al yMtodos. Parael l o se real i z el ectrof oresi s
en
gel
de agarosa si gui endo el protocol oexperi mental descri to enl a secci nde Materi al y
Mtodos. La f ragmentaci n del ADNse demuestra por una estel a en l as cal l es
correspondi entes. Cal l es 1 y7: Standard de ADN( DX174 DNA/ HAEI I I ) ; cal l e 2:
Control
negati vo ( sol vente del PN) ; cal l e 3: PN50 pM; cal l e 4: PN50 pM
+
3-AB
l OmM, cal l e 5: PN
50
bi M
+
NM
l OmM, cal l e 6: PN50; uM
+
NACl OmM.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Resul tados
Encl ul as expuestas ahi poxi aprol ongada, sereal i z unael ectrof oresi s
en gel de agarosa, en el que qued ref l ej ado l a "t pi ca" estel a de l a
f ragmentaci ndeADNenl a cal l e 3(3%02) no observadaenl acal l e 2 ( 21%
02) , comose observa enl a Fi gura
35,
tambi n
se qued ref l ej ado como l os
anti oxi dantes poseenunef ecto i nhi bi dor del a apoptosi s, si endoel msef i caz
TP(cal l e 6) , segui dodeNAC(cal l e 5) .
1 2 3 4 5 6 7
w
nnowmew
l qnnr~
WMEMEMW
Iwo
IMEN
rW
Fi gura 35.
EFECTOPROTECTOR DE LOS ANTIOXIDANTES SOBRE LA
APOPTOSISINDUCIDAPORHIPOXIA
AMBIENTAL.
Se determi n el ef ecto
protector de l os anti oxi dantes sobre l a muerte cel ul ar por
apoptosi s i nduci da por unexposi ci nprol ongada de baj as
concentraci ones de 02,
medi anteel ectrof oresi s engel deagarosasi gui endoel protocol oexperi mental descri to
enl a secci nde Materi al yMtodos. Cal l es 1 y7: StandarddeADN(DX174DNA/
HAEIII) ; cal l e 2: Control 21
%02; cal l e 3: Hi poxi a(3%02) cal l e 4: 3%
02 +
VC, cal l e
5: 3%
02 +
NAC
; cal l e6
:
3%02 +TP
.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
2. Model odeenvej eci mi ento
i nvi vo
2. 1 . Supervi venci ay
parmetros f i si ol gi cos del ratn
2. 1 . 1 .
Supervi venci adel a pobl aci n
En el grupo de ani mal es control ,
quea l o l argo de todo el ensayo
reci bi eronpi enso
demanteni mi ento, tuvol ugar unadi smi nuci n
progresi vade
l a supervi venci a parti endo deun
100%al i ni ci o del estudi o. As se observ
unadi smi nuci n
si gni f i cati va ( p<0, 05) del nmerode
ratones supervi vi entes
en el grupocontrol hasta
l l egar a un55%de supervi venci a al termi nar
el
estudi o.
Del os ratones queconsum an
anti oxi dantes, aquel l os cuyadi etaf ue
supl ementada conTP
ten an unamayor supervi venci a ( un 75 %) , y
l os que
reci bi eron NAC
en el pi enso mostraban l a mayor
l ongevi dad aunque, al
al canzar el l ti momesdel estudi o, l a
supervi venci adi smi nuy marcadamente
hastaunni vel del 60%.
Las medi as del os val ores del
porcentaj edesupervi venci adel os ratones
NACyTP,
desde50a62semanas, eransi gni f i cati vamente
di f erentes ( p<0, 05)
enrel aci nconel porcentaj edesupervi venci a
del os ratones control ( Tabl a 4;
Fi gura36) .
Resul tados
Tabl a4. EFECTODELOS
ANTIOXIDANTESSOBRELASUPERVIVENCIA.
Porcentaj ederatones supervi vi entes
enpobl aci ones control ( al i mentadas conpi enso
estndar) , yen l as que consum an pi enso supl ementado
con NACo TP. Entre
parntesi s: nmeroderatones supervi vi entes ( ( a) =p
<0, 05 vs
control 42semanas
;
( b) =p<0, 05 f rentecontrol mi smaedad) .
Edad
42 46 50 54 58
62
Control
1000, 01 95 0, 01 800, 01^
72, 5 3, 5^ 684, 01 55 0, 01^
( 20) ( 19) ( 16) ( 14) ( 13)
( 11)
NAC
1000, 01 900, 01KIb 900, 0P", 900, 0111b 90
0, 01' " l ) 67, 510, &, , "
( 20) ( 18)
( 18) ( 18) ( 18) ( 12)
TP
1000, 01 95 0, 01 85 0, 01, 111 82, 5
3, 5
: 1
. b
77, 5
41, b
75
0, 0P, b
( 20)
( 19) ( 17) ( 16) ( 15) ( 15)
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Resul tados
m
V
0
100
90
80
70
60
50
40
30
1
fNAC
tContro
32 34 36 38 40 42 44 4648 50 52 54 56 58 60 62
edad(semanas)
Fi gura36. EFECTO
DELOSANTIOXIDANTESSOBRELASUPERVIVENCIA.
Curvade supervi venci ade l a pobl aci n deratones hembraOF1, subdi vi di daentres
grupos
: control ,
NAC
y TP. Al i ni ci o del estudi o (a l as 32semanas) : n=20ratonespor
grupo, yal fi nal (62
semanas) :
n=11,
12 y 15, para l os grupos control , NACyTP,
respecti vamente
.
2. 1. 2. Pesocorporal
Al comparar l os
resul tados obteni dos enl os ratones control conl acurva
ponderal faci l i tada por l os sumi ni stradores de l a cepa estudi ada (OF1) se
observ que, a cada edad i nvesti gada, el peso de l os ratones de
nuestra
pobl aci nestabadentro
del
rango
normal . El pesoqueal i ni ci o del estudi o, a
l a
edadde32semanas, erade34, 8 3, 2g, se fuei ncrementandoprogresi va-
menteconel pasodel ti empo,
comose muestraenl aFi gura37. Al fi nal i zar el
estudi o, al aedadde62semanas, el pesoerade38, 44, 9g.
Enl os ani mal es del os
grupos queconsum ananti oxi dantes se observ
l ami smatendenci aal aumentodel pesocorporal . As enl os ani mal es
tratados
conNACel peso medi o al i ni ci o del
estudi o, de35, 31, 7 g, se fue el evando
hastaal canzar un
val or de38, 51, 7g, yenl os ratones queconsumi eronTP
el peso
aumentde 34, 63, 1ghasta39, 52, 8g(Fi gura37) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Aunque enl os
t r es gr upos deani mal es el
peso aument aba conel paso
del
t i empo, no se obser var oncambi os
si gni f i cat i vos enest e
par met r o debi dos
al a edado al consumo
deant i oxi dant es .
35
50
45- I
40
30
25
20
1
- El - NAC
* - Cont r o
32 34
36 38 40 42 44 46 48 50 52
54 56 58 60 62
edad (semanas)
Resul t ados
Fi gur a
37. EFECTODELOSANTI OXI DANTES
SOBREELPESOCORPORAL.
Cambi os
conl a edaddel peso cor por al de
l os r at ones. Ser eal i z unadet er mi naci n
cada dos
semanas del peso de cada uno
de l os ani mal es . En l a gr f i ca est n
r epr esent adas l as
medi as desvi aci n est ndar de l os
ani mal es cont r ol , NACy TP
(n=20por gr upo)
.
El envej eci mi ent o
se acompaa deunaument o
enl a het er ogenei dad de
l os r at ones enl o quer espect a al os val or es
del peso cor por al (como evi denci a
el aument o desudesvi aci n
est ndar , del as 32 al as
62 semanas de edad) .
Est e ef ect o
desor gani zador del envej eci mi ent o
no t uvo l ugar enl os ani mal es
queconsum anNACo TP, acausa
del a pr obabl eacci n pr ot ect or a de
est os
ant i oxi dant es sobr e l os
mecani smos homeost t i cos que pr eser van
mant eni mi ent o
de unpeso nor mal . Si se compar a
l a di st r i buci n de val or es
del peso cor por al en
i nt er val os de 5g, adi f er ent es edades,
se obser va queel
por cent aj eder at ones depeso el evado
(>40, 0g) del os t r es gr upos aument aal
envej ecer (Fi gur a
38) .
el
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Resultados
Como
seobservaen estaf i gura, lacurvadedi stri buci n del pesodelos
ratonescontrol se"aplana" consi derablementecon el pasodel ti empo, al
ganar pesolosratones, mi entrasqueen
lascurvasdelosgruposquef ueron
tratadosconanti oxi dantesdestacaun"pi co" queref lej aque
lahomeostasi sde
estosani malesescapaz
demantener el pesodelamayor adelosratonesen el
rangomsnormal paraestaedadde36, 0- 40, 9g.
Enresumen, el anli si s
estad sti codelosdatosrevelaqueel porcentaj e
deratonesque
seencuentran en el rangode31, 0- 35, 9 g essi gni f i cati vamente
mselevado( p<0, 05) alas32- 38 semanasqueauna
mayor edad
.
Por otra
parte, el porcentaj ederatonesqueseencuentran
en el
rango
de36, 9- 40, 9 g
esmayor ( p<0, 05)
paralosani malestratadoscon anti oxi dantesquepara
los
ani malescontrol.
0
Di stri buci npesos. 32- 38semanas
26- 30g 31- 35g
36- 40g 41- 45g >46g
peso( g)
20
_
Di stri buci npesos. 40- 46semanas
26- 30g 31- 35g 36- 40g 41- 45g >46g
peso( g)
Fi gura
38. EFECTODELENVEJECIMIENTOSOBRE
LADISTRIBUCINDE
PESOS
CORPORALES.
Serepresenta
ladi stri buci ndelospesoscorporalesdelosratones
devari asedades,
expresadacomo%
deratonesencadarangodepesos( n=20por grupo
decadaedad) .
( 26- 30 si gni f i ca
26, 0- 30, 9g, 31- 35si gni f i ca31, 0- 35, 9g, etc) .
100
80 -
- 0 Control
- NAC
- ~- TP
100
80
tControl
tNAC
-
f TP
60
-
G
60
0
v
40 -
s
40
20 -
20
0 -
26- 30g 31- 35g 36- 40g 41- 45g >46g
26- 30g 31- 35g 36- 40g 41- 45g >46g
peso( g) peso( g)
Di stri buci n pesos. 48- 54 semanas
Di stri buci n
pesos. 56- 62semanas
100
100
- Control
- Control
- NAC
80 -
- NAC 80
- ATP
N
f TP
C
60 - ,
y
C
60
0
m
u
40
4
40
O
X
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Comose puede
observar en l a Fi gura 39, nuestros
datos sugi eren que
el
estudi ode l a di stri buci n de l os pesos
corporal es de l os ratones en el rango
31, 0- 40, 9g proporci ona un senci l l o
mtodopara i denti f i car l os
ani mal es que
probabl emente
suf ren un envej eci mi ento
prematuro, es deci r muestran
una
edad bi ol gi ca mayor que
su edad cronol gi ca. Se puede
observar en esta
Fi gura que
el pi coen l a curva de l os pesos
de l os ratones supervi vi entes ( S)
corresponde al val or de 33- 34 g,
mi entras que l os pi cos de
l a curva de l os
ratones no- supervi vi entes
( NS) de l a mi smaedad
cronol gi ca ( 32- 38 semanas)
y l a de
l os ratones supervi vi entes,
cuya edad cronol gi ca era
aproxi mada-
mente dos meses mayor
( 40- 46 semanas) , coi nci den en
el val or de 35- 36 g.
0
0
Y
45
40
35 -
30
25
20
15
10
5
0
31- 32
33- 34 35- 36 37- 38
39- 40
peso( g)
Resul tados
Fi gura 39. DISTRIBUCINDELOSPESOSCORPORALES
.
Curvas de di stri buci n de l os val ores
medi os del peso corporal de ratones
supervi vi entes
yno- supervi vi entes ( "prematuramente envej eci dos")
. Las edades de
determi naci n
del peso f ueron 32- 38 semanas y40- 46 semanas
para l os ani mal es
supervi vi entes ( n=38) y 32- 38
semanas para l os no- supervi vi entes ( n=34) . ( 31- 32
si gni f i ca31, 0- 32, 9g, etc) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Resul tados
Unacomparaci nde l a Fi gura 39 conl a Tabl a 5 sugi erequeel anl i si s
de l a di stri buci n de l os pesos corporal es i ndi vi dual es, que
hace resal tar l a
apari ci nde subgrupos de ratones de peso no- pti mo, puede ser unmtodo
ms sensi bl e, para detectar
l os ef ectos del envej eci mi ento (normal o
prematuro), queel examendel os val ores medi os del peso, que aunqueti enden
a aumentar (de 35, 4 a 36, 5 g enl os S, y de 36, 3 a 36, 9 g en l os NS), no
muestranal teraci ones estad sti camente si gni f i cati vas desde
l as 32 al as 46
semanas
deedad.
As de acuerdo conl o antes expuesto, l a Tabl a 6 muestra que
el
porcentaj e
de ratones control cuyo peso est comprendi doentre l os 30, 0yl os
34, 9 g(pesopti mo) es mayor enel grupodel os supervi vi entes queenel del os
no- supervi vi entes
al o l argo del estudi o (conval ores al as 32- 38, 40- 46y48- 54
semanas de 73, 45 y27 %para l os S, yde 33, 22 y20 %para l os NS,
respecti vamente.
Tabl a5. EFECTODELENVEJECIMIENTO
SOBREELPESOCORPORAL.
Ef ectos del envej eci mi ento
yde l a supl ementaci ndel pi ensoconanti oxi dantes sobre
el peso medi o de l os ratones (engramos) . Entre
parntesi s: nmero de ratones en
cada grupo ((a) =p<0, 05 vs ani mal es de 32- 38semanas del mi smo
grupo) .
Edad
Supervi vi entes (S) No supervi vi entes (NS)
Semanas
Control NAC TP Control
NAC TP
I1 12 15 9 8 5
32- 38 35, 4 3, 6
35, 51, 6 35, 3 3, 2 36, 33, 2 35, 5 1, 9 35, 8 1, 5
40- 46 36, 54, 7 36, 21, 9 36, 52, 4 36, 94, 4
37, 52, 0 38, 02, 3
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Resul tados
Tabl a6. CALCULODEL
PORCENTAJEDEPESOENCADAEDAD.
Peso, a tr es edades, de cada
r atn contr ol del gr upo de super vi vi entes ( S) y
de l os
r atones
no- super vi vi entes ( NS) , queno
sobr evi vi er onhastael f i n del estudi o al as 62
semanas
. ( 30 si gni f i ca 30, 0- 30, 9 g, etc;
el aster i sco* i denti f i ca l os pesos pti mos) .
Entr e
par ntesi s: nmer o de ani mal es
en cada gr upo ( a= p<0, 05 con r especto al
val or
del por centaj e de pesos pti mos
en l os r atones super vi vi entes de l a mi sma
edad) .
Como
se i ndi ca en l a Tabl a 7, hay una r el aci n
di r ecta entr e el
por centaj e
de r atones contr ol que
muestr an un peso pti mo ( 30, 0- 34, 9 g) a
l as 32- 38semanas de edad
yel por centaj e de ani mal es quesobr evi ven hasta
el f i nal del estudi o.
Tabl a7. RELACION
PESOIDEALYSUPERVIVENCIA.
En esta tabl a se establ ece
l a r el aci n exi stente entr e el peso cor por al de l os r atones
contr ol a l a edad de 32- 38 semanas yl a
super vi venci a ( a=p<0, 05 con r especto al
por centaj e desuper vi vi entes de peso 30, 0- 34, 9g.
Edad
32- 38semanas 40- 46semanas
48- 54semanas
Gr upos
s( 11) NS( 9) s( 11) NS( 9)
s( 11) NS( 5)
32* 32* 32*
30* 32* 34*
33*
34* 33* 33* 34*
37
33*
34* 34* 35 34*
38
34* 35 34*
36 35 40
34*
35 34*
36 36 43
34* 37 35
38 37
34*
38 36
39 37
34*
40 36
42 38
38
42 37 44
39
39
48 43
44
48 46
PesoD. E. ( g)
35, 53, 6 36, 33, 2 36, 5 4, 7
36, 94, 4 37, 4 4, 1 38, 43, 4
r atones de
peso
"j uveni l "
73%
33%a 45% 22%a 27%
20%a
Edad( semanas)
Peso ( g)
Nototal de r atones %r atones
( SyNS)
super vi vi entes
30, 0- 34, 9 11
73
32- 38
> _ 35, 0 9
33'
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Resul tados
La comparaci ndel os datos de l a Tabl a 7
conl os de l a Tabl a 6sugi ere
quel a admi ni straci nde l os anti oxi dantes enel pi enso se asoci a conmayores
porcentaj es de
supervi vi entes enl os grupos de ratones de peso _35, Og a l as
32-38 semanas de edad, encomparaci n
conel porcentaj e de supervi venci a
que muestranl os ratones control
del mi smopeso. As , l os val ores son57%y
69%para el grupo NACy el grupo TP, respecti vamente, y sl o
33%para l os
control , condi f erenci as estad sti camente
si gni f i cati vas entre el porcentaj e de
supervi vi entes de cada uno de l os grupos de ratones
de peso no-pti mo
tratados conanti oxi dante yel porcentaj e
de supervi vi entes del grupo control
( p<0, 05) . En
l os ani mal es tratados con TP tambi n se observa
un ef ecto
benef i ci oso sobre el porcentaj e de supervi vi entes
de peso pti mo, segnl as
determi naci ones real i zadas al as
32-38semanas.
Tabl a8. EFECTODEANTIOXIDANTESSOBRE
ELPESOYENVEJECIMIENTO.
Porcentaj ederatones supervi vi entes ( S) en
rel aci n conel peso corporal yel consumo
de
anti oxi dantes. Entre parntesi s: nmero de ratones
en el grupo ( a=p<0, 05 con
respecto a l os porcentaj es
de ratones Scontrol de i gual peso, cuyos
datos se
muestranenl a Tabl a4) .
Los datos de
supervi venci a yde di stri buci n de
pesos corporal es del os
ratones tambi npuedenser obj eto de representaci n
grf i ca como hi stograma
ponderal tanto enestudi os
sobre l os mecani smos del envej eci mi ento
como en
l as
i nvesti gaci ones paraval orar l a ef i caci a
de l os tratami entos anti oxi dantes.
Comose puedeobservar en
l a Fi gura40, yde acuerdo con
l o antes expuesto,
el envej eci mi ento
se asoci a concambi os muymarcados
enl a di stri buci n de
l os pesos
corporal es del os ratones, condescenso
del porcentaj e deratones de
Edad
( semanas)
Peso ( g) NAC TP
30, 0-34, 9
67%
86%
a
32-38 ( 6)
35, 0
57%a 69%a
14
13)
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
peso "pti mo para l as expectati vas de supervi venci a"
( 30, 0- 34, 9 g) , del 70%
al 35%, desdel as 32- 38al as 48- 54 semanas deedad
.
Si mul tneamente, se
produce el correspondi ente aumento del
porcentaj e de ratones
de peso el evado ( >_35 g; "no- pti mos para
l a
supervi venci a") . Tambi n se puedeobservar
en l a Fi gura 40 queal a edad de
32- 38 semanas el porcentaj e
de ratones de peso pti mo ( ms "j uveni l ")
es
si gni f i cati vamente
mayor en el grupo de supervi vi entes
que en el de no-
supervi vi entes.
m
0
Y
.
ro
U
U
i .
O
Q.
100`%
90%
80%
70%
60`V>
50%
40%
30%
20%7
10%
0%
32- 38 40- 46 48- 54 32- 38
Supervi vi entes
Nosupervi vi entes
Resul tados
0<40
035, 0- 39, 9
E30, 0- 34, 9
Edad
Fi gura 40. EFECTOS DEL
ENVEJECIMIENTOSOBRELADISTRIBUCINDEL
PESOCORPORAL.
Con l a edad se producen cambi os en
l a di stri buci n de l os pesos de l os ratones
control ( n=11 para l os supervi vi entes, yn=9 para l os no- supervi vi entes
; edad en
semanas) . Exi sten di f erenci as si gni f i cati vas ( p<0, 05) entre l as vari as
edades para l os
porcentaj es de ratones supervi vi entes de peso 30, 0- 34, 9 g, as como paral os de peso
>35, 0g.
Por
otra parte, al a edad de32- 38semanas, el porcentaj e de ani mal es
de peso 30, 0- 34, 9 g ( peso pti mo)
de l os ratones no- supervi vi entes hab a
descendi do aunni vel
aproxi madamentei gual al del porcentaj e deani mal es
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Resul tados
de di cho peso que se encontraba en el grupo de supervi vi entes de
mayor
edadcronol gi ca
(48- 54 semanas) .
Comomuestral a Fi gura41, el hi stogramaponderal es muyadecuado
para presentar
en
forma
resumi da l os resul tados de l os estudi os con
anti oxi dantes en ratones cuyo peso est comprendi doen l os dos rangos de
peso corporal
i nvesti gados. Se observa que, mi entras l a admi ni straci n de
NACnoi nfl uye si gni fi cati vamente en el porcentaj e desupervi vi entes
depeso
30, 0- 34, 9 g, l a supl ementaci n
del pi enso conTPs quemuestra unefecto
benefi ci oso, aunque si n si gni fi canci a estad sti ca sobre el porcentaj e de
supervi vi entes quemostrabanestepeso pti moal aedadde 32- 38semanas.
Por otra parte, en l os grupos de
ani mal es de peso no pti mo l a
di ferenci a
entreel porcentaj e de supervi vi entes en el grupo control (41%)
y
en el queconsum aTP(68%) es estad sti camentesi gni fi cati va
(p<0, 05) .
ONS
pS
1000i 0
90%
80%
70%
60%
50%
40%
30%
20%
10%
0/ 0
Control
NAC TP Control NAC TP
30, 0- 34, 9 g
> 35, 0 g
Fi gura 41. EFECTODELOS
ANTIOXIDANTES SOBRELADISTRIBUCINDE
PESOSCORPORALES.
Supervi venci ade ratones control ytratados conl os
anti oxi dantes en l os dos rangos
depeso (pti moyno- pti mo), segnl os val ores del
pesocorporal determi nados al as
32- 38 semanas
de edad. Enl os ratones depeso nopti mo (>35, 0 g)
haydi ferenci as
si gni fi cati vas
(p<0, 05) entrel os porcentaj es de supervi vi entes en
el grupoderatones
control yen
el del os ratonessumi ni stradosTP.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
2. 1 . 3. Consumo
promedi odepi enso
Comoyase
ha
i ndi cado,
l ai ngestasecal cul pesandoal i ni ci odecada
per odo
de qui nced as el pi enso, quese col oca
enel comparti mentode cada
j aul aypesandode nuevo
el pi ensoquequedaenl a j aul aal os
qui nce d as.
Di vi di endo
el val or del a di f erenci a entre
ambascanti dades por el nmerode
d as ( o sea 15) ypor el
nmerodeani mal esal oj ados enl aj aul a, se
obti enen
l osval oresexpresados
enl aFi gura42.
El consumof uede4, 8 0, 7 gpor
ratn/ d a al i ni ci o del estudi oy
de
4, 6 0, 8g al termi narl o,
enel grupocontrol , mi entras
quel osval orescorres-
pondi entes a l os grupos tratados con anti oxi dantes
f ueron: 4, 4 0, 5g y
4, 7
0, 2gparaNAC; 3, 7 0, 6 gy4, 3
0, 3 gpara TP. Aunquel ai ngesta
de
pi enso al o l argo
del estudi o( 32 a62 semanasde
edad) f ue mayor enl os
ani mal es control ( que
reci b an pi enso estndar de manteni mi ento)
que en
aquel l os cuyadi etaf uesupl ementadacon
anti oxi dantes, l as di f erenci as entre
el consumoqui ncenal del os control y
el del os quei nger ananti oxi dantesno
al canzaron
si gni f i canci aestad sti ca.
42 44 46 48 50 52
54 56 58 60 62
edad( semanas)
Resul tados
Fi gura42. DI STRI BUCI NDELCONSUMO
DEALI MENTO.
Enl agrf i case representa l ai ngesta
medi ade pi ensopor ratn( gramos/ d a) de l os
ani mal escontrol y
del osqueconsum anNACoTP
.
( n=20 encadagrupo, al i ni ci odel
estudi o; n=11, 12y
15, respecti vamente, al termi nar) .
8
Control
- - E- - NAC
ATP
op
6~
5 - ~
P-
y, - . .
s5.
a
~*ss,
4 :
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Resul tados
2. 2. Efectos sobreparmetros
deconducta
2. 2. 1 . Pruebadecoordi naci nneuromuscul ar yvi gor
Los resul tados obteni dos medi ante esta prueba de "vi tal i dad"
demuestran que, al envej ecer, hay una prdi da progresi va del rendi mi ento
funci onal de l a pobl aci n expresado como el porcentaj e
de ani mal es que
real i zan correctamente
l apruebaenunmi nutoomenos.
En el grupo control , cuando se i ni ci el estudi o
a l a edad de 32
semanas el porcentaj e de ratones que
real i zaban l a prueba con xi to ( en
menos de un
mi nuto) erade un47%, yal termi nar el estudi o ( cuando
l os
ratones hab an al canzado l a edad de 62
semanas) di cho porcentaj e hab a
descendi do
al 11%.
70
60-
50-
40
30
20
10
0
32-38 40-46
48-54 56-62
edad( semanas)
tControl
-~NAC
f
TP
Fi gura 43. EFECTODEL
ENVEJECIMIENTOYLOSANTIOXIDANTESSOBRELA
COORDINACINNEUROMUSCULAR
.
Di smi nuci n con l a
edad del rendi mi ento en l a prueba de vi gor ycoordi naci n
neuromuscul ar ( mtodo de l a
cuerda ti rante) . Porcentaj e deratones quereal i zan el
test en560
segundos.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Resul tados
Tambi n
se observarondescensos con
l a edad en el nmero de ratones
de l os grupos NACyTPque
real i zabanconxi to el test de
l a cuerdati rante.
No obstante, l os datos
sugi eren quel os tratami entos
conestos anti oxi dantes
ti l i cos i nf l uyen
enl a prueba. As , l a
admi ni straci nde NACparece ej ercer
un
ef ecto
negati vo sobre l a
conducta de l os ratones,
mi entras que l a
supl ementaci n
del pi enso conTPprobabl emente
ej erce unef ecto protector
sobre l a prdi da de coordi naci n
neuromuscul ar y vi gor,
ya que exi sten
di f erenci as si gni f i cati vas
al comparar conel
grupo control (Fi gura43) .
Si nembargo, si expresamos
l os resul tados como
porcentaj e de prdi da
de rendi mi ento, asi gnando
el val or de 100%al os
resul tados obteni dos en l a
pri mera
prueba de cada grupo
(control , NAC, TP), es
obvi o que el
envej eci mi ento se acompaa
de una marcada
di smi nuci n de di cho
rendi mi ento. Como se
muestraenl a Fi gura43
(basada enl os mi smos
datos
del a Fi gura
43, converti dos en%
preservado de unval or i ni ci al
del 100%) l as
di f erenci as entrel os val ores del
rendi mi ento i ni ci al al as
32-38semanas yel
rendi mi ento
posteri or, al as 40-46, 48-54 y
56-62 semanas son
si gni f i cati vas
tanto parael grupo control
como paral os que
consumi eronNACo TP(p<0, 05,
atodas l as
edades menci onadas) .
Por otra parte, l a
admi ni straci n de TPaparentemente
preservaba el
rendi mi ento enl a prueba
de l a cuerdati rante (Fi gura 44) .
As l os val ores del
rendi mi ento en
l a cuerda ti rante al as
40-46, 48-54 y56-62 semanas
eran
mayores
para el grupo de ratones
al que se admi ni str TP, en
comparaci n
conel grupo control yel
grupo-NAC(p<0, 05al as tres
edades)
.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Resul tados
120
100
0
80
60 -
0
40
20
0
0 Control
fNAC
- A
- TP
32- 38 40- 46 48- 54 56- 62
edad(semanas)
Fi gura 44. EFI CACI ADELOSANTI OXI DANTESSOBRELACOORDI NACI N
NEUROMUSCULAR.
En
estaFi guraserepresentael rendi mi ento(coordi naci nneuromuscul ar yvi gor) en
l apruebadel a cuerda ti rante: efectos del a edadydel asupl ementaci n del adi eta
con l os anti oxi dantes
ti l i cos
NAC
y
TP. Exi sten
di ferenci as si gni fi cati vas (p< 0, 05)
entrel aedadde56- 62yl aedadde32- 38semanas
.
2. 2. 2. Efectossobrel aacti vi dadexpl oratori a
Segn estudi os anteri ores, el envej eci mi ento se acompaa de una
di smi nuci n de l a acti vi dad expl oratori a de l os ratones en l a prueba del
l aberi nto en formade T(De J uan, 1994; Del a Fuente
et al
. ,
1998a) . La
presente
i nvesti gaci n l ongi tudi nal tambi nmuestraundescensoconl aedad
en el nmeroderatones "rpi dos" control (quecompl etanen unti empo
<
20s
l aexpl oraci ndel brazodeentradaen
el l aberi nto) .
No obstante, segn se muestraen l a Fi gura 45, el %de
ratones
"rpi dos" en l apobl aci n
queal canz unal ongevi dadmayor de62 semanas
(ratones supervi vi entes) podr aaumentar del as 32- 38al as 40- 46
semanas,
debi do al entrenami ento, en l os
ratones queconsumen NAC TP, mi entras
queeste
efecto posi ti vo del entrenami entonotendr al ugar en l os ani mal es
control .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Noobstante, l os cambi os
queti enen l ugar en este
parmetrodesde l as
32-38
a l os 56-62 semanas no son
estad sti camente si gni f i cati vos ni
en el
grupocontrol ni en l os tratados
conl os anti oxi dantes.
100
90 -
80
70-
40
30 -'
20
l o-
0
Resul tados
0 Control
-*-
NAC
f TP
32-38
40-46 48-54
56-62
edad( semanas)
Fi gura 45. EFECTOS
DEL ENVEJECIMIENTO SOBRE
LA ACTIVIDAD
EXPLORATORIA.
En esta f i gura se
representan l os ef ectos del entrenami ento
y del envej eci mi ento
sobre l a acti vi dad expl oratori a
espontnea en l a prueba del l aberi nto
en f orma de t
porcentaj e de ratones "rpi dos" ( que real i zan
el test conxi toen un ti empo520s) .
Como compl emento
a l a i nvesti gaci n anteri or, de entre
l a pobl aci n
i ni ci al de
ratones se i denti f i c el conj unto de ani mal es
que no sobrevi vi
durante el ti empo
del estudi o, es deci r que no
sobrepas l as 62 semanas de
edad( al que, comoya hemos
i ndi cado, desi gnamos "grupo nosupervi vi ente"
o
de "escasa
l ongevi dad") yse determi naron l os
porcentaj es de ratones "rpi dos"
que este grupoconten a al as di versas
edades i nvesti gadas.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Resul tados
Si se comparanl as Fi guras 45y46, al as edades de40 a62semanas
hab aunporcentaj emayor deratones capaces decompl etar l a expl oraci ndel
l aberi nto en <_20 segundos ( ratones "rpi dos") en l a pobl aci n"l ongeva" que
en l os quemostraron unamortal i dad prematura ( p<0, 05) . Estas di f erenci as
entreel porcentaj e de "ratones
rpi dos" presentes enl a pobl aci n
"l ongeva"
y
en l a de "escasa l ongevi dad" de cada grupo ( control , NAC, TP) son
estad sti camente
si gni f i cati vas
al os 48-54
y
56-62semanas ( p<0, 05) .
100
90
80
70
60 -
50
40
30
20
l o-
o-
0 Control
f NAC
-*- TP
32-38 40-46
48-54 56-62
edad( semanas)
Fi gura 46. EFECTO DE LOS
ANTIOXIDANTES SOBRE LA ACTIVIDAD
EXPLORATORIAENRATONES DEESCASA
LONGEVIDAD.
Serepresentanl os ef ectos
del envej eci mi ento ydel aadmi ni straci ndeanti oxi dantes
enratonesdeescasal ongevi dad
sobreel porcentaj e de ratones"rpi dos" enl a prueba
de acti vi dadexpl oratori aespontneaenel
l aberi nto.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
2. 3 . Determi naci onesBi oqu mi cas
2. 3. 1 . Consumodeox geno
mi tocondri al
El ndi ce de control
respi ratori o ( I CR) es una
medi da di recta del
acopl ami ento
del a mi tocondri ayde l a
ef i ci enci a enl a generaci nde
ATPpor
l a
cadenadetransporte de
el ectrones mi tocondri al . La
determi naci ndel I CR
( que se def i ne como
el coci ente entre el estado 3, o acti vo,
de l a respi raci n
mi tocondri al y
el estado 4, o de reposo)
es ti l paracomprobar l a i ntegri dad
f unci onal de l as mi tocondri as
. El mtodo de determi naci n
del I CR se
estandari z en
mi tocondri as de ani mal es j venes
( 16 semanas) , obteni ndose
unval or
de I CR de 4, 160, 36 en
mi tocondri as hepti cas, mi entras
que en
mi tocondri as renal es f ue
de 5, 361, 38.
Latemperaturael egi dapara
medi r el consumodeox genof ue
25 C, ya
que en control es
previ os se observ unamayor
ef i caci a respi ratori a aesta
temperaturaquea37 C, yel
medi outi l i zadof ueel tampndeensayo
descri to
enl a secci n"Materi al y
Mtodos", supl ementadoconrotenona
parabl oquear
el compl ej o I
de
l a
cadenadetransporte deel ectrones
.
El
consumode ox geno
se i ni ci
tras l a adi ci n de succi nato
5 mMcomo sustrato de l a cadena
de
transporte de el ectrones
( estado4) , unavez estabi l i zado el regi stro
se aadi
ADP/Mg2+
0, 4 mM( estado3) .
Se determi n el consumo
deox genoenambos estados del arespi raci n
mi tocondri al ,
enmi tocondri as ai sl adas deh gadoyri n
deani mal es adul tos
yvi ej os.
*
Mi tocondri as hepti cas
Al
comparar l os val ores deI CRobteni dos
enmi tocondri as hepti cas de
ani mal es de 48 ( I CR=2, 160, 26)
y62 ( I CR=2, 560, 30) semanas de edad con
l os obteni dos aparti r
deani mal es de 16 semanas ( I CR =
4, 160, 36) , se puede
observar
que se produce un deteri oro de l a i ntegri dad
mi tocondri al
paral el amenteal envej eci mi ento cronol gi codel os ratones
.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Resul tados
Comose observa
enl aTabl a9, di chaal teraci nse paral i za unavez que
el ani mal al canza l a edad adul ta ya que no se observaron di f erenci as
si gni f i cati vas enel consumode02
entrel os ani mal es adul tos yvi ej os .
Al estudi ar el ef ecto de l os anti oxi dantes sobre el consumo de 02
mi tocondri al ( Tabl a 9) se observ
queel I CRde l as mi tocondri as de ani mal es
adul tos cuya di eta conten a
f ue si gni f i cati vamente superi or al I CR de l as
mi tocondri as
de ratones adul tos control , mi entras que TP en este grupo
careci deef ecto ( Fi gura47) .
En mi tocondri as
hepti cas procedentes de ani mal es vi ej os ( de 62
semanas) no se observaron di f erenci as enel consumode
02
enel estado 3y4
conrespecto al os ani mal es adul tos ( Tabl a 9) . En
este grupo de ani mal es que
consumi eron
pi enso conanti oxi dantes durante 5 meses, sl o TPmantuvol as
mi tocondri as hepti cas acopl adas ya
que el
val or
de I CRobteni do en l os
ani mal es de 62 semanas
de edad f ue semej ante al de l os ani mal es j venes
( Fi gura47) .
Tabl a 9. EFECTO
DE ANTI OXI DANTESSOBREEL CONSUMODEOX GENODE
MI TOCONDRI ASHEPTI CAS
.
Consumo de ox geno en l os estados 4y 3
de
l as
mi tocondri as hepti cas ai sl adas de
ani mal es adul tos yvi ej os, expresado como mol es
02"
mgprote na- 1 "
mL- i ( ( a) = p<0, 05
vs adul to
control ; ( b) = p<0, 05 vs vi ej ocontrol ) .
Adul tos ( 48semanas) Vi ej os ( 62
semanas)
Estado
4
Estado3 Estado4 Estado 3
Control 0, 68 0, 11 1, 46
0, 19 0, 500, 07 1, 28 0, 49
NAC 0, 65 0, 07 1, 72 0, 50 0, 38 0, 18 1, 14
0, 36
TP
0, 27 0, 04a 0, 64 0, 07a 0, 24 0, 07b 0, 96
0, 21
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
2
1, 5 - '
1I
0, 5
= '
0
- -
Cont rol
NAC
TP
Resul t ados
Fi gura 47. EFECTODEANTI OXI DANTES
SOBREELI CRDEMI TOCONDRI AS
HEPTI CAS.
Consumo de ox geno
de mi t ocondri as hept i cas de rat ones cont rol y de l os que
consumi eron ant i oxi dant es, expresado
como I CR. (j venes= 16 semanas, n= 8;
adul t os= 48 semanas, n= 8; vi ej os= 62 semanas
;
n= 12) ((* ) = p<
0, 05 vs j oven cont rol ;
(* * ) = p<
0, 05 vs adul t o cont rol ; (* * * ) = p
<
0, 05
vs vi ej o cont rol ) .
* Mi t ocondri as renal es
Al cont rari o que
en mi t ocondri as hept i cas, el envej eci mi ent o de l os
rat ones
no
l l ev
asoci ado unaal t eraci n del as mi t ocondri as renal es yaque
no
suf ri eron un desacopl ami ent o ent rel a cadena
de t ransport e el ect rni co y l a
f osf ori l aci n oxi dat i va
ya quel os val ores de I CRobt eni dos en mi t ocondri as
ani mal es adul t os (4, 190, 69) y vi ej os (4, 410, 46)
no
si gni f i cat i vament e
del val or obt eni do en ani mal es j venes
Tampoco se observ ni nguna di f erenci a si gni f i cat i va
en el
ox geno
y por t ant o en el val or de I CR obt eni do de l as
renal es ent re l os ani mal es de ambos grupos adul t os o vi ej os
renal es de
di f erenci aron
(5, 361, 38) .
consumo de
mi t ocondri as
t rat ados con NACo TP(Tabl a10) .
se
5
O
j ovenes
"
adul t os
"
vi ej os
4, 5-
T
4-
3, 5
3 I
T
2, 5-
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Resul tados
As pues, nuestros datos i ndi canquel as mi tocondri as renal es nosuf ren
al teraci ones de su capaci dad respi ratori a debi das al envej eci mi ento o al a
supl ementaci n de l a di etacon anti oxi dantes.
Tabl a 10. EFECTODENACYTP SOBREEL CONSUMODEOXGENOEN
MITOCONDRIASRENALES.
Consumode Oz de l as mi tocondri as renal es ai sl adas de ani mal es adul tos yvi ej os,
expresadacomo bemol es de Oz " mgprote na- I " mL- 1 e ICR. ( Adul tos=48 semanas,
n=8;
vi ej os=62
semanas,
n=12)
.
2. 3. 2. Acti vi dadci tocrorno c oxi dasa
Se
determi n l a acti vi dad espec f i ca de este compl ej o respi ratori o en
mi tocondri as hepti cas yrenal es ai sl adas deratones dedi f erentes
edades. Los
val ores obteni dos a parti r de ani mal es j venes, de 16 semanas, f ueron
0, 680, 3 M" mgprot- 1 " mi n- 1 en mi tocondri as hepti cas y0, 570, 15 M" mg
prot- 1 " mi n- 1 enmi tocondri as renal es ( Fi gura48y49) .
Con l a edad,
l a acti vi dad enzi mti ca del enzi ma ci tocromo c oxi dasa
mostr una di smi nuci n si gni f i cati va en ambos ti pos mi tocondri al es,
descendi endohastaval ores de0, 200, 05en el caso
de mi tocondri as hepti cas
de ani mal es
adul tos o de 0, 290, 08 en el caso de mi tocondri as renal es de
ani mal es del ami smaedad( Fi gura48y49) .
Adul tos( 48semanas) Vi ej os ( 62semanas)
Estado4 Estado3 ICR Estado4 Estado 3 ICR
Control
0, 98 0, 34 4, 12 0, 90 4, 19 0, 69 0, 84 0, 21 3, 47 0, 96 4, 41 0, 46
NAC 0, 75 0, 24 3, 15
0, 48
4, 20
0, 71 0, 96 0, 20 3, 66 0, 82 3, 82
0, 90
TP 0, 61 0, 11 2, 85 0, 50 4, 67 0, 59 0, 81 0, 18 3, 65 0, 69 4, 50 0, 70
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
* Mi tocondri as hepti cas
Con respecto a l as mi tocondri as hepti cas, no se observ un cambi o
estad sti camente
si gni f i cati vo
enl a acti vi dad del ci tocromo c oxi dasadesdel as
48a l as 62semanas de edadyal determi nar l a acti vi dad de este enzi ma en
mi tocondri as hepti cas de l os ratones que consumi eron anti oxi dantes, se
observ que tanto NACcomo TP
f ueron i nef i caces tanto en ani mal es adul tos
comovi ej os (Fi gura48) .
1-
0, 9
0, 8
0, 7
0, 6
0, 5 -
0, 4
0, 3
0,
0, 1
-
0
T
Control NAC TP
pJ venes
"
Adul tos
Vi ej os
Resul tados
Fi gura 48. ACTIVIDAD CITOCROMOC OXIDASA DE MITOCONDRIAS
HEPTICAS: EFECTODELOSANTIOXIDANTES
Acti vi dad de l a ci tocromo c oxi dasa (expresada como Acti vi dad espec f i ca en M"mg
prote na- ] *mi n-1) en mi tocondri as hepti cas control yde ani mal es que consum an
di eta supl ementada con anti oxi dantes. (J venes: 16 semana s; n=8; Adul tos=48
semanas; n=8; Vi ej os=62semanas; n=12, (*) p<0, 05 vs j oven control ) . )
* Mi tocondri as renal es
Con respecto a l as mi tocondri as renal es de l os ani mal es control , se
observ una
di smi nuci n si gni f i cati va
de l a
acti vi dad enzi mti ca entre l os
ani mal es adul tos yvi ej os, en l os que el val or de l a acti vi dad espec f i ca del
enzi ma
ci tocromo c oxi dasadescendi hastaval ores de 0, 15 M"mg prot - ' "mi n-1
enel grupode l os ani mal es vi ej os, de 62semanas de edad(Fi gura49) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Resul tados
Enl a mi smaf i gur a se puede obser var queel tr atami ento conNACno
f ue ef i caz ni en l os ani mal es adul tos, cuya acti vi dad enzi mti ca i ncl uso
di smi nuy si gni f i cati vamente con r especto al adul to contr ol , ni en l as
mi tocondr i as
de ani mal es vi ej os cuya di eta f ue supl ementada con NAC
dur ante5meses.
El tr atami ento conTP,
no f ue
ef i caz
tr as
2
meses de tr atami ento con
este anti oxi dante per o s l o f ue tr as 5 meses de tr atami ento, ya que l a
acti vi dad
enzi mati ca de ci tocr omo c oxi dasa al canz val or es si mi l ar es al os
obteni dos enani mal es adul tos contr ol de48semanas deedad, ysuper i or es a
l aacti vi dadobser vadaenani mal es vi ej os si nsupl ementodeTP enl a di eta.
ANTIOXIDANTES
Acti vi dad de l a ci tocr omo
c oxi dasa de mi tocondr i as r enal es (expr esada como
acti vi dadespec f i ca
en
M"
mg pr ote na- 1" mi n- 1. (J venes: 16 semanas, n=8; Adul tos:
48semanas, n=8; Vi ej os: 62
semanas,
n=12)
. (* ) p<0, 05f r entej ovencontr ol , (* * ) p<
0, 05 f r ente adul to contr ol , (#) p<0, 05 vi ej o f r ente
adul to,
(##)
p<0, 05 f r ente vi ej o
contr ol ) .
0, 8-
OJ venes
MAdul tos
0, 7- ,
Vi ej os
U
0,
0, 5
0, 4
0, 3
* *
U
Q
0, 2
0, 1
Contr ol NAC
TP
Fi gur a 49. ACTIVIDAD
CITOCROMO COXIDASA RENAL: EFECTO DE
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Resul tados
2. 3
. 3.
Acti vi dadci tratosi ntetasa
Se determi n l a acti vi dad
de este enzi ma marcador
de matri z
mi tocondri al , en mi tocondri as hepti cas
yrenal es ai sl adas de ratones de
di f erentes edades. Los
val ores obteni dos aparti r de ani mal es
j venes, de 16
semanas, f ueron
94, 23 19, 75 nmol es" mi n- 1" mg
prote na- 1 en mi tocondri as
hepti cas y307, 6760, 44 nmol es" mi n-
I
"
mgprote na- i en mi tocondri as
renal es
( Fi guras 50y51) .
Con
l a edad, esta acti vi dad enzi mti ca
mostr una di smi nuci n
si gni f i cati va en ambos ti pos
mi tocondri al es, descendi endo en el
caso de
mi tocondri as hepti cas hasta
val ores de 45, 916, 34 en
ani mal es adul tos o
55, 66
12, 09 en ani mal es vi ej os; y
en el caso de mi tocondri as renal es
hasta
val ores de 116, 9417, 29
en ani mal es adul tos o
148, 010, 05 en ani mal es
vi ej os ( Fi guras 50y51) .
* Mi tocondri as
hepti cas
Cuando
se proporci on pi enso con NAC
al os ratones de 9meses
de
edaddurante2 5meses, l a
acti vi dad enzi mti ca f uel i geramente
superi or a
l os val ores obteni dos en
l os grupos control deani mal es
adul tos yenvej eci dos.
Si n embargo
tras el consumo de TP, el ef ecto
protector sobre l a acti vi dad
enzi mti ca f ue muyevi dente,
manteni endodi chaacti vi dad enzi mti ca,
tanto
en l os ratones adul tos
como en l os vi ej os, semej antes
al os obteni dos en l os
ani mal es
j venes de 16 semanas de
edad( Fi gura50)
*
Mi tocondri as renal es
Cuando se determi n l a acti vi dad
de l a ci trato si ntetasa
de
mi tocondri as renal es de
ani mal es control no se observaron
di f erenci as
si gni f i cati vas entre
ani mal es adul tosyvi ej os.
El
ef ecto de NACen ani mal es de
48y62 semanas de edadno mostr
di f erenci as si gni f i cati vas con respecto
al os ani mal es de l a mi smaedad del
grupo control . Si n
embargo el tratami ento con TP
durante 5 meses si f ue
ef i caz previ ni endounadi smi nuci n
del a acti vi daddel enzi maci tratosi ntetasa
en l os ani mal es
de62semanas de edad, al canzando
val ores semej antes al a
acti vi dad enzi mti ca de mi tocondri as
de ratones de 16 semanas de edad
( Fi gura51) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Resul tados
r o
120 -
100
80 - '
60
40
20 -
0
450 -
400 -
350 -
300 -
250
200 -
150
100
50
r
pJvenes
/Adul tos
/Vi ej os
Contr ol NAC
TP
Fi gur a 50. EFECTODELOSANTIOXIDANTESSOBRE
LAACTIVIDADCITRATO
SINTETASAHEPTICA.
Acti vi dad del enzi ma ci tr ato si ntetasa
enmi tocondr i as hepti cas (expr esada como
[ Acti vi dad catal ti ca] =nmol es" mi n
- ' " mg
pr ote na- ' )
. (Jvenes: =16 semanas, n=8;
Adul tos=48 semanas, n=8; Vi ej os=62
semanas,
n=12) . (* )
p<0, 05vs j ovencontr ol ;
(* * ) p<0, 05vs adul to contr ol ; (* * * ) p<0, 05vs. vi ej o contr ol .
p
JvenesaAdul tos gVi e, - - - ~
Contr ol NAC
TP
Fi gur a51. EFECTO
DENACYTPSOBRECITRATOSINTETASA
RENAL.
Acti vi dad del enzi ma ci tr ato si ntetasa en
mi tocondr i as r enal es (expr esada como
[ Acti vi dad
catal ti ca] =nmol es" mi n
- ' " mg
pr ote na
- ' ) . (Jvenes: 16 semanas, n=8;
Adul tos=48
semanas, n=8; Vi ej os=62 semanas, n=12 ) . (* )
p<0, 05 f r ente j ovenes
contr ol ; (* * )
p<0, 05vs vi ej os contr ol )
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Resul tados
2. 3. 4. Determi naci n
deMal ondi al deh do
El obj eti vo pri nci pal deestadetermi naci n
f uecomprobar si l os rganos
de l os ratones vi ej os ( 62
semanas) mostrabanunamayor peroxi daci n
l i p di ca
ypor l o tanto suf r an unenvej eci mi ento
prematuro. Para el l o se anal i zaron,
comoi ndi cador deperoxi daci n
l i p di ca, l os ni vel es demal ondi al deh do( MDA),
medi antel a reacci n conci do ti obarbi tri co
( TBARS) enhomogeni zados de
h gado y corteza renal . En
paral el o se cuanti f i caron estos ni vel es
en l os
rganos de ratones adul tos control
prematuramente envej eci dos yen l os de
ratones de l a mi sma
edad cronol gi ca ( 24 2 semanas) que no hab an
envej eci do
prematuramente.
Los ni vel es de MDA en
ratones prematuramente envej eci dos ( PE)
( ratones "l entos" en el
test del l aberi nto) sonsi gni f i cati vamente superi ores
a
l os no prematuramente envej eci dos ( NPE; ratones
"rpi dos" en el test del
l aberi nto), yl os ni vel es en
l os adul tos PE sonsi mi l ares al os del os vi ej os NPE
( resul tados nomostrados) .
Lai ngesti n
deanti oxi dantes ti l i cos NACyTP ti ene unef ecto evi dente
enl os ani mal es PE, reduci endo si gni f i cati vamente l os ni vel es
de MDA, que
l l egan aser si mi l ares al os que
muestranl os NPE control ( Fi guras 52y53) .
Por ej empl o,
en l a cortezarenal de ani mal es PE se demostraron ni vel es
de
2, 03 0, 28 nmol es/mgprotei na, yen ani mal es PE supl ementados
conNAC
l os val ores f ueron de 1, 56 0, 34 nmol es/mg
protei na, val ores si mi l ares al os
del MDAdel os ani mal es
control NPE ( 1, 51 0, 14)
.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Resultados
0
m
2, 5
2
0, 5
0
Fi gura 52. EFECTOPROTECTO DE LOS ANTIOXIDANTES SOBRE
LA
PEROXIDACINLIP DICAHEPTICA.
Ni veles
deMDAen h gadoderatonesde62 semanas. Prematuramenteenvej eci dos
( PE) ynoprematuramente
envej eci dos( NPE) , ( *=p<0, 05vscontrol NPE; #p<0, 05vs
control PE( n=6) ) .
2, 50
2, 00
-
1, 50 -
1, 00
0, 50
0, 00
control NAC TP
control NAC
TP
E3 NPE
MPE
0NPE
"PE
Fi gura 53.
EFECTOPROTECTODE LOS ANTIOXIDANTES SOBRE LA
PEROXIDACIN
LIP DICARENAL.
Ni veles de MDA
en corteza renal de
envej eci dos( PE) y
noprematuramenteenvej eci dos( NPE) . ( *=p<0, 05vscontrol NPE;
#p<0, 05vscontrol PE. ( n=6) ) .
ratones de 62 semanas. Prematuramente
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Volver al ndice/Tornar a l'ndex
DISCUSIN
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Di scusi n
1.
Model odeenvej eci mi entocel ul ar i n vi tro
Los resul tados
obteni dos en l os model os i n vi tro descri tos en esta
memori a pueden i nteresar a of tal ml ogos ygerontl ogos,
ya que desde el
puntode vi sta de l a patol og a
cel ul ar estos model os ayudan a entender l os
mecani smos espec f i cos
i mpl i cados en l a senescenci a del as cl ul as del EPRy
de
otras cl ul as del SNCyespeci al mente el papel del estrs
oxi dati vo en el
procesodel a muertecel ul ar l i gada al
envej eci mi ento (Harman, 1956; Mi quel ,
1991; Cohenet al . , 1994) .
Ef ectodel a exposi ci n decl ul as
de
EPRb
al peroxi ni tri to.
Nuestros resul tados sugi eren
que el dao oxi dati vo i nduci do por el
peroxi ni tri to en l as cl ul as del EPRbmanteni das
en cul ti voes semej anteal as
al teraci ones queti enen l ugar
duranteel envej eci mi ento normal , i n vi vo, del as
neuronas del encf al o y
otros ti pos de cl ul as di f erenci adas
(Mi quel , 1998) .
As , el tratami ento con peroxi ni tri to causa
una di smi nuci n de l a vi abi l i dad
cel ul ar . Adems, el peroxi ni tri to
produce una depl eci n energti ca de estas
cl ul as, como
demuestra l a reducci n si gni f i cati va del conteni do
i ntracel ul ar
de ATP(como ocurre al envej ecer) y, por otra parte, el
dao
cel ul ar
se
mani f i esta en f orma
de apoptosi s cuando se exponen l as cl ul as a l as
concentraci ones ms baj as de peroxi ni tri to
usadas en este trabaj o, mi entras
quehayunai nducci n
ms i ntensadenecrosi s cel ul ar a l as concentraci ones
ms al tas.
Nuestros resul tados apoyan el concepto
dequeel enzi ma PARPpuede
tener un ef ecto i mportante
tanto en el proceso de apoptosi s como en el
de
necrosi s. Como se demuestra por l a di smi nuci n de NAD,
sustrato espec f i co
del enzi ma PARP(Szabo et a1. , 1996
; Messmer et a1. , 1996) . As el presente
estudi ocorrobora l a hi ptesi s de
queel peroxi ni tri to es ci totxi co f rente a l as
cl ul as
del
EPRb, provocando una acti vaci n del PARP que pri va
compl etamentea l a cl ul a de su
conteni do de ATPytermi na i nduci endo l a
necrosi s cel ul ar .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Di scusi n
Reacci ones patol gi cas al estr s oxi dati vo, semej antes al as que causa
el per oxi ni tr i to, pr obabl emente contr i buyen al a r uptur adel a bar r er ahemato-
r eti ni ana en l as enfer medades i nfl amator i as ydegener ati vas de l a r eti na. En
enfer medades
i nfl amator i as
i ntr aocul ar es (como l as uve ti s) l a i nfi l tr aci n de
cl ul as i nfl amator i as desdel os vasos cor oi deos puedeser l a fuente de EROde
acci n anl oga al a del per oxi ni tr i to, per o tambi n
se
ha obser vado
que l as
cl ul as r esi dentes de l os tej i dos i ntr aocul ar es, como l as del epi tel i o
pi gmentar i o de l a r eti na o l as cl ul as gl i al es de Ml l er , pueden ser una
fuente
i mpor tante de r adi cal es l i br es de ox geno
(Gour eau et al . , 1994a, 1 . 994b; De
Kozaket a1. , 1997) . As , en l as
enfer medades degener ati vas de l a r eti na, como
l a DMAE, l a for maci n der adi cal es l i br es yel daooxi dati vo de l as cl ul as del
EPRpodr anestar i mpl i cadas (Snodder l y,
1995; Del cour t et al . , 1999) .
Enr esumen, enl o quer espectaal a patol og aocul ar , l as EROde acci n
l esi va semej ante al a del per oxi ni tr i to estar an especi al mente
i mpl i cadas enl os
pr ocesos i nfl amator i os y
degener ati vas del oj o, comol a uve ti s, l a r eti nopat a
di abti ca y l a DMAE(Rao, 1990; Gour eauet
al . , 1995;
Beatty
et al . , 2000) . De
acuer do con
este
concepto,
se ha obser vado per oxi daci n l i p di ca i n vi vo
i nduci da
por EROen enfer medades r eti ni anas degener ati vas (Ander son et al . ,
1984) yenel daofotooxi dati vo del a r eti na (Kaganet al . , 1973) .
De acuer do con nuestr os pr esentes
r esul tados, el estr s oxi dati vo
podr a tener
un
papel
fundamental en l os mecani smos quedesencadenan l a
muer te de l as cl ul as del EPR, un tema al que hasta ahor a
se ha pr estado
poca atenci n. En condi ci ones
fi si ol gi cas no par ece di smi nui r apr eci a-
bl emente el nmer o decl ul as del EPRi n vi vo. Si n
embar go,
l a
muer te cel ul ar
s quepodr aocur r i r en condi ci ones
patol gi cas enl as quehayunaumentoen
l a
pr oducci n de r adi cal es l i br es y/ o un descenso de
l as defensas
anti oxi dantes. Este concepto est de acuer do con
estudi os epi demi ol gi cos
sobr e l a di etayotr os factor es de r i esgo
tanto ambi ental es como de esti l o de
vi da que sugi er en que
el estr s oxi dati vo contr i buye, al a DMAE, cuya
l esi n
i ni ci al sel ocal i za en el EPR(Ca et al . , 2000) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Di scusi n
Depl eci n de ATPen cl ul as expuestas a hi poxi a qu mi ca,
gl ucopeni a, i squemi aohi poxi aambi ental .
General mente se acepta que l a hi poxi a i nhi be l a producci n de
radi cal es l i bres, si n embargo,
segn Smi th ( 1986) l os radi cal es l i bres de
ox genopuedendesencadenar reacci ones patol gi cas enel encf al o tras breves
epi sodi os deanoxi aoi squemi a. Estosedebeaqueel repenti nodescenso
del a
tensi n de
02 que
acompaa
a l a i squemi a permi te que se produzcan
reacci ones de radi cal es l i bres que al teran l as membranas mi tocondri al es,
provocando cambi os queal teran
su f l ui dez, con l a resul tante prdi da de
rendi mi ento
bi oenergti co y f unci n cel ul ar. Segn Smi th ( 1986) ,
concentraci ones de
02
tanbaj as como10-100 p. p. m. , comose encuentran
en
graves epi sodi os dei squemi a,
bastanparai ni ci ar cadenas dedesorgani zaci n
peroxi dati val i gadas al as ERO. Por el l o, ennuestraopi ni n, tantol os
model os
i n vi tro deestrs oxi dati vo, como
l os model os de hi poxi a e i squemi a qu mi ca
causadas por
bl oqueo de l os mecani smos de l a cadena respi ratori a
mi tocondri al ydel a f osf ori l aci n oxi dati va
puedencontri bui r al a el uci daci n
de l a patogeni a
del proceso de envej eci mi ento cel ul ar y de muchas
enf ermedades degenerati vas asoci adas adi cho proceso.
De
acuerdo
con l o
anteri or, l os presentes experi mentos ennuestros model os
dehi poxi aqu mi ca,
gl ucopeni a, i squemi a e hi poxi a
ambi ental revel an un dao bi oqu mi co
expresadopor undescensoenl os ni vel es deATPenl as cl ul as del EPRb.
La
depl eci ndeATP
( queest di rectamenterel aci onadaconuni ncrementoenl a
l i beraci n al medi odecul ti vodeLDH) ti enel ugar enl as cl ul as quehan
si do
expuestas aci anuroyol i gomi ci na,
quei nducenunahi poxi a qu mi cayaqueel
ci anuro( CN) i nhi be
el enzi mami tocondri al ci tocromooxi dasayl a ol i gomi ci na
( OL) acta bl oqueando el enzi ma ATPsi ntetasa, al i gual
que
ocurre
en
condi ci ones dehi poxi aambi ental
. Los ni vel es deATPdescendi eron anms
cuandol as cl ul as del EPRb
f ueronexpuestas al as condi ci ones degl ucopeni a
i nduci das por el I AAyconl a consi gui ente i squemi a qu mi ca, l o quesugi ere
quel a gl ucl i si s no puedeser
esti mul ada para supl i r el descenso de ATP
( Pal meroet al . , 2000; 2001; Casti l l o et al . , 2002) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Di scusi n
Estas
observaci ones
concuerdan conestudi os de otros l aboratori os que
demuestranque l a exposi ci n de cl ul as del EPRba
estrs
oxi dati vo, hi poxi a
e
i squemi a qu mi ca i nduce cambi os
en el metabol i smo cel ul ar, debi do a una
al teraci n de l a cadena de transporte
mi tocondri al , como se observa al
envej ecer ( Mi quel , 1998;
2002), o de l a gl ucl i si s ( Rego et al . , 1995, 1996,
1997) . Si n
embargo,
segn
l as observaci ones de Rego et al . ( 1995), l os ni vel es
deATPno suf rencambi os si gni f i cati vos enl as cl ul as del areti na compl eta, l o
que sugi ere quese produce unaesti mul aci n
de l a gl ucl i si s para mantener
l os ni vel es normal es de ATP. La di f erenci a entre nuestros resul tados yl os de
Rego et al . ( 1995) se debe probabl emente aque
nosotros separamos el EPRb
de l a reti na neural , yl as cl ul as
de EPRb no pueden acumul ar gl ucgeno
debi do a que l a gl ucosa no es converti da en gl ucgeno en
este ti po cel ul ar
( Basi nger et al . , 1983) .
La condi ci n de gl ucopeni a no aumental a l i beraci n al medi o de LDH,
de acuerdo con el concepto de que l as
cl ul as del EPRb podr an uti l i zar
pi ruvato yl actato para l a
producci n de ATP si n i mpl i car l a v a de l a
gl ucl i si s
. Cuandose aade al medi oI AAyl a concentraci nde l actato
es al ta,
no se produceundescenso si gni f i cati vo enl os
ni vel es deATPenl as cl ul as de
reti na, segndemostraronl os estudi os
deWi nkl er ( 1981, 1983) .
Ef ectos benef i ci osos de l os anti oxi dantes
sobre l a concentraci n
deATP
El uso de supl ementos anti oxi dantes
para preveni r o f renar al gunas
degeneraci ones reti ni anas asoci adas
a l a edad encuentraj usti f i caci n en el
si gui ente comentari o de
Wi nkl er ( 1999) : "El compl ej o f otorreceptores/ EPR
est expuesto al a l uz sol ar, se encuentrarodeado
por ni vel es de
02
quel l egan
de l as arteri as y l as membranas
de este compl ej o conti enes al tas
concentraci ones
de
ci dos
grasos pol i i nsaturados" . En ef ecto,
todos el l os son
f actores deri esgo quevuel venal a reti na
muyvul nerabl e al daooxi dati vo. De
acuerdo con estos conceptos,
l a supl ementaci n del medi o de cul ti vo
con
anti oxi dantes est de acuerdo con l a si gui ente
opi ni n: "Probabl ementel os
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Di scusi n
si stemas de def ensa anti oxi dante en l a reti na soni mperf ectos,
permi ti endo
reacci ones
de autooxi daci nque ocurrenabaj avel oci dad al o l argo de l a vi da"
( Katz yRobi nson, 1986) .
Enel presente estudi o se ha escogi do l a vi tami na
Ccomo tratami ento
anti oxi dante debi do a que el
ascorbato es capaz de detener ef i cazmente l as
reacci ones de peroxi daci nl i p di ca que se producenencadena, a
causade l os
radi cal es
l i bres,
por l a
transf erenci ade unel ectrn ( Katz yRobi nson, 1986) .
Por otraparte se hanescogi do l os anti oxi dantes ti l i cos NACyTPpor el
papel f undamental de l os grupos ti l i cos
reduci dos para proteger contra l a
peroxi daci n
l i p di ca
y
l as mutaci ones uotros cambi os del ADNmi tocondri al
( Mi quel yFl emi ng, 1986; Mi quel , 1991, 1998) . Estudi os
anteri ores muestran
quetanto NACcomoTPretrasan
l os cambi os causados por el envej eci mi ento a
ni vel mi tocondri al y cel ul ar ( Mi guel
et al . , 1995) . Tambi n es rel evante en
rel aci nconel presente estudi o quel os ef ectos protectores de
l os tratami entos
con anti oxi dantes han si do
demostrados en l as cl ul as del si stema
i nmunol gi co. Enef ecto, l as i nvesti gaci ones del grupo
de De l a Fuente han
mostrado unapreservaci n de l as
f unci ones de l as cl ul as de este si stema en
experi mentos
enque se admi ni strNACy/oTPi n vi tro o i nvi vo ( Del
R oet al . ,
1998; De l a Fuente et al . , 1998a; Correa et al . ,
1999; Puerto et al . , 2002) . En
base a sus
resul tados, De l a Fuente ( 2002) concl uye que: " . . . puesto que el
si stemai nmunol gi co es uni ndi cador de sal udyunpredi ctor de
l ongevi dad,
l a protecci n de este si stemaproporci onada por l os
anti oxi dantes puedej ugar
unpapel i mportante paraconsegui r
unenvej eci mi ento consal ud" .
Nuestra presente i nvesti gaci n sugi ere que l os anti oxi dantes
tambi n
ti enen un ef ecto benef i ci oso sobre
el EPRb. As , l a supl ementaci n del medi o
convi tami na C( VC) causun aumento si gni f i cati vo enl os ni vel es
de
ATP
en
cl ul as de EPRb expuestas al as condi ci ones
de
hi poxi a
e i squemi a qu mi caas
como en l a hi poxi a ambi ental , pero no en condi ci ones de gl ucopeni a. Estos
resul tados concuerdan con l a f unci n f i si ol gi ca del ci do ascrbi co en el
metabol i smode estas cl ul as ( Del amereet al . , 1993; Hou
et
al . , 1998)
.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Di scusi n
Por ot ra part e, vari os est udi os real i zados enrat as muest ranque
cuando
se supl ement a
l a di et a con ci do ascrbi co aument a l a ext ensi n del
dao
ret i ni ano( Organi sci ak, 1990) .
Cuando el medi o
de
cul t i vo
de EPRb se supl ement con NAC, el ef ect o
benef i ci oso
sobre l a concent raci n de ATPf ue ms pronunci ado cuando l as
cl ul as f ueron
somet i das a condi ci ones de gl ucopeni a e hi poxi a qu mi ca.
Si n
embargo, en condi ci ones de i squemi a e
hi poxi a ambi ent al , el ef ect o de NAC
sobre l a
concent raci n de ATPf ue menor. El ef ect o benef i ci oso de NAC
sobre
est as cl ul as puede ser debi do a que
es un precursor del GSH, el cual
pot enci a vari os
mecani smos de def ensa f rent e a l a peroxi daci n excesi va
y
adems puede desempear una f unci n
cl ave en el mant eni mi ent o de l a
i nt egri dad f unci onal
de l as mi t ocondri as f rent e al est rs oxi dat i vo, con
l a
consi gui ent e
preservaci n de l a s nt esi s de ATP.
La ef i caci a de TP sobre l a concent raci n
de
ATP
f ue mayor en
condi ci ones de hi poxi a qu mi ca y
ambi ent al , ya que l a concent raci n de ATP
era si mi l ar
a l a observada en condi ci ones cont rol . Tambi n
t uvo ef ect o,
aunque menor en condi ci ones
de gl ucopeni a e i squemi a. El ef ect o benef i ci oso
de TP sobre
l as cl ul as somet i das a est os est reses met abl i cos se
debe
probabl ement e a que est e compuest oes unprecursor
del gl ut at i on, yt i ene un
papel esenci al
en l a conservaci n de l a i nt egri dad
f unci onal de l as
mi t ocondri as
( Haugaard et al . , 1969; Scadut o, 1990) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Di scusi n
Rel aci n del
estrs oxi dati vo yel envej eci mi ento con l a muerte
cel ul ar.
En l a presente
i nvesti gaci n se uti l i zaron di f erentes tcni cas para
comprobar
si con nuestros model os de estrs
oxi dati vo se pod a causar
apoptosi s. Con l a ti nci n de Hoechst,
que demuestra l a f ragmentaci n del
ADNnucl ear yencondi ci ones dehi poxi a
qu mi ca, gl ucopeni aei squemi a, no se
observ esta f ragmentaci n
. Esta aparente f al ta de ef ecto sobre
el proceso
apoptti co puededeberseaque,
aunquese hayaacti vadol aseal i zaci n para
que se desencadene
el proceso deapoptosi s, el ti empo que
estn l as cl ul as
someti das al estrs oxi dati vo es
demasi ado corto (15 mi nutos) para que
puedan aparecer l os
cambi os morf ol gi cos de l a muerteapoptti ca
(Anderson
et al . , 1999) . Mi entras que durante una
exposi ci n aprol ongada ahi poxi a
ambi ental s quese observ l a
muertepor apoptosi s comorevel al a ti nci nde
Hoechst .
Desde un punto de vi sta ms
general , rel aci onado con el
envej eci mi ento, l os model os i n vi tro
i nvesti gados en el presente estudi o pueden
tener i nters bi omdi co conrespecto al papel
de l as EROen l a patogeni adel
envej eci mi ento (Mi quel , 1998;
Pal mero et al . , 2000; Casti l l o et al . , 2002) . As ,
estos
model os
puedenayudar ael uci dar l os mecani smos que
rel aci onan al as
ERO, ya l os procesos de
muerte cel ul ar (necrosi s yapoptosi s), con l a
senescenci ade
l as cl ul as del EPRydeotros ti pos cel ul ares del
SNC. Aunque
l a prdi dadeneuronas yel i ncremento de l a
gl i a se hanobservado en vari as
regi ones encef l i cas en el
envej eci mi ento f i si ol gi co (Terryet al . , 1987) yen el
asoci ado con
enf ermedades neuro- degenerati vas (Whi tehouseet al . , 1982), no
se sabe cual es el pri nci pal mecani smo responsabl e
de
l a
muerte neuronal .
Al gunos autores proponen que
l a pri nci pal causade prdi daneuronal es l a
necrosi s uni da a al tos ni vel es de gl utamato (Dessi et al . ,
1993),
pero otros
como Zhang et al . (1995),
uti l i zando l a tcni ca del TUNELhan observado
cl ul as
apoptti cas
en l a zonaestri ada decerebros deratas adul tas. Adems,
estos autores sugi eren quel a
muerteprogramadadel as neuronas comi enza
en
un
estadi o
temprano del a vi daadul taysevai ncrementandoconl aedad.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Di scusi n
Aparentementel as
cl ul as del EPRnomuestranunmarcadodescenso
en nmero durante l a senescenci a pero pueden suf ri r apoptosi s
cuando el
envej eci mi ento se encuentra acompaado por
di f erentes ti pos de estrs
metabl i co como f al ta si mul tnea de
02 y
gl ucosa a causa de l a i squemi a
(Woodet al . , 1997) ohi peroxi a(Ca et al . , 2000) .
De acuerdo conl os datos anteri ores,
nuestros resul tados sugi eren que
l a i ncubaci n de l as cl ul as de EPRb, en condi ci ones de
hi poxi aambi ental y
en presenci a de
todos
l os
nutri entes, desencadena el proceso apoptti co
(Casti l l o et al . , 2002), l o que apoyael concepto de quel a muerte
apoptti ca
puedeser unmecani smode prdi dade cl ul as del EPR
durantel a f ase i ni ci al
del a DMAE(Ca et al . , 2000), yj usti f i cal a
i nvesti gaci nde l os tratami entos de
supl ementaci n
anti oxi dante.
El ef ecto protector contra l a muerte cel ul ar
de l os i nhi bi dores de l a
PARP, tambi nobservado enel presente
estudi o, apoyal a hi ptesi s de quel a
i nducci n
de l a PARP es un mecani smo i mpl i cado en
el dao cel ul ar
provocado por el estrs oxi dati vo. As
l o sugi ere el hechode que l a i nhi bi ci n
de l a PARPconsi gue mej orar l os ni vel es i ntracel ul ares
de ATP, ydi smi nuyeel
nmerodecl ul as apoptti cas encomparaci n
conel nmerode estas cl ul as
en muestras tratadas con
peroxi ni tri to si n el menci onado tratami ento
i nhi bi dor. La
parti ci paci n de l a PARPen el dao cel ul ar
provocado por el
estrs oxi dati vo podr areal i zarse medi anteuna
al teraci n del as mi tocondri as
quel l evar a al amuerte
cel ul ar (Bol aos et al . , 1996; Vi raget al . ,
1998) .
Comoya hemos comentado, l os trabaj os anteri ores
sugi eren que l os
anti oxi dantes ti l i cos ti enen un
ef ecto benef i ci oso sobre el proceso
de
envej eci mi ento, probabl emente a causa de su acci n
protectora sobre l as
membranas yel genoma
mi tocondri al f rente a l as EROproduci das en l a
cadena respi ratori a
(Mi quel 2002) . Adems, se acepta que
undescenso del
rendi mi entobi oenergti co(l i gadoaundao
mi tocondri al ) ti ene uni mportante
papel enel desencadenami entodel a
apoptosi s (Sastreet al . , 2000a) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Di scusi n
Por el l o es razonabl e
suponer que l a supl ementaci n de l a di eta con
anti oxi dantes ti l i cos podr a f renar tanto l a
desorgani zaci n mi tocondri al ( y
di smi nuci nde l a s ntesi s de ATP) comol amuerte cel ul ar apoptti ca que ti ene
l ugar al envej ecer.
Enbase a l o anteri or, hemos i nvesti gado
l os ef ectos de NACsobre l a
respuesta
de
l as
cl ul as del EPRb al oxi dante peroxi ni tri to, cuya
acci ntxi ca
ya ha si do demostrada por Behar-Cohen et al .
( 1996) . Segn nuestros
resul tados, NACprotege a l as cl ul as del EPRb
f rente al estrs oxi dati vo
i nduci dopor
peroxi ni tri to as comol a muerte cel ul ar programada
i nduci da por
ste.
Cuandol as cl ul as
se i ncubaronconl os anti oxi dantes y se
someti eron
ahi poxi a ambi ental , l os anti oxi dantes preservaron
l a muerte por apoptosi s del
mi smomodo que mantuvi eron
l os ni vel es de ATPy l a vi abi l i dad
cel ul ar, de
maneraque TP f ue el anti oxi dante msef i caz segui do
de NACy por l ti mo VC
que f ue el anti oxi dante que menos
protegi .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
2. Model odeenvej eci mi entoi nvi vo
Di scusi n
Longevi dad, pesocorporal yconsumodepi ensoen l os ratones
control yenl os tratados con
anti oxi dantes ti l i cos
El obj eti vo pri nci pal de esta parte del estudi o f ue
i nvesti gar si l os
anti oxi dantes ti l i cos NACy
TP ti enen al gn ef ecto benef i ci oso sobre el
envej eci mi ento de l os ratones. Segn nuestros resul tados,
l os grupos de
ratones cuyopi ensohab a si dosupl ementado
conestos anti oxi dantes ten an
unporcentaj e
desupervi venci a al as 62semanas mayor queel grupo
control ,
si endoel anti oxi danteTPel ms ef i caz parapreveni r contral a
mortal i dadal o
l argodetodo el estudi o. Por otra parte, l a
admi ni straci n deNACf uemuy
benef i ci osaparal a l ongevi dad
desdel as 44hastal as 58semanas deedad, a
j uzgar por el 100 %desupervi venci a de l os
ani mal es queconsum aneste
anti oxi dante
. Noobstante, desdel as 58 semanas hasta l a termi naci n del
estudi o, 62semanas, tuvol ugar unmarcado
descenso enl a supervi venci a
desdeel 90 %hasta el 60 %
del a pobl aci ni ni ci al , quees un
porcentaj e
si mi l ar al del asupervi venci adel grupocontrol .
Estos resul tados sugi erenque
el tratami entoprol ongadocon
NACpodr a tener unef ecto negati vo sobre el
equi l i bri ohomeostti coestrs
oxi dati vo/ def ensaanti oxi dante.
Otroparmetrodef ci l determi naci n, que
ti enei mportantes rel aci ones
conl os mecani smos del envej eci mi entonormal y
patol gi coyl al ongevi dad, es
el pesocorporal del ani mal . Si secomparanl os pesos
de
nuestros
ratones con
l a curvaponderal f aci l i tada por el centro
di stri bui dor del a cepa, seobserva
queel peso
del os ani mal es perteneci entes al os tres grupos ( control ,
NACy
TP) sehal l adentrodel rangoque
se
consi dera
normal paraestacepa. Adems,
comoen
estudi os anteri ores ( Economos yMi quel , 1980; DeJ uan, 1994), tuvo
l ugar unaumentoprogresi vodel pesoydel adesvi aci nestndar,
tantoenl os
ani mal es queconsum an el pi enso estndar
comoen l os queconsum an
pi ensosupl ementadoconNACoTP.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Di scusi n
Las desvi aci ones estndar de l as medi as de l os val ores de l os pesos de
l os dos grupos de ratones tratados conanti oxi dantes sonmenores
que l as del
grupo control , yde entre l os dos grupos que consum ananti oxi dantes,
l os
val ores de l os ratones TP son l os
que muestran una menor desvi aci n
estndar .
Esto sugi ere que TP puede tener un ef ecto benef i ci oso sobre el
metabol i smo,
l o que tambi nconcuerdaconel aspecto de l os ratones, pues l os
tratados con este anti oxi dante ti enen un aspecto semej ante a
l os ratones
j venes ms que l os ani mal es control ,
sobre todo en l o que respecta a l a
apari enci ade supel aj e yoj os.
La hi ptesi s de que NACyTP pueden proteger
contra l a prdi da de
homeostasi s
l i gada al envej eci mi ento tambi nest de acuerdo conl a
presente
observaci nde que, como en i nvesti gaci ones
anteri ores (Economos yMi quel ,
1980;
DeJ uan, 1994) , l a curvade di stri buci n de l os pesos corporal es
de l os
ratones control se "apl ana" conl a edad, mi entras quel as curvas
ponderal es de
l os grupos que consumenl os supl ementos
anti oxi dantes muestranunpi co en
el rango de pesos
"pti mos" (36, 0- 40, 9 g) , predi ctores de l ongevi dad,
que se
manti ene al o l argo de todo el per odo de estudi o
.
Resul ta evi dente, al comparar
l a Tabl a 5 con l a Fi gura 39, que el
anl i si s
de l a di stri buci nde l os pesos corporal es i ndi vi dual es
sumi ni strams
i nf ormaci nqueel si mpl e estudi o de l os
ef ectos de l a edadsobreel peso medi o
de l os
ratones. Enef ecto, di cho anl i si s detectal a apari ci n
de subgrupos de
ratones de peso no pti mo, es deci r
permi te i denti f i car l os ratones que
muestranuna
"edadbi ol gi ca" mayor que suedad cronol gi ca con
respecto a
l a
homeostasi s ponderal (Col l i er y Col eman, 1991) , con
l a consi gui ente
di smi nuci n de l as probabi l i dades de
al canzar una edad avanzada. Segn
nuestra
experi enci a, l a expresi n de l os datos en f orma grf i ca
(como en l a
Fi gura 39) f aci l i ta l a comparaci n retrospecti va
de
l as
di f erenci as del peso
entre el subgrupo de ani mal es
que sobrepasan l a edad de 62
semanas
(ratones
supervi vi entes; S) yl os que mueren
antes
de esa
edad (ratones no
supervi vi entes,
NS) . As se observa en estaf i gura queel pi co enl a
curvade
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Di scusi n
di st ri buci n de l os pesos de l os Scorresponde al val or de 33-34 g, mi ent ras
que casi coi nci den
l a curva de di st ri buci n de l os pesos de l os rat ones NS
(det ermi nados a l a edad de 32-38 semanas) yl a curva de l os
pesos de l os S
(det ermi nados a l as 40-46 semanas) . Osea que a l a
edad de 32-38 semanas
l os rat ones NSmost rabanen
l o que
respect a a
su homeost asi s ponderal una
edad bi ol gi ca aproxi madament e 8 semanas mayor que
l os que t en an
mayores expect at i vas de
l ongevi dad (S) .
Los dat os resumi dos enl a Tabl a 7 yFi gura 40permi t en
concl ui r que
l os rat ones quea l a edad
de 32-38semanas t i enenunpeso pt i mo o "j uveni l ",
comprendi do
ent re l os 30, 0 yl os 34, 9 g t i enenuna probabi l i dad de al canzar
l as 62 semanas de edad de aproxi madament e un
70 %, mi ent ras que l a
probabi l i dad de al canzar
est a edad es sl o de aproxi madament eun30%para
l os rat ones
depeso superi or a 34, 9 g a l as 32-38semanas.
La ut i l i dad del est udi o del peso para acl arar l os
mecani smos de acci n
"ant i -envej eci mi ent o" de
l os ant i oxi dant es ha si do conf i rmada por nuest ra
observaci n de que, como se muest ra en l a Tabl a
8 y Fi gura 41, l a
admi ni st raci n de NAC
y TP es especi al ment e ef i caz para aument ar l a
supervi venci a de l os rat ones de peso no pt i mo,
desde unos ni vel es de
aproxi madament e
el 30 %para l os cont rol hast a el 60 %y 70 %,
respect i vament e, para l os que consumen NACo
TP (l os porcent aj es
correspondi ent es a l os rat ones de peso pt i mo sonde
aproxi madament e70
para l os cont rol ,
70 %para l os NACy85%para l os TP) . El hecho de queel
ef ect o benef i ci oso de NACyTPsobre l a l ongevi dad sea mas evi dent e en
l os
rat ones de peso mayor
que 34, 9 g puededeberse, por l o menos enpart e, a
queest os aport adores de grupos t i ol reduci dos prot egen cont ra l a di sf unci n
del si st ema i nmunol gi co yconsi gui ent e aument o de
procesos neopl si cos (De
l a
Fuent e, 2002), quesonprobabl ement e ms f recuent es enl os ani mal es que
suf renl a def i ci ent e homeost asi s responsabl e del excesi vo
peso corporal
.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Di scusi n
Parmetros conductual es: pruebas de coordi naci n yvi gor y
de
acti vi dadexpl oratori a
espontnea.
Con respecto a l os ef ectos de l a edadydel tratami ento anti oxi dante
sobre l os parmetros conductual es, se observa queel
envej eci mi ento l l eva a
unaprdi da progresi va de l a capaci dad
de pasar con xi to l a prueba de l a
cuerdati rante, mi entras
quel os dos anti oxi dantes i nvesti gados di f i eren en su
acci n. As , l a supl ementaci n del adi eta
con
NAC
se acompaadeunef ecto
negati vosobrel acoordi naci n
neuromuscul ar. Por el contrari o, en el grupode
ratones queconsumi eronTP, el porcentaj e
deratones queresol v an el test con
xi to al f i nal i zar
el estudi o era mayor queen l os ani mal es control .
Si l os
resul tados seexpresancornoporcentaj ederendi mi ento
preservadoenrel aci n
con el rendi mi entoi ni ci al ( al quese asi gna
el
val or
de 100 %) , se observaun
marcadodescensodel rendi mi ento
del os ratones control ytratados conNAC,
mi entras
que
l a
prdi daderendi mi entoes menos evi dente
en
l os
ani mal es que
consumenTP.
En cuanto a
l a acti vi dadexpl oratori a espontneaval oradamedi ante
el
mtodo del l aberi nto en f orma de T, el
porcentaj e de ratones quepasan l a
pruebacon xi to en un
ti empo <_20 segundos ( ratones rpi dos) se manti ene
rel ati vamente constante en l os grupos
de
ratones
supervi vi entes del grupo
control , ei ncl usol os datos sugi eren
queel rendi mi entopodr amej orar
desde
l as 32-38 hastal as
40-46 semanas de edad. Noobstante, l os
cambi os que
ti enen
l ugar en este parmetroduranteel
tratami ento, desdel as 32-38al as
56-62 semanas, no son
estad sti camente si gni f i cati vos ni en el grupo
control
ni en l os
tratados con anti oxi dantes. Esta aparente
preservaci n del
rendi mi ento, quenormal mentedi smi nuye
al envej ecer ( DeJ uan, 1994) , qui zs
sedebeaqueenel
presenteestudi ol apruebadel l aberi nto
se
real i za
cadados
semanas ( en l ugar de cadames, como
en l os experi mentos deDeJ uan) . Por
el l o l os ef ectos
posi ti vos del entrenami ento podr an compensar
l os ef ectos
negati vos
del aedadencuantoal aacti vi dad
expl oratori aespontnea.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Di scusi n
SegndeDeJ uan( 1994) , l os ratones de 12 meses deedad quepasan
con xi to l a prueba de acti vi dad expl oratori a espontnea, al canzando l a
i ntersecci n de l os tres brazos del l aberi nto enunti empo 520 segundos
( ratones rpi dos) , ti enenunamayor l ongevi dadquel osratones
quenecesi tan
msti empo parareal i zar
l a
prueba( ratones
l entos) .
Conrespecto
a
l as causas de l a mortal i dad prematuradel os ratones
l entos, Del aFuenteet al . ( 1998a) handemostrado que, en
ratones OF1- Swi ss
vi ej os, l os ani mal es l entos ti enenpeores f unci ones
i nmunol gi cas ( y por l o
tanto menor protecci n congai nf ecci ones ytumores) que
l os rpi dos de l a
mi smaedad, otros
estudi os del grupo deDel aFuentehanconf i rmado quel os
ratones l entos ti enenunaf unci n f agoc ti ca menos acti va
que l os rpi dos
" . . . de acuerdo conl a estrecha rel aci n entre
el si stema i nmunol gi co y el
si stemanervi oso"
( Correaet al . , 1999) , yqueal a mi smaedad cronol gi ca, y
conrespecto anumerosos parmetros de i nmunocompetenci a,
" l os ratones
l entos son bi ol gi camente ms vi ej os que
l os rpi dos" ( Guayerbas et al . ,
2002a) . Este model o
de envej eci mi ento prematuro en ratn ( expresado en
df i ci ts conductual es enl as pruebas del l aberi nto yl a
cuerdati rante yen
di sf unci n i nmunol gi ca)
no sl o se hai denti f i cado enl a cepaOF- 1
Swi ss
pues, segnCorrea et al . ( 1999) , tambi nse puede
detectar en machos y
hembrasdel acepa
BALB/c.
SegnGi l ad y Gi l ad ( 1995) , l a menor l ongevi dad
de l os ratones que
muestran una def i ci ente conducta expl oratori a
( e i ncl uso l a denomi nada
f reezi ng behavi o~,
est l i gada aunadi sf unci n enl as reas del SNCque
determi nan l a respuesta a f actores estresantes. Como
una reacci n
i nadecuada a estos f actores es unade l as caracter sti cas
esenci al es del
envej eci mi ento,
ef ecti vamentesepuedeconsi derar al os ratones queno pasan
l a
pruebadel l aberi nto enunti empo <_20 segundos como prematuramente
envej eci dos en
l o querespectaaci ertas f unci ones de suSNCencomparaci n
conl osani mal esquepasanl apruebaconxi to ( ratones rpi dos) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Di scusi n
Nuestros datos confi rman que el porcentaj e de ratones rpi dos ti ene
una rel aci n posi ti va con l as expectati vas de supervi venci a de l os ani mal es
control , pero, como en el estudi o de De J uan ( 1994) , l a mayor expectati va de
l ongevi dad de l os ani mal es
tratados con NACno se refl ej a en una mayor
rapi dez de estos ratones en l a prueba del l aberi nto en comparaci n con l os
ani mal es control quereci benpi enso no supl ementado conanti oxi dante.
Parmetros bi oqu mi cos
El envej eci mi ento l l eva aun deteri oro de l as cl ul as somti cas yauna
i nvol uci n progresi va
de l as funci ones fi si ol gi cas, probabl emente l i gados a
una
al teraci n en l a funci n de l as mi tocondri as. La denomi nda "teor a
mi tocondri al " del envej eci mi ento est acredi tada por numerosos
trabaj os
( Mi quel , 1996; 1998) , donde
se demuestra que el proceso de envej eci mi ento
est
asoci ado aun acumul odedao en l a mi tocondri acomo consecuenci adel
estrs oxi dati vo. En este
senti do, en el presente trabaj o se handetermi nado
vari os parmetros bi oqu mi cos i ndi cadores del estado funci onal de l sa
mi tocondri as deh gadoyde ri n
deratones control ytratados conNACoTP.
Se acepta
que l a respi raci n mi tocondri al est estrechamente acopl ada
a l a fosfori l aci n oxi dati va en cl ul as i ntactas manteni das
en condi ci ones
fi si ol gi cas ( Ozawa et al . , 1987) , y
que el rango de val ores que defi ne l a
i ntegri dad
funci onal pti ma de l as mi tocondri as osci l a entre 3<I CR<15, como
demuestran trabaj os real i zados conmi tocondri as de h gado yri n ( Tager
et
al . , 1983; Hi rschet al . , 1989)
.
Por otro l ado, se hanobservado di smi nuci ones conl a
edaden
l a
efi caci a
de l a fosfori l aci n oxi dati va
en mi tocondri as de procedenci a si naptosomal
( Curti et al . ,
1990; Benzi et al . , 1992; Bowl i nget al . , 1993) ; de encfal o
entero,
no si naptosomal y corazn
( Guerri eri et al . , 1992; Takasawa et al . , 1993) ;
mscul o
( Trounce et al . , 1989; Cooper et al . , 1992; Mul l er- Hocker, 1992
;
ByrneyDennett, 1992) e h gado ( Trounce et al . ,
1989
;
Yenet al . , 1989) , tanto
deani mal es comodehumanos.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Di scusi n
Con
rel aci n a l os resul tados obteni dos en este estudi o se pudo
comprobar que l a
i ntegri dad f unci onal de mi tocondri as hepti cas yrenal es,
procedentes de ani mal es j venes ( 16 semanas de edad) , se mantuvoi ntacta,
antes de comenzar l a di eta supl ementada conanti oxi dantes, pues
l os val ores
obteni dos
de I CRse manti enendentrode este rango, conval ores de I CR4, 16
( mi tocondri as hepti cas) y5, 36 ( mi tocondri as renal es) .
Al comparar el I CR
de l as mi tocondri as hepti cas del grupocontrol , si n
supl emento anti oxi dante en l a di eta, se observ una di smi nuci n de este
ndi ce conl a edad, obteni endounval or de I CRenani mal es de 48semanas de
edad de 2, 16. Di choval or es i nf eri or al val or m ni moque
def i ne
l a
i ntegri dad
f unci onal de l as mi tocondri as, sugi ri endo que l a membrana de mi tocondri as
hepti cas suf re cambi os durante el envej eci mi ento
. Estos resul tados estnen
concordanci a conestudi os real i zados enmi tocondri as hepti cas de ratones de
l a cepa C57BL/6J que demuestran
tambi nque
conl a
edaddi smi nuye el I CR
adems
de
al terar
l a permeabi l i dad de l as membranas mi tocondri al es de
h gado( Godel l yCortopassi , 1998) .
Si nembargol as mi tocondri as renal es noparecen
suf ri r ni ngnproceso
degenerati voconl a edad,
manteni ndose el val or de I CRsi empre en el rango
de val ores
que def i nen l a i ntegri dad f unci onal de l as mi tocondri as
.
Estos
resul tados se aj ustan a
l a hi ptesi s pl anteada por Kwong ySohal ( 2000) ,
qui enes def i enden en sus estudi os que l a degeneraci n de l a i ntegri dad
mi tocondri al con l a edad puede deberse
a di f erenci as espec f i cas para cada
tej i doy/oal gradodi f erenci al de envej eci mi entoencada unode el l os.
Puesto que se ha sugeri do que l a i nacti vaci n oxi dati va
de enzi mas
cl ave enel metabol i smo
puede j ugar unpapel i mportante enel envej eci mi ento
cel ul ar,
hemos i nvesti gado l os ef ectos de dos compuestos ti l i cos sobre
enzi mas de l a f osf ori l aci n oxi dati va enmi tocondri as de h gadoyde ri n
de
ratones hembra OF1, que f ueron
al i mentados
con
pi ensosupl ementadocon
TPy
NACdurante 2y5 meses.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Di scusi n
Enl os ratones que consumi eron anti oxi dantes durante dos meses, el
estudi o de sus mi tocondri as hepti cas mostr que l a admi ni straci n de NAC
en el pi enso ej erc a una l i gera acci n protectora f rente a l os ef ectos del
envej eci mi ento sobre el I CRtras 2meses de tratami ento pero no durante 5
meses. Este hecho podr a expl i carse por unposi bl e ef ecto txi co deNACsobre
l a homeostasi s del ani mal
como
as
demuestra el i ncremento demortal i dad en
ratones tratados condurante ms de3meses. Tal vez este ef ecto seadebi do a
que aesa edadl as mi tocondri as de cl ul as hepti cas
sean
todav acapaces
de
adaptarse aunest mul o
txi co ymantener el coci ente respi ratori o aunque,
basndonos en l os reci entes hal l azgos sobre el dobl e ef ecto anti - ypro-
oxi dante de NAC( Sagri sta et al . , 2002), l os enzi mas pudi eran
estar
daados
por
l a acci n
deNAC
.
Por otro l ado, para poder observar unef ecto protector del
TP
f ueron
necesari os 5meses de
tratami ento, tras l os cual es, el I CRdel as mi tocondri as
hepti cas de ratones vi ej os f ue si mi l ar al de mi tocondri as de
ratones
control
de 16 semanas de edad. Estos datos
estnde acuerdo conestudi os real i zados
sobre
el
si stema i nmunol gi co, en l os que el tratami ento con TP es ms
ef ecti vo cuando l os ani mal es poseen df ci ts f unci onal es en su si stema,
( ani mal es envej eci dos) que
en
ani mal es
con un si stema i nmunocompetente
( ani mal es de corta
edad) ( Correaet al . , 1999) .
De acuerdo con trabaj os anteri ores real i zados con mi tocondri as
de
di f erentes rganos
de di f erentes especi es o cepas ( Vi tori ca et al . , 1981; Yenet
al . , 1989; Rooyackers et al . , 1996; Ti an
et
al .
1998;
Navarro
et al . 2002),
nuestros
resul tados tambi n muestran un marcado descenso con l a edad
tanto enl a acti vi dad espec f i cade ci tocromo coxi dasa
( compl ej o I V) como enl a
acti vi dad del enzi ma marcador de
l a matri z mi tocondri al , ci trato si ntetasa,
tanto en l as mi tocondri as
de h gado como enl as de ri n. Si n embargo, un
estudi o comparati vo de l a acti vi dad de todos l os compl ej os enzi mti cos de
l a
cadena de transporte de el ectrones
entre mi tocondri as de di f erentes rganos
deun
mi smo
ani mal , demostrquel os cambi os conl a edadenl a acti vi dad de
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Di scusi n
est os
si st emas
no si guen un "model o comn" en l os di f erent es t ej i dos
est udi ados ypodr a ser debi do al hecho de que l as mi t ocondri as en l os
di st i nt os t ej i dos son est ruct ural yf unci onal ment e
di f erent es (Kwong ySohal ,
2000) . Est os aut ores observaron que en h gado yri n, en concret o el
compl ej o I V yl a act i vi dad de ci t rat o si nt et asa, no suf ri an vari aci ones con
l a
edaden su act i vi dad en unperi odo de t i empo semej ant e, comprendi do ent re
l as 14 y56 semanas de edad. Est as di screpanci as con
nuest ros resul t ados
podr an basarse
t ant o en el mt odo de ai sl ami ent o de mi t ocondri as as como
en el mt odo ut i l i zado en l a det ermi naci n de l a act i vi daddel compl ej o I V, en
el cual l as mi t ocondri as f ueron somet i das a soni caci n,
mi ent ras que en
nuest ro est udi o
se
ut i l i zaron
i nt act as .
Con respect o a l a act i vi dad del compl ej o I V mi t ocondri al en rat ones
cuya di et a f ue supl ement ada con ant i oxi dant es, ni NACni
TP most raron un
ef ect o prot ect or en mi t ocondri as hept i cas. Si n embargo en mi t ocondri as
renal es, sl o l a admi ni st raci n prol ongada
(5 meses) de TP f ue ef i caz,
mant eni endo l a act i vi dad espec f i ca de ci t ocromo c oxi dasa semej ant e a
l a
obt eni da en el grupo cont rol de edad adul t a. Sobre l a act i vi dad
del enzi ma
ci t rat o si nt et asa al i gual que
en ci t ocromo c oxi dasa, NACno ej erci ef ect o
prot ect or ni en mi t ocondri as hept i cas ni en renal es, mi ent ras que TP
f ue
ef ect i va en mi t ocondri as hept i cas t ras 2 meses
de
t rat ami ent o
yen renal es
nuevament e f ueron necesari os
5 meses de t rat ami ent o para observar un
i ncrement o de
l a
act i vi dadenzi mt i ca.
NACes un reconoci do
ant i oxi dant e, barredor de radi cal es l i bres y
precursor del gl ut at i on (Aruoma et al . , 1989) ypuede act uar
por reacci n
di rect a de su grupo reduct or t i ol
yl os
radi cal es
l i bres de ox geno, prot egi endo
f rent e al dao oxi dat i vo i n vi t ro, como handemost rado ot ros aut ores (Hof f er et
al . , 1993; YanyGreene, 1998; Mart i nez et al . , 1997; Banacl ocha, 2001) y
l os
est udi os real i zados en est a
memori aen el model o i n vi t ro con cl ul as de EPR.
Si n embargo,
no aparecen ref erenci as en l a bi bl i ograf a consul t ada sobre l os
ef ect os de NACo TPi n vi vo osobre l os compl ej os de l af osf ori l aci n oxi dat i va.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Di scusi n
Los ni cos datos sonl os aportados
por DeJ uan ( 1994) qui en observa
un
ef ecto protector sobre l os dos compl ej os enzi mti cos. En este estudi o
observamos una f al ta de
ef ecto de NACsobre ci tocromo c oxi dasa, ci trato
si ntetasa y
sobreel I CRen mi tocondri as renal es, yunl i gero
ef ecto protector
de NACsol o en el I CRde l as mi tocondri as
hepti cas tras 2 meses de
tratami entopero notras 5 meses. Posi bl emente,
l a di f erenci a en cuanto al a
edad decomi enzo del
tratami ento, 8semanas mas tarde, o tal vez l a dosi s
uti l i zada ( 1%of rentea0, 2%uti l i zada en este estudi o) por De
J uanpodr an ser
l a causadel as di f erenci as
observadas.
Para
obtener ms i nf ormaci n sobrel os ef ectos desorgani zadores
del
estrs oxi dati vo, se determi n l a
concentraci n de perxi dos de l pi do en
homogeni zados
de h gado y de corteza renal , observndose
un aumento
si gni f i cati vo dedi chaconcentraci n
enl os control es vi ej os encomparaci ncon
l os control es
j venes. Por otra parte, unacomparaci n del os ni vel es de
MDA
en homogeni zados de
ratones control de242 semanas deedad
cronol gi ca
muestra quel os rganos de l os
ratones prematuramente envej eci dos ( PE) ,
segn l a prueba del
l aberi nto, han suf ri do una mayor peroxi daci n
l i p di ca
que l os de l os ani mal es no prematuramente
envej eci dos ( NPE) . Los
anti oxi dantes ti l i cos, NAC
y TP di smi nuyeron si gni f i cati vamente l a
peroxi daci n l i p di ca en homogeni zados
de
h gado
ycorteza renal deratones
vi ej os .
En
resumennuestros resul tados sugi eren quel a
admi ni straci n deTP
puedeproteger a l as mi tocondri as
hepti cas yrenal es f rente a l a al teraci n
peroxi dati va de sus
l pi dos estructural es que ocurre al
envej ecer,
contri buyendo
as apreservar l os compl ej os
respi ratori os yl a s ntesi s deATP
quees l a base del a f unci n
f i si ol gi ca de cada rgano yde l a l ongevi dad
f unci onal . Ms concretamente,
convi ene consi derar que el estrs
oxi dati vo
puede al terar l os l pi dos de l a
membrana mi tocondri al i nterna, con l a
consi gui enteprdi da
def unci ndel os compl ej os respi ratori os,
sobretododel I
yI V, quesonparti cul armentevul nerabl es
al ataquedel as especi es reacti vas
deox geno ( Zhang et al . ,
1990) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Di scusi n
Asi mi smo, el compl ej o I ( Fry y Green, 1981) , el compl ej o I V ( Whartony
Gri f f i th, 1965; Fry y Green, 1980; Vi k
et
al . ,
1981
;
Paradi es et
al . ,
1993) y
el
compl ej o V( Nohl y Kramer, 1980) pueden ser al terados por cambi os en el
entorno l i p di co de l a membrana i nterna mi tocondri al durante el
envej eci mi ento, especi al mente por el descenso en l os ni vel es de cardi ol i pi na
( Robi nson, 1993) .
Desde el punto de vi sta de l os probabl es mecani smos
del
envej eci mi ento,
nuestros datos apoyanel concepto de que, de acuerdoconl a
antes menci onadateor a mi tocondri al , l as l esi ones oxi dati vas del mtADNy l a
resul tante prdi dade capaci dad
bi oenergti cacel ul ar ti enenunpapel cl ave en
el
envej eci mi ento de dos rganos f undamental es para l a homeostasi s: el
h gadoyel ri n.
I mpl i caci ones cl ni cas,
desde el punto de vi sta de l a geri atr a
preventi va, de l os
presentes estudi os sobre l os anti oxi dantes ti l i cos.
Las ventaj as de l a supl ementaci n
de l a di eta con compuestos que
protegen a l as
mi tocondri as contra el estrs oxi dati vo y l a posi bl e apl i caci n
cl ni ca de estos compuestos,
que i ncl uyen a l os anti oxi dantes ti l i cos, han
si do obj eto
de unareci ente revi si n ( Mi quel , 2002) . Segnl as publ i caci ones
revi sadas, l as pri meras i nvesti gaci ones enani mal es tratados con
anti oxi dantes
ya demostraron que al gunos
de estos compuestos pod an aumentar l a
l ongevi dad
medi a de ratones y ratas, y que entre l os anti oxi dantes ms
ef i caces para retrasar el envej eci mi ento
se
encontraban
l os anti oxi dantes
ti l i cos. As , Mi quel y
Economos ( 1979) demostraronque l a supl ementaci nde
l a di eta
conti oprol i na sdi cao magnsi canosl o aumentabal a l ongevi dad de
drosf i l as y ratones si no que
tambi n f renaba su prdi da de capaci dad
reproductoray coordi naci nneuromuscul ar .
Apesar del ti empo
transcurri do desde estos estudi os enani mal es, aun
no se hani nvesti gado
l os ef ectos de l a i ngesta de estos anti oxi dantes para
detener el envej eci mi ento prematuroque suf rennumerosos humanosni para
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Di scusi n
proteger contra l os s ndromes degenerati vos l i gados a l a edad, en cuya
patognesi s parece estar i mpl i cado el estrs oxi dati vo,
comocataratas, DMAE,
arteri oscl erosi s ydi sf unci n
i nmunol gi ca (Harman, 1986; Mi quel , 1998) .
SegnYu (1999), unprobl ema rel aci onado conl a admi ni straci n de
anti oxi dantes es l a di f i ci l i nterpretaci n de l os
resul tados
a
causa de l a
homeostasi s "estrs oxi dati vo/ def ensa
anti oxi dante" de l os organi smos yel
consecuente reaj uste de l os ni vel es de l os di versos anti oxi dantes (f i si ol gi cos y
admi ni strados enl adi eta) .
Por otra parte, Rottkamp et al . (2000) comentan
que
l os
anti oxi dantes
general mente uti l i zados no pueden
causar aumentos consi stentes en l a
l ongevi dad mxi ma
de
l as
especi es y que esta rel ati va i nef i caci a de l os
tratami entos anti oxi dantes probabl emente se debe a
que se
di ri gen
a "di anas
i nadecuadas" .
Las
opi ni ones
antes resumi das j usti f i can el presente estudi o centrado
en mej orar l a protecci n anti oxi dante de l a
i mportante "di ana" del ataque
desorgani zador de l as EROquesonl as
mi tocondri as.
Laj usti f i caci npri nci pal del estudi o de l os
ef ectos de NACyTPsobre l a
l ongevi dad y l a sal ud en humanos
se basa en l a observaci n de que el
envej eci mi ento se acompaa de una progresi va oxi daci n del
gl utati n y de
otros compuestos ti l i cos en l os tej i dos
de l os vertebrados e i nvertebrados
(Mi quel y Weber, 1990) .
Adems, el uso de anti oxi dantes acti vos
sobre l as
mi tocondri as se j usti f i ca por el dato ms reci ente
de que el aumento del
coci ente GSSG/ GSHes mucho ms marcado
en l as mi tocondri as que en el
comparti mento extrami tocondri al
(Esteve et al . , 1999; Sastre et al . , 2000b) .
Por el l o ser a conveni ente real i zar ensayos cl ni cos
de NACyTP, puesto que
el evan l os ni vel es ti sul ares
de l os pri nci pal es ti ol es f i si ol gi cos
como el
gl utati on y ci ste na y por l o tanto aumentanl a
protecci n contra el estrs
oxi dati vo quesuf renl as
mi tocondri as (Dansette et al . , 1990) .
Tambi nj usti f i can una f utura apl i caci n
cl ni ca de l os anti oxi dantes
ti l i cos l os estudi os i nmuno-gerontol gi cos
de otros l aboratori os (De l a Fuente
yMi quel , 1994) .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Di scusi n
De entre l os
numerosos trabaj os del grupo de De l a
Fuente, ti ene
parti cul ar rel evanci a que el
tratami ento de ratones OF1 con NAC, TP o
mezcl as de ambos,
modul a numerosas f unci ones del si stema
i nmunol gi co,
con ef ectos benef i ci osos sobre l a i nmunocompetenci a. Es
muyi mportante
desde el punto de vi sta de una posi bl e
acci n preventi va del envej eci mi ento
patol gi co de l os
humanos, que en estudi os uti l i zando el model o de
envej eci mi ento
prematuro del ratn ( l i gado a sus al tos ni vel es
de ansi edad,
quei mpi denunarpi da expl oraci n del l aberi nto) ,
l a acci nesti mul antedel a
TP sobre l a f unci n
f agoc ti ca de l os macrf agos yprotectora de l a
normal
homeostasi s anti oxi dantes/ estrs
oxi dati vo ( con di smi nuci ndel os ni vel es de
superxi do
extracel ul ar) , f ue mas evi dente en l os ratones
prematuramente
envej eci dos queenl os quemostraban
unenvej eci mi ento normal ( Correa et al . ,
1999) .
Tambi n
ti ene gran i nters gerontol gi co l a observaci n
de que l a
canti dad deNACyTPnecesari a para aumentar l a
f agoci tosi s, l a acti vi dad de
l as cl ul as natural ki l l er
yespeci al mentel a respuesta prol i f erati va al mi tgeno
Concanaval i na
A( una f unci nquedi smi nuyemucho conl a edad) es mayor
en
l os ratones vi ej os que en l os adul tos ( De l a
Fuente, 2002) , l o queapoya el
concepto de que
l a homeostasi s se deteri ora al envej ecer . Esta
observaci n
est de acuerdo con
nuestros resul tados sobre l as al teraci ones del peso
corporal
en l os ratones queenvej ecen, yel mayor benef i ci o queobti enen del
tratami ento con l os anti oxi dantes ti l i cos l os ratones de
peso excesi vo ( y
menos ef i caz homeostasi s
ponderal ) en comparaci nconl os ani mal es depeso
normal .
Enresumen, nuestros datos sugi eren quel a acci n
anti oxi dante deTP
puedeproteger l as mi tocondri as
de
h gado yri n
de
l os
ratones, f renando el
envej eci mi ento
yconsi gui ente prdi da dehomeostasi s yf unci n ( i ncl uyendo l a
f unci n excretora y
detoxi f i cadora) de estos dos rganos. Adems, l os
resul tados del grupo de De l a Fuente muestran que NACyTP ti enen un
marcado ef ecto normal i zador de l a f unci n i nmunol gi ca,
sobre todo en l os
ani mal es de
homeostasi s def i ci entedebi do asu envej eci mi ento .
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Di scusi n
Esta acci n anti - envej eci mi ento de NACyTPsobre el h gado, ri n y
si stema i nmunol gi co ya demostrada
en el ratn de l aboratori o, aconsej a l a
real i zaci n
de estudi os cl ni cos. Su pri nci pal obj eti vo ser a i nvesti gar si estos
anti oxi dantes podr an, a travs de su acci n protectora sobre
ml ti pl es
mecani smos
homeostti cos quese al teranconl a edad, contri bui r apreveni r o
f renar vari as de l as enf ermedades degenerati vas, de patogeni al i gada al estrs
oxi dati vo, que di f i cul tan
o
i mpi den
l a consecuci n de unal arga vi da, con
sal udysati sf actori o rendi mi ento f unci onal
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Volver al ndice/Tornar a l'ndex
CONCLUSIONES
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Model ode
envej eci mi entocel ul ar i n vi tro
Concl usi ones
1 . Seobserv undaocel ul ar demostradopor l a i nhi bi ci n de l a s ntesi s de
ATPyel i ncrementodeLDHenel medi ocel ul ar al i ncubar l as cl ul as conPN,
conagentes qu mi cos i nductores
de
hi poxi a, gl ucopeni a ei squemi a
y
durante
una exposi ci n
prol ongada a baj as concentraci ones de ox geno. Por l o tanto
una di smi nuci n del ATPsugi ere que estos model os de estrs metabl i co
proporci onan
unnuevo
model o
de envej eci mi ento cel ul ar i n vi tro en cl ul as
di f erenci adas. Este model opuedeser uti l i zadocomocompl ementoal model o
ms ampl i amente uti l i zado hasta ahora que son f i brobl astos en cul ti vo
pri mari o.
2. El tratami ento
con
anti oxi dantes
del as cl ul as del EPRb, protegi f rente al
daocel ul ar i nduci do enl as condi ci ones anteri ores, si endo el ms ef i caz TP
segui dodeNACypor l ti mo
VC.
3. Del as anteri ores condi ci ones de i nducci n
de daopor estrs oxi dati vo,
sl o peroxi ni tri to
y una exposi ci n prol ongada a baj as concentraci ones de
ox geno
provocaronl a muertepor apoptosi s enestas cl ul as.
4. La apoptosi s
f ue
evi tada
cuandol as cl ul as setrataronprevi amenteconTP,
NACoVCantes deser expuestas aunahi poxi aprol ongada.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Concl usi ones
Model odeenvej eci mi ento i n vi vo
l . Lasupervi venci af uemayor enl os grupos
supl ementados conanti oxi dantes,
obteni endol a mayor supervi venci aenel gruposupl ementadoconTP.
2. Los ani mal es tratados conanti oxi dantes
manten ansupeso pti mo(36- 40
g) al o l argodel estudi o.
3. Los ratones no supervi vi entes,
pose anunadi stri buci n depeso semej ante
al a obteni daenani mal es demayor edad(48- 54 semanas) .
4. Lasupl ementaci ndel adi etaconTPmantuvounmej or aspectof i si co
y
de
coordi naci nneuromuscul ar .
5. Seproduj oundescenso del I CRmi tocondri al
tantoenh gadocomoenri n
dependi entede l a edad.
6. Laadmi ni straci ndeTPenel pi ensoej erci
unef ecto protector l os si stemas
enzi mti cos estudi ados
enmi tocondri as renal es.
7. Laadmi ni straci ndeanti oxi dantes
di smi nuyl a f ormaci nde perxi dos de
l pi dos tantoenmi tocondri as hepti cas comorenal es.
El anti oxi dante ti l i co TPes ms ef ecti vo en l a
prevenci n del dao cel ul ar
tantoi nvi tro comoi n vi vo
.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Volver al ndice/Tornar a l'ndex
BI BLI OGRAF A
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Bi bl i ogr af sa
- Adl er , R. , Cur ci o, C. , Hi cks, D. , Pr i ce, D. , Wong, F. . Cel l death i n age-
r el ated macul ar
degener ati on. Mol
.
Vi s. 5: 31- 39. 1999.
- Ai kens, J. , Di x, T. A. .
Per hydr oxyl r adi cal (HOO' ) i ni ti ated l i pi d per oxi dati on:
the r ol e of f atty aci dhydr oper oxi des . J. Bi ol .
Chem
. ,
266: 15091- 15098. 1991.
- Al der , R. , Jer dan, J. , Hewi tt,
A. T
. .
Responses of cul tur ed neur al r eti nal
cel l s
to
substr atum- bound l ami ni nand other extr acel l ul ar matr i x mol ecul es .
Dev. Bi ol . , 112: 100- 114. 1985.
- Ames, A. , Li , Y. Y. , Heher , E. C. , Ki mbl e,
C. R. . Ener gy metabol i smof r abbi t
r eti na as r el ated to f uncti on: hi gh cost of Na+tr anspor t. J. Neur osci . ,
12
: 840-
853. 1992.
- Ander son, K. M. , Seed, T. , Ou, D. , Har r i s, J. E. . Fr ee r adi cal s and
r eacti ve
oxygen speci es i n pr ogr ammed cel l death
. Med. Hypotheses, 52: 451- 463.
1999.
- Ander son, R. E. , Rapp, I . M. , Wi egand, R. D. . Li pi d
per oxi dati on and r eti nal
degener ati on. Cur r . Eye. Res. , 3: 223- 227.
1984.
- Ar mstr ong, R. S. , Sohal R. G. ,
Cutl er , R. G. , Sl ater , T. F. . Fr ee Radi cal s
Mol ecul ar . EnBi ol ogy, Agi ng andDi sease. Amstr ong
R. S. , Sohal R. G. , Cutl er
R. G. and Sl ater T. F(eds) .
Raven, NewYor k. 1984.
- Asahi , M. , Fuj i i , J. , Suzuki , K. , Seo, H
. G
. ,
Kuzuya, T. , Hor i , M. , et al . .
I nacti vati on of gl utathi oneper oxi dase
by ni tr i c oxi de. J. Bi ol . Chem. ,
270: 21035- 21039. 1995.
- Auer , R. N. . Exci totoxi c mechani sms, and
age r el ated suscepti bi l i ty to br ai n
damage i n i schemi a,
hypogl ycemi a and toxi c mussel poi soni ng.
Neur otoxi col ogy, 12: 541- 546. 1991.
- Aur oma, O. I . , Hal l i wel l , B. , Hoey, B. M. , Butl er ,
J. . The anti oxi dant acti on
of N- acetyl cystei ne
: i ts r eacti on wi th hydr ogen per oxi de, hydr oxyl r adi cal ,
super oxi de, andhypochl or ous aci d. Fr ee Radi c
.
Bi ol .
Med. , 6: 593- 597
.
1989.
- Autexi er ,
C
. , Gr ei der ,
C. W. . Tel omer ase and cancer : r evi si ti ng the tel omer e
hypothesi s . Tr ends Bi ochem. , 21: 387- 391. 1996.
- Baccanar i , D. P. . Coupl ed oxi dati on
of
NADPH
wi th thi ol s at neutr al pH. Ar ch.
Bi ochem. Bi ophys. , 191: 351- 357. 1978.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Bi bl i ograf a
-Banacl ocha, M. M. . Therapeuti c potenci al of N-acetyl cystei ne i n age-rel ated
mi tochondri al neurodegenerati ve di seases . Medi cal Hypotheses, 56: 472-477.
2001
.
-Basi nger, S. F. , Hof f man, R. T. . Bi ochemi stry of the pi gment epi thel i um.
En:
Bi ochemi stry of
the
Eye. Anderson
R. E
. (Ed
. ) . Ameri can Academy of
Ophthal mol ogy Manual s Program, SanFranci sco, pgs. 256-264. 1983
-Bast, A. , Haenen, G. R. M. M. . Cytochrome
Paso
and gl utathi one: What i s the
si gni f i cance of thei r i nterrel ati onshi p i n l i pi d peroxi dati on?. Trends Bi ochem
.
Sci . , 9: 510-513. 1984.
-Beatty, S. , Koh, H. , Phi l , M. , Henson, D. , Boul ton, M. . Therol e of oxi dati ve
stress i n the pathogenesi s of age-rel ated macul ar degenerati on
. Surv.
Ophthal mol . , 45: 115-134.
2000.
-Beckman, J . S. , Beckman, T. W. , Chen, J . , Marshal l , P. A. , et al . . Apparent
hydroxyl radi cal producti on by peroxyni tri te: i mpl i cati ons f or
endothel i al
i nj ury f romni tri c
oxi de
and
superoxi de. Proc. Natl . Acad. Sci . , 87: 1620-1624.
1990.
-Behar-Cohen,
F. F. , Heydol ph, S. , Faure, V. , et al . . Peroxyni tri te cytotoxi ci ty
on bovi ne reti nal pi gmented epi thel i al cel l s i n cul ture. Bi ochem. Bi ophys.
Res.
Commun. , 226: 842-849
.
1996.
-Bendi ch, A. , Ol son,
J . A
. .
Bi ol ogi cal acti ons of carotenoi ds. FASEBJ. , 3: 1927-
1931. 1989.
-Bennett, S. A. , Tenni swood, M. , Chen,
J . H
. ,
Davi dson, C. M. , et al . . Chroni c
cerebral
hypoperf usi on el i ci ts neuronal apoptosi s andbehavi oral
i mpai rment .
Neuroreport . , 9: 61-66. 1998.
-Benzi ,
G. , Pastori s, O. , Marzati co, F. , Vi l l a, R. F. , Dagani , F. , Curti
D. .
The
mi tochondri al el ectron transf er al terati on
as a f actor i nvol ved i n the brai n
agi ng. Neurobi ol .
Agi ng, 13: 361-368. 1992.
-Bergmeyer, H. U. , Brent, E. . En: Methods of
enzymati c anal ysi s. Bergmeyer,
H. U. (Eds) , NewYork, Academi c Press,
pgs. 574-579. 1974.
-Berman, E
. R. . Reti na. En: Bi ochemi stry of the eye. Perspecti ves i n vi si on
research,
Pl enumPress, pgs. 309-467. 1991.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Bi bl i ograf i a
- Bi anchi , A. , de Lange, T. . Ku
bi nds tel omeri c DNAi n vi tro. J. Bi ol . Chem. ,
274: 21223- 21227. 1999.
- Bi rnboi m,
H
. C. , Kanabus- Kami nska,
M. . The producti on of DNAstrand
breaks i n human l eukocytes by superoxi de may i nvol ve a metabol i c process.
Proc. Natl . Acad. Sci . , 82: 6820- 6824. 1985.
- Bl ackburn,
E. H
. .
Structure andf uncti on of tel omeres. Nature, 350: 569- 573.
1991.
- Bl ake, D. R. , Ral l en, R. E. , Lunec J. . Free radi cal s i n bi ol ogi cal
systems- a
revi ewori entatedto i nf l ammatory
processes. Br. Med. Bul l . , 45: 371- 385. 1987.
- Bl asco, M. A. , Lee, H. W. , Hande, P. , Samper, E. , et al . . Tel omereshorteni ng
andtumor f ormati on by
mousecel l s l acki ng tel omerase RNA. Cel l , 91: 25- 34.
1997.
- Boi x, J. , Ll echa, N. , Yuste, V. J. ,
Comel l a, J. X. . Characteri zati on of the cel l
death process i nducedby Staurospori ne i n HumanNeurobl astoma
Cel l
l i nes
.
Neuropharmacol ogy, 36: 811- 821. 1997.
- Bok, D. , Young,
R
. W. .
Phagoci tyc properti es of the reti nal pi gment
epi thel i um. I n: The
Reti nal Pi gment Epi thel i um. Zi nn, K. M. , Marmor, M. F. ,
( Eds) , Cambri dge, Massachusetts, Harvard Uni versi ty
Press, pgs. 148- 174.
1979.
- Bol aos, J. P. , Heal es, S. J. R. , Peuchen, S. , Barker,
M
. ,
Land, J. M. . Ni tri c
oxi de- medi ated mi tochondri al
damage: a potenti al neuroprotecti ve rol e f or
gl utathi one. FreeRadi c. Bi ol . Med. , 21: 995- 1001. 1996.
- Bol aos, J. P. , Peuchen, S. ,
Heal es, S. J. R. , Land, J. M. , Cl ark, J. B. . Ni tri c
oxi de- medi ated i nhi bi ti on of the mi tochondri al respi ratory chai n
i n cul tured
astrocytes. J. Neurochem. , 63: 910- 916. 1994.
- Boonman, Z. , I sacson, O. .
Apoptosi s i n neuronal devel opment and
transpl antati on: rol e of caspases andtrophi c f actors. Exp. Neurol . , 156: 1- 15.
1999.
- Boveri s,
A. , Oshi no, N
. , Chance,
B. . The cel l ul ar producti on of hydrogen
peroxi de. Bi ochem. J. , 128: 617- 630. 1972.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Bi bl i ograf t
- Bowl i ng, A. C. , Muti sya,
E. M
. , Wal ker, L
. C. ,
Pri ce, D. L. ,
Cork,
L. C- ,
Beal ,
M. F. . Age- dependent i mpai rment of mi tochondri al f uncti on i n pri mate brai n.
J. Neurochem. , 60
: 1964- 1967. 1993.
- Buettner, G. R. , Jurki ewi cz, B. A. . Catal yti c metal s, ascorbate and f ree
radi cal s: combi nati ons to avoi d. Radi at .
Res.
145: 532- 541.
1996.
- Buettner. G. R. . The pecki ng order of f ree radi cal s and anti oxi dants: l i pi d
peroxi dati on, al pha- tocopherol and
ascorbate
. Arch.
Bi ochem
.
Bi ophys. ,
300: 535- 543. 1993
.
- Burton, G. W. , I ngol d, K
. V. . R- carotene: Anunusual type of l i pi d anti oxi dant .
Sci ence, 224: 569- 573. 1984.
- Burton,
G. W
. ,
Joyce, A. , I ngol d, K. V. . Fi rst proof that vi tami nEi s themaj or
l i pi d- sol ubl e, chai n- breaki ng anti oxi dant i n human pl asma. Lancet,
2: 327-
328. 1982.
- Byrne, E. , Dennett, X. . Respi ratory chai n f ai l ure i n adul t muscl e
f i bres:
rel ati onshi p wi th agei ng
and possi bl e i mpl i cati ons f or the neuronal pool .
Mutat. Res. , 275: 125- 131. 1992.
- Cadenas, E. . Bi ochemi stry of oxygen toxi ci ty. Annu. Rev
.
Bi ochem. ,
51: 79-
110. 1989.
- Cadet, J. , Ravanat, J. L. , Buchko, G. W. , Yeo, H. C. , Ames, B. N. . Si ngl et
oxygen DNAdamage: chromatographi c and mass
spectrometri c anal ysi s of
damageproducts
. Methods enzymol . , 234: 79- 88. 1994.
- Cai , J. , Nel son, K. C. , Wu, M. , Sternberg, P. , Jones, D. P. .
Oxi dati vedamage
andprotecti on of the
RPE. Progr
.
Ret . EyeRes. , 19: 205- 221. 2000.
- Campi si , J. . Cancer, agi ng and cel l ul ar
senescence. I n Vi vo, 14: 183- 188.
2000.
- Campi si ,
J. .
The
bi ol ogy of repl i cati ve senescence. J. Cancer, 33: 703- 709
.
1997.
- Carl son, J. C. , Ri l ey, J
. C. M. .
A
consi derati on of some notabl eagi ng theori es.
Exp.
Gerontol
. ,
33: 127- 134. 1998.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Bi bl i ograf i a
- Cassari no, D. S . , Bennett, J . P. J r . . Aneval uati on of therol e of mi tochondri a
i n neurodegenerati ve di seases
:
mi tochondri al mutati ons and
oxi dati ve
pathol ogy, protecti ve nucl ear responses, andcel l death i n neurodegenerati on.
Brai nRes. Rev. , 29: 1- 25. 1999.
- Casti l l o, M. , Bel l ot, J . L. , Garc a- Cabanes, C. , Mi quel , J . , Orts, A. , Pal mero,
M. . Ef f ects of hypoxi a on reti nal pi gmented epi thel i umcel l s: Protecti on by
anti oxi dants. Ophthal mi c Res. Enprensa.
- Cerutti , P. A. . Prooxi dant states andtumor promoti on. Sci ence, 227: 375- 381.
1985.
- Chakraborti ,
T. , Das, S. , Mondal , M. , Roychoudhury, S. , et al . . Oxi dant,
mi tochondri a andcal ci um: anovervi ew. Cel l Si gnal , 11: 77- 85. 1999.
- Chance, B. , Si es, H. , Boveri s, A. . Hydroperoxi de
metabol i smi n mammal i an
organs. Physi ol . Rev. , 59: 527- 605. 1979.
- Choe, M. , J ackson, C. , Yu, B.
P. .
Li pi d
peroxi dati on contri butes to age-
rel atedmembraneri gi di ty. FreeRadi cal Bi ol . Med. , 18: 977- 984. 1995.
- Choi , J . H. , Yu, B. . PBrai n
synaptosomal agi ng f ree radi cal s andmembrane
f l ui di ty. Free
Rad.
Bi ol .
Med. , 18: 133- 139. 1995.
- Choi sy- Rossi , C. , Rei sdorf , P. , Yoni sh- Rouach, E. . Mechani sms of
p53-
i nduced apoptosi s: i n search
of genes whi ch are regul ated duri ng p53-
medi atedcel l death. Toxi col . Lett. , 102: 491- 496. 1998.
- Cl arke, P. G. , Posada, A. , Pri mi , M. P. , Castagne, V. . Neuronal deathi n
the
central nervous system
duri ng devel opment. Bi omed. Pharmacother. , 52: 356-
362. 1998.
- Cl ementi , E. , Brown, G. C. , Feel i sch, M. , Moncada, S. Persi stent i nhi bi ti on
of cel l respi rati on by ni tri c oxi de: cruci al rol e of S- ni trosyl ati on of
mi tochondri al compl ex
I andprotecti ve acti on of gl utathi one. Proc. Natl . Acad.
Sci . , 95: 7631- 7636. 1998.
- Cl utton, S. The i mportance of oxi dati ve stress i n apoptosi s. Bri ti sh Medi cal
Bul l eti n, 53: 662- 668.
1997.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Bi bl i ograf a
-Cohen,
G. M. , Sun, X. M. , Fearnhead, H. , MacFarl ane, M. , et al . . Format i on
of l arge mol ecul ar wei ght f ragment s of DNAi s a
key
commi t t ed
st ep of
apopt osi s
i nt hymocyt es. J. I mmunol . , 153: 507-516. 1994.
-Cohen, G. M. . Caspases
: t heexecut i oners of apopt osi s. Bi ochem. J. , 326: 1-16.
1997.
-Col l i er,
T. J . , Col eman, P. D. . Di vergence of bi ol ogi al andchronol ogi cal agi ng:
evi dencef romrodent st udi es. Neurobi ol . Agei ng, 12: 685-693. 1991.
-Col t on, C. A. , Pagan,
F. , Snel l , J. , Col t on, J. S. , et al . . Prot ect i on f rom
oxi dat i on enhances t he survi val of cul t ured mesencephal i c
neurons. Exp.
Neurol . , 132: 54-61. 1995.
-Cooper, J. M. , Mann, V. M. , Schapi ra, A. H. . Anal yses of
mi t ochondri al
respi rat ory chai nf unct i on
andmi t ochondri al DNAdel et i on i n humanskel et al
muscl e: ef f ect of agei ng. J. Neurol . Sci . , 113: 91-98. 1992
-Correa, R. , Bl anco, B. , Del Ri o, M. , Vi ct or, V. ,
Guayerbas, N. , Medi na, S. ,
De l a Fuent e,
M. . Ef f ect of a di et suppl ement ed wi t h t hi oprol i ne onmuri ne
macrophagef unct i oni namodel of premat ureagei ng. Bi oFact ors,
10: 195-200.
1999.
-Cot greave, I
. A
. ,
Mol deus, P. , Orreni us, S. . Host bi ochemi cal def ense
mechani smagai nst prooxi dant s. Annu. Rev. Pharmacol .
Toxi col . , 28: 189-212.
1988.
-Curt i , D. , Gi angare, M. C. , Redol f i , M
. E. , Fugacci a, I . , Benzi , G. . Age-
rel at ed
modi f i cat i ons of cyt ochrome C oxi dase act i vi t y i n di scret e brai n
regi ons.
Mech. Agei ngDev. , 55: 171-180. 1990.
-Dam, H. , Granados, G. H. .
Peroxi dat i on of body f at i n vi t ami n Edef i ci ency.
Act aPhysi ol .
Scand. , 10: 162-170. 1945
-Danset t e, P. M. , Sassi , A. , Deschamps,
C
. ,
et al . . Sul f ur cont ai ni ng
compounds as ant i oxi dant s.
En: Ant i oxi dant s t herapyandprevent i vemedi ci ne.
Emeri t , I . ,
Packer, L. , Aucl ai r, C. ( eds) , Pl enumPress. NewYorky
London, pgs.
209-215. 1990.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Bi bl i ograf i a
- Davi es, K. J. A. , Wi ese, A. G. , Sevani an, A. , Ki m, E. H. . Repai r systems i n
oxi dati ve
stress. En: Mol ecul ar Bi ol ogy of Agi ng. Fi nch, C. E. , Johnson, T. E.
( eds) , Wi l ey- Li ss, NewYork. 1990.
- Davi s, M. , Whi tel y, T. , Turnbul l , D. M. , Mendel ow, A. D. .
Sel ecti ve
i mpai rments
of mi tochondri al respi ratory chai n acti vi ty duri ng agi ng and
i schemi c brai ndamage. Acta Neurochi r. Suppl . , 70: 56- 58. 1997.
- De Duve,
Ch. . The peroxi some: a newcytopl asmi c organel l e. Proc. R. Soc.
LondonSer. B. , 173: 710- 720. 1969.
- De Fl ora, S. , I zzotti , A. , D' Agosti ni , F. ,
Cesarone, C. F
. .
Anti oxi dant acti vi ty
and other mechani sms of thi ol s i nvol ved i n chemopreventi onof mutati onand
cancer . Am. J. Med. , 91: 122S- 130S. 1991.
- De Juan, E
. J
.
. Marcadores de edad bi ol gi ca enel envej eci mi ento del ratn:
Apl i caci n
f armacol gi ca Tesi s Doctoral . Facul tad de Ci enci as . Uni versi dad de
Al i cante. 1994.
- De Kozak, Y. ,
Coti net, A. , Goureau, O. , Hi cks, D. , Thi l l aye, B. . Tumor
necrosi s f actor and ni tri c oxi de producti on by resi dent
reti nal gl i al cel l s f rom
rats presenti ng
heredi tary reti nal degenerati on. Ocul ar I mmunol . I nf l amm. ,
5: 85- 94. 1997.
- De l a Fuente, M. ,
Ferrandez, M. D. , Del Ri o, M. , et al . . Enhancement of
l eukocyte f uncti ons i n aged mi ce suppl emented wi th the
anti oxi dant
thi oprol i ne. Mech. Agei ngDev. , 104: 213- 225. 1998a.
- De l a Fuente, M. , Ferrndez,
M. D. , Muoz, F. , De Juan, E. , Mi quel , J. .
Sti mul ati on
by the anti oxi dant thi oprol i ne of the l ymphocyte f uncti ons of ol d
mi ce. Mech. Agei ng
Dev. , 68: 27- 36. 1993.
- De
l a
Fuente, M. , Mi ano, M. , Vi ctor, V. M. , Del Ri o, M. , Ferrndez M. D. ,
Di ez, A. , Mi quel ,
J. .
Rel ati on between expl oratory
acti vi ty and i mmune
f uncti on i n aged mi ce: a prel i mi nary study. Mech. Agei nDev. , 102: 263- 277.
1998b.
- De l a Fuente, M. ,
Mi quel , J
.
. Ef ecto protector de anti oxi dantes ti l i cos enl a
i nmunodepresi nasoci ada al envej eci mi ento. Rev. Esp. Geri atr. Gerontol . , 29:
246- 251. 1994.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Bi bl i ograf i a
-De l a Fuente, M. . Ef f ects of anti oxi dants oni mmune
system
agei ng.
Eur
.
J.
Cl i n. Nutr. ,
56: 55-58. 2002.
-DeVri es, N. , DeFl ora, S. . N-acetyl -l -cystei ne. J.
Cel l . Bi ochem. ,
17
: 270-277.
1993.
-Dean, P. , Key,
C
.
. Spati al def i ci ts onradi al mazeaf ter l arge tectal l esi ons i n
rats : possi bl e rol e of i mpai red scanni ng. Behav. Neural .
Bi ol
. ,
32: 170-190.
1981.
-Del
R o, M. , Ruedas, G. , Medi na, S. , Vi ctor, V. M. ,
De
l a Fuente,
M. .
I mprovement by several
anti oxi dants of macrophage f uncti on i n vi tro. Li f e
Sci ence,
63
: 871-881.
1998.
-Del amere, W. A. , Coca-Prados, M. ,
Aggarwal l , S. . Studi es on regul ati on of
the ascorbi c aci d
transporter i n a cel l deri ved f romrabbi t non-pi gmented
ci l i ary
epi thel i um. Bi ochi m. Bi ophys. Acta, 1149
: 102-108. 1993.
-Del court, C. , Cri stol , J. P. , Tessi er, F. , Leger,
C. L. , et al . . Age-rel ated
macul ar degenerati on
andanti oxi dant status i nthePOLAstudy. POLA
Study
Group. Pathol ogi es Ocul ai res Li ees a1' Age
. Arch
.
Ophthal mol . , 117: 1384-1390.
1999.
-Dempl e,
B
. , Li nn,
S. . 5, 6-saturated thymi nel esi ons i n
DNA: producti onby
ul travi ol et l i ght or hydrogenperoxi de
. Nucl ei c Aci ds Res. , 10: 3781-3789.
1982.
-Dempl e,
B. . Oxi dati ve DNAdamage: repai r and
i nduci bl e cel l ul ar responses.
Mutat. Envi ron. , A: 155-167. 1990.
-Dessi , F. ,
Charri aut-Maarl angue, C. , Khrestchati sky, M. , Ben-Ari , Y. .
Gl utamate-i nduced neuronal death i s not a
programmed cel l death i n
cerebel ar cul ture. J. Neurochem. , 60
: 253. 1993.
-Dorey, C. K. , Wu, G. ,
Ebenstei n, D. , Garsd, A. , Wei ter, J
. J. .
Cel l
l oss i nthe
agi ng reti na. Rel ati onshi p to
l i pof usci n accumul ati on and macul ar
degenerati on. I nvest.
Ophthal mol . Vi s. Sci . , 30: 1691-1699.
1989.
-Duboul oz, F. , Dumas, J. . Sur l e
metabol i smedes peroxi des l i pi di ques dans
l e peau au cours
des pr6cesses "i nf l ammatoi res" . Bul l .
Soc. Chem. Bi ol . ,
36: 983-987
. 1954.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Bi bl i ograf i a
-Duboul oz, F. , Dumas, J. . Sur l e rol e des peroxi des dans l a gense
desl esi ons
produi t par
l es radi ati ons. J. Radi ol . El ectrol . , 36: 343-370. 1955.
-Duke, R. C. . Apoptosi s i n cytotoxi c Tl ymphocytes and thei r
targets. Semi n.
I mmunol . , 4: 407-412. 1. 992.
-Economos, A. C. , Mi quel , J. .
Usef ul ness of stochasti c anal ysi s of body wei ght
as atool i nexperi mental agi ng research. Exp. Agei ngRes. , 6: 417-430
. 1980.
-Esteve, J . , Momp6, J. , Garci a de l a Asunci n,
J. . Oxi dati ve damage to
mi tochondri a) DNAand gl utathi one i n apoptosi s studi es i n vi vo and i n vi tro.
FASEBJ. , 13: 1055-1064. 1999.
-Evans, N
. M. , y Bi shop, K. S. . Onthe exi stence of a hi therto unrecogni zed
di etary f actor esenti al f or reproducti on. Sci ence,
56: 650-658. 1922.
-Fal t, L. Ani ntroducti on
to the vegetati ve f uncti ons. The l ens. En: Physi ol ogy
of the eye. Fal t, I . , Wei ssman, B. , ( Eds), Butterwoth, Hei nemann,
U. S. A
. , pgs
.
93-95. 1992.
-Ferrandi z, M. L. , Marti nez,
M. , De Juan, E. , Di ez, A. , et al . . I mpai rment of
mi tochondri a) oxi dati ve phosphoryl ati oni n the brai nof aged mi ce. Brai n
Res. ,
644: 335-338. 1994.
-Ferrari ,
G. , Yan, C. Y. I . , Greene, L. A. . N-acetyl cystei ne
( D-and-L- stereo-
i somers) prevent apoptoti c death
of
neuronal cel l s
. J. Neurosci . , 15: 2857-2866.
1995.
-Fi nch, C. E. . Longevi ty, senescence and the genome. Chi cago
: Uni versi ty of
Chi cago Press. 1990.
-Fi sher, D
. B. , Kauf man, S. . Tetrahydropteri n oxi dati onwi thout hydroxyl ati on
catal ysed by rat l i ver phenyl al ani ne hydroxyl ase. J. Bi ol . Chem. ,
248: 4300-
4304. 1973
.
-Fnan, S. , Cohen, G. . The oxi dant stress hypothesi s i n Parki nson' s di sease :
evi dence supporti ng i t . Ann.
Neurol
. ,
32
: 804-881
. 1992.
-Fraga, C. G. , Lei kovi tz, B. E. , Tappel , A. L. . Li pi d peroxi dati on measured as
thi obarbi turi c aci d-reacti ve substances i n ti suue sl i ces
: characteri zati on and
compari son
wi th homogenates andmi crosomes. Free Radi c. Bi ol . Med. , 4: 155-
161. 1988.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Bi bl i ograf a
-Freeman, B. A. ,
Grapo, J. D. . Bi ol ogy of di sease. Free radi cal s
and
t i ssue
i nj ury. Lab. I nvest . , 47: 412-426. 1982.
-Frei , B. , Engl and, L. , Ames, B. N. . Ascorbat ei s anout st andi ngant i oxi dant i n
human
bl oodpl asma. Proc. Nat l . Acad. Sci . , 86: 6377-6381. 1989.
-Frei , B. , St ocker, R. , Engl and, L. , Ames, B. N. . Ascorbat e: t hemost ef f ect i ve
ant i oxi dant i nhumanbl ood
pl asma. Adv.
Exp. Med
. Bi ol . , 264
: 155-163.
1990.
-Fri dovi ch, L. Superoxi dedi smut ases. Annu. Rev. Bi chem. , 44: 147-159. 1975.
-Fri dovi ch, L.
Superoxi de radi cal : An endogenous t oxi cant . Annu. Rev.
Pharmacol . Toxi col . , 23: 239-257. 1983.
-Fry, M. , Green, D. E. . Cardi ol i pi n requi rement f or el ect ron
t ransf er i n
compl ex I and I I I of t he
mi t ochondri al respi rat ory chai n. J. Bi ol . Chem. ,
256: 1874-1880. 1981.
-Fry, M. , Green, D. E. . I on-t ransport chai n of
cyt ochrome oxi dase: t he t wo
chai n-di rect coupl i ng
pri nci pl e of energy coupl i ng. Proc. Nat l . Acad. Sci . ,
77: 6391-6395. 1980.
-Garci a de l a
Asunci n, J. , Mi l l n, A, Pl a, R. , Bruseghi ni , L. , et al . .
Mi t ochondri al gl ut at hi one oxi dat i on correl at es wi t hage-associ at ed
oxi dat i ve
damaget o mi t ochondri a)
DNA. FASEBJ. , 20: 333-338. 1996.
-Gebi cki ,
S. , Gebi cki , J. M. . Format i on of peroxi des i n ami no aci ds and
prot ei ns exposedt o oxygenf ree radi cal s. Bi ochem.
J. , 289
: 743-749. 1
. 993.
-Gems, D. . Nemat odeagi ng:
Put t i ngmet abol i c t heori es t o t he t est . Curr. Bi ol ,
9: 614-616
. 1999.
-Gerschman, R. , Gi l bert , D. , Nye, S. W. ,
Dwyer, P. , Fenn,
W
. O. . Oxygen
poi soni ngand
X-i rradi at i on: amechani smi ncommon. Sci ence, 119: 623. 1954.
-Gerschman, R. .
Man' s dependence ont he eart hl y at mosphere. En: Proc
1St
symp
submari ne and space medi ci ne. Schaef f er, K. S. ( Ed) . MacMi l l an, New
York, pgs. 475. 1962.
-Gi l ad, G. H. , Gi l ad, V. H. .
St rai n, st ress, neurodegenerat i on and l ongevi t y.
Mech.
Agei ngDev. , 78: 75-83. 1995.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Bi bl i ograf i a
- Gl avi nd, J. , Hartmann, S. , Cl emmenses, J. , JensenK. E. , Dam,
H. .
Studi es
on the rol e of l i poperoxi des i n human pathol ogy. I I . The presence of
peroxi di zedl i pi ds i n thearteri ocl eroti c aorta
. Acta Pathol
.
Mi crob. Scand. , 30: 1-
6. 1952.
- Godel l , S. , Cortopassi , G. . Anal ysi s of oxygen consumpti on and
mi tochonddri al permeabi l i ty wi th age i n mi ce. Mech. Agei ng Dev. , 101
: 245-
256. 1998.
- Goureau,
O. , Bel l ot, J. , Thi l l aye, B. , Courtoi s, Y. , de Kozak, Y. . I ncreased
ni tri c oxi de producti on i n endotoxi n- i nduced uvei ti s.
Reducti on
of
uvei ti s by
ani nhi bi tor
of
ni tri c
oxi desynthase. J. I mmunol . , 154: 6518- 6523. 1995.
- Goureau, O. , Hi cks, D. , Courtoi s, Y. , de Kozak,
Y. . I nducti on and
regul ati on
of ni tri c oxi desynthase i n reti nal Ml l er gl i al cel l s. J. Neurochem. ,
63: 310- 317. 1994b.
- Goureau, O. , Hi cks, D. ,
Courtoi s, Y. . Humanreti nal pi gmented epi thel i al
cel l s
produce ni tri c oxi de i n response to cytoki nes. Bi ochem. Bi ophys. Res
.
Commun. , 198: 120- 126. 1994a.
- Gow, A. J . , Staml er, J. S. . Reacti on between m ni tri c
oxi deandhaemogl obi n
under physi ol ogi cal
condi ti ons. Nature, 391: 169- 173. 1998.
- Goyns, M. H. , y Lavery, W. L. .
Tel omerase and mammal i an agi ng: a cri ti cal
apprai sal .
Mech. Agei ngDev. , 114: 69- 77. 2000.
- Goyti sol o, F. , Samper, E. , Edmonson, S. ,
Tacci ol i , G. E. , et al . . Absenceof
ADN- Pkcs i n mi ce
resul ts i n anaphase bri dges and i n i ncreased tel omeri c
f usi ons wi th normal tel omere l ength and G- strand overhang
. Mol . Cel l Bi ol . ,
21 : 3642- 3651 .
2001.
- Goyti sol o,
F
. ,
Samper, E. , Marti n- Cabal l ero, J . , Fi nni n, P. , et al . . Short
tel omeres resul t i n organi smal hypersensi ti vi ty to i oni zi ng
radi ati on i n
mammal s. J. Exp. Med. , 192: 1625- 1636.
2000.
- Gray, L. H. , Scott, O. C. A. . Oxygentensi onandtheradi osensi ti vi ty
of
tumors
.
En: Oxygen i n the Ani mal Organi sm
. . Di ckens Fy. Nei l E. ( Eds) , Pergamon
Press, New
York, pgs. 537- 541 . 1964.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Bi bl i ograf i
-Greenwal d, R. W. ,
Moy, W.
W. .
Ef f ect of oxygen-deri ved f ree radi cal s on
hyal uroni c aci d. Art hri t i s Rheum. , 23: 455-463
.
1980.
-Guayerbas, N. , Cat al n,
M. , Vi ct or, V. M. , Mi quel , J. , De l a Fuent e, M. .
Rel at i on of behavi our and macrophage f unct i on t o l i f e
span
i n a
model
of
premat urei mmunosenescence. Behav.
Brai nRes. , 134: 41-48. 2002a.
-Guayerbas, N. , Puert o, M. , Ferrandez, M. D. , De La Puent e, M. . A di et
suppl ement ed wi t h t hi ol i c
ant i -oxi dant s i mproves l eucocyt e f unct i on i n t wo
st rai ns of premat urel y agei ng mi ce. Cl i n. Exp. Pharmacol . Physi ol . , 29: 1009-
1014. 2002b
-Guerri eri , F. , Capozza, G. , Kal ous,
M. , Papa, S. . Age-rel at ed changes of
mi t ochondri a) F0F1
ATPsynt hase. Ann. N. Y. Acad. Sci . , 671: 395-402. 1992
.
-Gut t eri dge, J. M. C. , Hal l i wel l , B. . The
measurement and mechani smof
l i pi d peroxi dat i on i n
bi ol ogi cal syst ems. Trends Bi ochem. Sci . , 15: 129-135.
1990.
-Gut t eri dge, J. M. C. .
Ant i oxi dant propert i es of t he prot ei n caerul opl asmi n,
al bumi nandt ransf erri ng: Ast udyof
t hei r act i vi t y i n serumandsynovi al f l ui d
f rompat i ent s wi t h rheumat oi d
art hri t i s Bi ochem. Bi ophys. Act a, 869: 119-127.
1986.
-Hal l i wel l , B. , Gut t eri dge, J. M. , Cross, C. E. . Free
radi cal s, ant i oxi dant s, and
humandi sease: wherearewenow?. J.
Lab. Cl i n. Med. , 119: 598-620. 1992.
-Hal l i wel l , B. ,
Gut t eri dge, J. M. C. . Li pi d peroxi dat i on: a radi cal chai n
react i on. I n: Free radi cal s i n bi ol ogy
and medi ci ne. Hal l i wel l , B. and
Gut t eri dge, J. M. C.
( eds), Oxf ordCl arendonPress, pgs. 188-276. 1989.
-Hal l i wel l , B. , Gut t eri dge, J. M. C. .
Oxygenradi cal s andt he nervous syst em.
Trends Neurosci . ,
6: 22-26. 1985.
-Hal l i wel l , B. , Gut t eri dge, J. M. C. . Oxygen
t oxi ci t y, oxygenradi cal s, t ransi t i on
met al s anddi sease. Bi ochem. J. , 219: 1-14
. 1984.
-Hal l i wel l , B. . Drug
ant i oxi dant s ef f ect s . A basi s f or drug sel ect i on.
Drugs,
42: 569-605. 1. 991.
-Hal l i wel l , B. . React i ve oxygen
speci es and t he cent ral nervous syst em. J.
Neurochem. , 59: 1609-1623
. 1992
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Bi bl i ogr af sa
- Hal l i wel l , B. . Vi tami nC: anti oxi dant or
pr o- oxi dant
i n
vi vo?
.
Fr ee
Rad. Res. ,
25: 439- 454. 1996.
- Hal l i wel l , B. . What ni tr ates tyr osi ne? I s
ni tr otyr osi ne
speci f i c as
a
bi omar ker
of per oxyni tr i te f or mati oni nvi vo?. FEBSLett. , 411: 157- 160. 1997
- Hande, M. , Samper , E. , Lansdor p, P. , Bl asco, M. A. . Tel omer e
l ength
dynami cs i n cul tur ed cel l s f r omnor mal andtel omer ase nul l mi ce. J. Cel l . Bi ol . ,
144: 589- 601. 1999.
- Har man,
D
.
. Agi ng: a
theor y
basedonf r ee
r adi cal
andr adi ati onchemi str y. J.
Ger ontol . , 11: 298- 300. 1956.
- Har man, D. . Fr ee r adi cal theor y of agi ng: r ol e of f r ee r adi cal s i n the
or i gi nati on
andevol uti on l i f e, agi ng, anddi sease pr ocesses . En: Fr ee r adi cal s,
agi ng and degener ati ve di seases. Wal f or d, R. L. , Johnson, J . , Har man,
D. ,
Mi quel ,
J. (Eds . ) Al anR. Li ss I nc. NewYor k. pgs. 3- 49. 1986.
- Har man, D. . Rol e of f r ee r adi cal s i n agi ng and di sease. Ann. N. Y.
AcadSci
. ,
673
: 126- 141. 1992.
- Har man, D. . Theagi ngpr ocess. Pr oc. Nat. Acad. Sci . , 78: 7124- 7128.
1981.
- Har man, D. . The bi ol ogi cal cl ock: The mi tochondr i a. J. Am. Ger i atr
. Soc. ,
20: 145- 147.
1972.
- Har r i s, E. D. . Regul ati on of anti oxi dant enzymes. FASEBJ. , 6
: 2675- 2683.
1992.
- Haugaar d, N. , Lee, N. H. , Kostr zeva, R. , Hor nR. S. , Haugaar dE. S. . The r ol e
of sul f hydr i l gr oups i n oxi dati ve phosphor yl ati onandi on
tr anspor t
by
r at l i ver
mi tochondr i a. Bi ochem. Bi ophys. Acta, 172: 198- 207. 1969.
- Hayf l i ck, L. . Thel i mi ted i n vi tr o l i f e span of humandi pl oi d
cel l str ai ns. Exp.
Cel l Res. , 37: 614- 636. 1965.
- Hi guchi , Y. , Li nn, S. . Pur i f i cati on of al l f or ms of HeLa cel l mi tochondr i al DNA
and assessment of damage to i t caused by hydr ogen per oxi de
tr eatment of
mi tochondr i al
or cel l s. J. Bi ol . Chem. , 270: 7950- 7956. 1995.
- Hi r sch, J. D. , Beyer , C. F. , Mal kowi tz, L. , Loul l i s, C. C. , Bl ume, A. J . .
Char acter i zati on of l i gand bi ndi ng to mi tochondr i a) benzodi azepi ne
r eceptor s.
Mol . Phar macol . 35: 164- 172. 1989.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Bi bl i ograf i a
- Hof f er, E. ,
Avi dor,
I . ,
Benj ami nov, O. , Shenker, L. , Tabak, A. , Tami r, A. ,
Merzbach. D. , Tai tel man, U. . N- acetyl cystei ne del ays the i nf i l trati on
of
i nf l ammatory
cel l s i nto the l ungs of paraquat- i ntoxi cated rats . Toxi col . Appl .
Pharmacol . , 120: 8- 12. 1993.
- Hof f man,
M
.
E. , Mel l o Fi l ho, A. C. , Meneghi ni , R. . Correl ati on between
cytotoxi cef f ect of H202 andthe yi el d of DNAstrandbreaki ncel l s of di f f erent
speci es. Bi ochi m. Bi ophys. Acta, 781
: 234- 238
.
1984.
- Hol z, F. G. , Schutt, F. ,
Kopi tz, J. , El dred, G. E. , et al . . I nhi bi ti on of
l ysosomal degradati ve f uncti ons i n RPEcel l s by a reti noi d
component of
l i pof ucsi n. I nvest . Ophthal mol . Vi s. Sci . ,
40: 737- 743. 1999.
- Horton, A. A. ,
Fai rhurst, S. . Li pi d peroxi dati onandmechani smof toxi ci ty
.
CRCCri t . Rev. Toxi col . , 18: 27- 79
.
1987.
- Horvi tz, H. R. . Geneti c control of programmed cel l death i n the
nematode
Caenorhabdi ti s el egans. Cancer Res.
59: 1701s- 1706s. 1999.
- Hou, Y. , Pi erce, W. M. ,
Del amere, W. A. . The i nf l uence of ascorbi c aci d
on
acti ve
sodi umtranspor i n cul tured rabbi t non- pi gmented
ci l i ary epi thel i um.
I nvest . Ophthal mol . Vi s. Sci . ,
39: 143- 150. 1998.
- Hsu, H. L. , Gi l l ey,
D
. ,
Backburn, E. , Chen, D. J . . Ku i s associ ated wi th the
tel omere i nmammal s. Proc. Natl . Acad. Sci . , 96
: 12454- 12458. 1999.
- I ml ay, J. A. , Li nn, S. .
DNAdamage and oxygen radi cal toxi ci ty. Sci ence,
240: 1302- 1309. 1988.
- I ngram, D. K. , y Jucker,
M. . Devel opi ng mouse model s of agi ng: a
consi derati on of strai n di f f erences i n age rel ated behavi oral and
neural
parameteers. Neurobi ol Agi ng,
20: 37- 145. 1999.
- Jacotot, E. ,
Costanti ni , P. , Laboureau, E. , et al . . Mi tochondri al
membrane
permeabi l i zati on duri ng the apoptoti c
process. Ann. N. Y. Acad. Sci . , 887: 18-
30. 1999.
- Jami eson, D. .
Oxygentoxi ci ty andreacti ve oxygenmetabol i tes
i nmammal s.
FreeRad. Bi ol . Med. , 7: 87- 108. 1989.
- Jazwi nski , S
. M. . The RASgenes: a homeostati c devi ce i n
Saccharomyces
cerevi si ae l ongevi ty. Neurobi o. I Agi ng,
20
: 471- 478
. 1999.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Bi bl i ograf i a
-J ohnson, T. E. . Geneti c i nf l uences onagi ng. Exp. Gerontol . , 32: 11-22. 1997.
-Kagan, V. , Shvedova, A: , Novi kov, K. , Kozl ov, Y. . Li ght-i nducedf ree radi cal
oxi dati on of membrane l i pi ds i n photoreceptors of f rog reti na. Bi ochem.
Bi ophys. Acta, 330: 76-79. 1973.
-Kappus, H. , Si es, H. . Toxi c drug ef f ects associ atedwi thoxygenmetabol i sm:
redoxcycl i ngandl i pi d
peroxi dati on. Experenti a, 37: 1233-1237. 1981.
-Katz, M. L. , Robi nson, W. G. . Nutri ti onal i nf l uences on autoxi dati on,
l i pof usci n accumul ati on, and agei ng. En: Free radi cal s, agei ng and
degenerati ve di seases.
J ohnson, J . E. , Wal f ord, R. , Harman, D. , Mi quel , J .
( Eds) , Al anR. Li ss, NewYork, pgs 221-259. 1986.
-Kennedy, C. J . , Rakoczy, P. E. , Robertson, T
. A. , Papadi mi tri ou, J . M. ,
Constabl e, I . J . . Ki neti c
studi es on phagocytosi s andl ysosomal di gesti on of
rodouter segments byhumanreti nal pi gment epi thel i al cel l s i nvi tro. Exp. Cel l
Res. , 210: 209-214. 1994.
-Ketterer, B. , Meyer, D. J . . Gl utathi one transf erases
: Apossi bl e rol e i n the
detoxi f i cati on and repai r
of DNAand l i pi d hydroperoxi des. Mutat . Res. ,
214: 33-40
. 1989.
-Ketterer, B. . Detoxi f i cati on reacti ons of
gl utathi one and gl utathi one
transf erases. Xenobi oti ca,
16: 957-973. 1986.
-Ki rkwood, T. B. . Bi ol ogi cal theori es of agi ng: an
overvi ew. Agi ng, 2: 144-146.
1998.
-Kl ei n, M. L. , J ori zzo,
P. A. , Watzke, R. C. . Growth f eatures of choroi dal
neovascul ar membranes i nage-rel atedmacul ar
degenerati on. Ophthal mol ogy,
96: 1416-1419. 1989
-Kobayashi , H. . The cel l ci cl e and tumor suppressor genes. Ri nsho Byori ,
44: 3-11.
1996
.
-Kono, Y. , Fri dovi ch, I . . Superoxi de radi cal i nhi bi ts catal ase. J. Bi ol . Chem. ,
257: 5751-5761. 1982.
-Kwong, L. K. , Sohal , R
. S
.
. Age-rel atedchanges i n acti vi ti es of mi tochondri al
el ectron transport compl exes i n vari ous ti ssues of the mouse. Arch. Bi ochem.
Bi ophys. , 373: 16-22. 2000.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Bi bl i ograf a
- Li pton, P. , Kal i l , R. . Neurotrophi c factors : thei r rol e i n the devel opment,
traumaanddi sease. Neural . Notes, 1
: 3- 7
.
1995.
- Li pton, S. , Choi , Y. B. , Pan, Z. H. , Lei , S. Z. , Chen, H. V. , et al . . Aredox-
based mechani smfor then neuroprotecti ve and
neurodestructi ve effects of
ni tri c oxi deandrel atedni troso- compounds
. Nature, 364: 626- 632. 1993.
- Li u, X. , Zhu, X. Z
. .
Rol es of p53, cMyc, Bcl 2, Bax and caspases i n serum
depri vati on- i nduced neuronal apoptosi s: apossi bl e neuroprotecti ve
mecha-
ni smof basi c fi brobl ast growthfactor.
Neuroreport. , 10: 3087- 3091 . 1999.
- Longo, V. D. . Mutati ons i n si gnal transducti on protei ns i ncrease
stress
resi stance and l ongevi ty i n yeast,
nematodes, frui t fl i es, and mammal i an
neuronal cel l s . Neurobi ol . Agi ng, 20: 479- 486. 1999.
- Lowry, O. H. , Rosebrough, N. J . , Farr,
A. L. , Randal l , R. J . . Protei n mea-
surement wi th the Fol i n phenol reagent . J . Bi ol . Chem. , 193: 265- 275. 1951 .
- Luo, Q. , Peyman, G. A. , Conway, M. , Wol teri ng, E. A. . Effect
of a
somatostati n anal og ( octreoti de acetate)
on the growth or Reti nal Pi gment
Epi thel i umcel l s i n cul ture. Curr. Eye. Res. , 15: 909- 913. 1996.
- Ly, D. H. , Lockhart, D. J . , Lerner, R. A. ,
Schul tz, P. G. . Mi toti c mi sregul ati on
andhumanagi ng
.
Sci ence,
287: 2486- 2492. 2000.
- Machl i n, L. J . , Bendi ch, A. . Free radi cal ti ssue
damage: Protecti ve rol e of
anti oxi dant
nutri ents
. FASEB
J . , 1 : 441- 445. 1987.
- Mal mstr6m, B. G. . Enzymol ogy of oxygen.
Annu. Rev. Bi ochem. , 51 : 21- 59.
1982
.
- Markl und,
S. L
. , Hol me, E. ,
Hel l ner, L. . Superoxi de di smutasa i n
extracel l ul ar fl ui ds. Cl i n. Chi m. Acts, 126: 41- 51 .
1982.
- Markl und, S. L. . Extracel l ul ar
superoxi de di smutasa i n human ti ssue and
humancel l l i nes. J . Cl i n. I nvest. , 74: 1398- 1403. 1984.
- Marks, N. , Berg, M. J . . Recent advances
on neuronal caspases i n
devel opment andneurodegenerati on. Neurochem. I nt. ,
35: 195- 220. 1999.
- Marti nez, D. E. . Mortal i ty patterns suggest
l ack of senescence i n hydra. Exp.
Gerontol . , 33: 217- 225. 1998.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Bi bl i ograf i a
-Marti nez, M. , Hernndez, A. I . , Marti nez, N. ,
Ferrndi z, M. L. . N-
acetyl cystei ne protecs agai nst age-rel ated i ncrease i n oxi di zed protei ns
i n
mousesynapti c mi tochondri a. Brai n
Res. , 762: 256-258. 1997.
-Marti nez-Cayuel a, M. .
Oxygen f ree radi cal s andhumandi sease. Bi ochi mi e,
77: 147-161. 1995.
-Marti nez-Cayuel a, M. . Toxi ci dad de xenobi 6ti cos medi ada por
radi cal es
l i bres deox geno. Ars.
Pharmaceuti ca, 39: 5-18. 1998.
-Mason, R. P. . Free-radi cal i ntermedi ates i n the metabol i smof toxi c
chemi cal s. En: Pryor W. A.
( ed) Free Radi cal s i n Bi ol ogy, ( 5) . Academi c Press,
NewYork, pgs. 161-173. 1982.
-Matsunaga, H. , Handa, J. T. , Aotaki -Keen,
A. , et
al .
. R-gal actosi dase
hi stochemi stry andtel omere
l oss i n senescent reti nal pi gment epi thel i al cel l s .
I nvest. Ophthal mol . Vi s. Sci . , 40: 197-202. 1999.
-May, J. , Cobb, C. E. , Mendi ratta, S. , Hi l l ,
K. E
. ,
Burk, R. F. . Reducti onof the
ascorbyl
f ree
radi cal
to ascorbate by thi oredoxi n reductase. J. Bi ol . Chem. ,
273: 23039-23045. 1998.
-Mayer, M. , Nobl e, M. .
N-acetyl -L-cystei ne i s a pl uri potent protector agai nst
cel l death andenhancer of trophi c f actor . medi ated
cel l survi val i nvi tro. Proc.
Natl . Acad. Sci . ,
91: 7496-7500. 1994.
-McAndrew, J. , Patel , R. P. , Jo, H. , Cornwel l , T. , Li ncol n, D. ,
et
al . .
The
i nterpl ay of ni tri c oxi de and
peroxyni tri tewi th si gnal transducti on pathways :
i mpl i cati ons
f or desease. Semi n. Peri natol . , 21: 351-366. 1997.
-McCord, J. M. , Fri dovi ch, L. Superoxi de and
superoxi de di smutases. En
Mi chel son, A. M. , McCord,
J. M. , Fri dovi ch, I . ( eds) . Academi c Press, NewYork.
1977.
-McCord, J. M. , Fri dovi ch, L.
Superoxi de di smutase: anenzymi c f uncti on f or
erythrocuprei n ( hemocuprei n) . J. Bi ol . Chem. , 244: 6049-6055. 1969.
-McCord, J. M. . Free radi cal s andi nf l ammati on:
Protecti onof synovi al f l ui d by
superoxi dedi smutase.
Sci ence, 185: 529-531. 1974.
-Medvedev, Z. . Anattempt at a rati onal cl assi f i cacati on of theori es of
agi ng.
Bi ol . Rev. , 65: 375-398.
1990.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Bi bl i ograf a
- Mei krantz, W. , Schl egel , R. .
Apoptosi s andthe cel l ci cl e. J. Cel l . Bi ochem. ,
58: 160- 174. 1995.
- Meri no, J. J . , Cordero- Campana, M. L. Mol ecul ar
bases of the programmed
cel l death process:
i mpl i cati ons of tumor suppressor protei n p53andother
protei ns i n
the control of cel l cycl e. Mechani sms of apoptoti c acti on. I nvest
.
Cl i n. , 39: 323- 358. 1998.
- Messmer, U. K. ,
Brne,
B. .
Ni tri c oxi de ( NO) i napoptoti c versus
necroti c RAW
264. 7 macrophage cel l death: the
rol e of NO- donor exposure, NAD+content,
and
p53accumul ati on. Arch. Bi ochem. Bi ophys. , 327: 1- 10. 1996.
- Mi chel i s, L. . Fundamental pri nci pl es
i n oxi do- reducti ons. Bi ol . Bul l . ,
92: 2939- 2947. 1949.
- Mi l l s, L. R. . The
sodi umcal ci umexchanger andgl utamate
i nducedcal ci um
l oads i n agedhi ppocampal neurons i nvi tro
. Ann. N. Y. Acad. Sci . , 15: 379- 390.
1996.
- Mi l l s, R. G. , Wei ss,
A. S. . Does progeri a provi de the
best model of accel erated
agei ng i n humans?. Gerontol ogy,
36: 84- 98. 1990.
- Mi not, C
. S. . The probl emof age, growth andage.
Pop. Sci . Monthl y, 71: 509-
527. 1907.
- Mi quel , J. , Bl asco,
M. . Asi mpl e techni que f or eval uati on
of vi tal i ty l oss i n
agi ng mi ce, by testi ng thei r muscul ar
coordi nati on andvi gor . Exp. Gerontol . ,
13: 389- 396. 1978.
- Mi quel , J. , Economos,
A. C. , Fl emi ng, J. , Jonhson, J. E. .
Mi tochondri al rol e
i n
cel l agi ng. Exp. Gerontol . , 15: 579- 591.
1980.
- Mi quel , J. , Economos,
A. C. , Johnson, J. E. . A
systems andl i si s-
thermodynami c vi ewof cel l ul ar andorgani smi c
agi ng. En: Agi ng andcel l ul ar
f uncti on. Johnson,
J. E
. ,
Jr
.
( Ed) , Pl enumPress, NewYork, pgs. 247- 280
. 1984.
- Mi quel ,
J. , Economos, A. C. . Favorabl e ef f ects
of the anti oxi dants sodi um
and magnesi umthi azol i di ne
carboxyl ate on the vi tal i ty and
l i f e span of
Drosophi l a
sp.
and
mi ce . Exp. GerontoL, 14
: 279- 285. 1979.
- Mi quel , J. , Ferrandi z, M. L. , De Juan, E. ,
Sevi l a, 1. , Marti nez, M. . N- acetyl -
cystei ne protects agai nst age- rel ated
decl i ne of oxi dati ve phosphoryl ati on
i n
l i ver mi tochondri a. Eur. J.
Pharmacol . , 292: 333- 335
. 1995
.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Bi bl i ograf a
- Mi quel , J. , Fl emi ng, J. E. . Theoreti cal and
experi mental support
f or
an
"oxygen radi cal - mi tochondri al i nj ury" hypothesi s of cel l agi ng. En: Free
radi cal s, agi ng and degenerati ve di seases.
Johnson, J . E. , Wal f ord, R. ,
Harman, D. , Mi quel , J. ( Eds) , Al anR.
Li ss, New
York, pgs
51- 74. 1986.
- Mi quel , J. , Weber, J. .
Agi ng andi ncreasedoxi dati onof the sul f ur pool . En:
Gl utathi one Metabol i smandPhysi ol ogi cal Functi ons.
Vi aJ. ( Ed. ) , .
CRC
Press,
BocaRaton, Fl ori da, pgs. 187- 192
. 1990
- Mi quel , J. .
Ani ntegratedtheory of agi ng as the resul t of mi tochondri al DNA
mutati oni ndi f f erenti atedcel l s. Arch. Gerontol . Geri atr. ,
12: 99- 117
. 1991.
- Mi quel ,
J. . An
update on the mi tochondri al - ADNmutati on hi p6tesi s of cel l
agi ng. Mutati onRes. , 275: 209- 216
. 1992.
- Mi quel ,
J
.
. Anupdate onthe oxygenstress- mi tochondri al mutati ontheory of
agi ng: geneti c andevol uti onary i mpl i cati ons. Exp. Gerontol . , 33: 113- 126. 1998.
- Mi quel , J. . Cananti oxi dant di et suppl ementati on
protect agai nst age- rel ated
mi tochondri al damage?.
Ann. N. Y. Acad. Sci . , 959: 508- 516. 2002.
- Mi quel , J. . Determi nati onof bi ol ogi cal age i n anti oxi dant treatedDrosophi l a
andmi ce. EmI nterventi on i n the Agi ng Process, Part B:
Basi c
Research and
Precl i ni cal Screeni ng
( Regel son, Ed. ) , Al an R. Li ss, NewYork, pgs. 317- 358.
1983.
- Mi quel , J. . Envej eci mi ento cel ul ar y mol ecul ar. En: Manual de geri atr a.
Sal gado, A. , Gui l l n, F. , ( eds) , Sal vat, Barcel ona, pgs
. 1- 18. 1990.
- Mi quel , J. . Envej eci mi ento
f i si ol 6gi co, cel ul ar ysubcel ul ar . En: Bi oqui mi ca y
bi ol ogi a mol ecul ar. Lel oi r, S. , Ochoa, S. , Oro, J . ,
Sol s,
A. ( Eds) , Sal vat,
barcel ona, pgs. 241- 248.
1986.
- Mi quel , J
.
. Nutri ti onandagei ng. Publ i c Heal thNutri ti on, 4: 1385- 1388. 2001.
- Mi quel , J. . Hi stori cal i ntroducti on to f ree radi cal andanti oxi dant bi omedi cal
research. En: Handbook
of f ree radi cal s and anti oxi dants i n bi omedi ci ne.
Mi quel , J. , Qui ntani l ha, A. , Weber, H. ( Eds) , Boca Raton Fl ori da: CRCPres,
pgs. 3- 13. 1989.
- Moncada, S. , Pal mer, R. M. , Hi ggs, E. A. . Ni tri c oxi de: physi ol ogy,
pathophysi ol ogyandpharmacol ogy. Pharmacol . Rev. ,
43: 109- 142.
1991.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Bi bl i ograf a
- Morel , F. , Mazet, F. , Tourai l l e, S. , Al zi ari , S. . Changes i n the respi ratory
chai n compl exes acti vi ti es
and
i n
the mi tochondri al DNA content duri ng
agei ngi n D. subobscura. Mech. Agei ngDev. , 84: 171- 181. 1995.
- Mul l er, H. J . . The remaki ng of chromosomes. Col l ect
. Net. Woods Hol e,
13: 181- 198.
1938.
- Mul l er- Hocker, J . . Mi tochondri aandagei ng. Brai nPathol . ,
2: 149- 158. 1992.
- Mul l i ken, R. S. . I nterpretati on of the atmospheri c absorpti on bands of
oxygen. Physi ol . Rev. , 32: 880- 887. 1929.
- Mund, M. L. , Rodri gues,
M. M. . Embryol ogy of the humanreti nal pi gment
epi thel i um
.
I n: The Reti nal Pi gment Epi thel i um. Zi nn, K. M. , Marmor, M. F. ,
( Eds) , Cambri dge, Massachusetts, HarvardUni versi ty
Press, pgs. 45- 52. 1979.
- Murphy, M. E. , Si es, H. .
Reversi bl e conversi nof ni troxyl ani onto ni tri c oxi de
by superoxi de di smutase. Proc. Natl . Acad. Sci . , 88: 10860- 10864. 1991.
- Nagata, S. . Fasl i gan- di nducedapoptosi s. Annu.
Rev. Genet. , 33: 29- 55. 1999.
- Nagy, Z. S. , Esi ri , M. M. . Apoptosi s- rel atedprotei n expressi on
i nthe hi ppo-
campusi nAl zhei mer' s
di sease. Neurobi ol . Agi ng, 18: 565- 571. 1997.
- Naqui , A. , Chance, B. , Cadenas, E. . Reacti ve oxygen i ntermedi ates i n
bi ochemi stry. Annu. Rev. Bi ochem, . 55: 137- 166. 1986.
- Navarro, A. , Snchez del
Pi no,
M
. J . , Gomez, C. , Peral ta, J . L. , Boveri s, A. .
Behavi oral dysfuncti on, brai n oxi dati ve stress, andi mpai red
mi tochondri al
el ectron transfer i n agi ngmi ce. Am. J . Physi ol .
Regul . I ntegr. Comp. Physi ol . ,
282: R985- R992. 2002.
- Ni el sen, H. . Coval ent bi ndi ng of peroxi di zed phosphol i pi d
to protei n. I I I .
Reacti on of i ndi vi dual
phosphol i pi ds wi th di fferent protei ns. Li pi ds, 16
: 215-
222. 1981.
- Ni ki , E. , Yamamoto, Y. ,
Komuro, E. , Sato, E. . Membrane damage due to
l i pi d
oxi dati on. Am. J . Cl i n. Nutr. , 53: 2015- 2055. 1991.
- Nohl , H. , Hegner, D. . Domi tochondri a
produce oxygenfree radi cal s i nvi vo?.
Eur. J . Bi ochem. ,
82: 563- 567. 1978.
- Nohl , H
. ,
Kramer, R. . Mol ecul ar basi s of age- dependent
changes i n the
acti vi ty of adeni nenucl eoti de transl ocase. Mech.
Agei ngDev. , 14: 137- 44. 1980.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Bi bl i ograf i a
- Ol ney, J. W. . Newmechani sms of exci t at ory t ransmi t t er neurot oxi ci t y. J.
Neural . Transm. , 43: 47- 51.
1994
.
- Organi sci ak, D. T. , Ji ang, Y. L. , Wang, H. M. , Bi cknel l , l . . The prot ect i ve
ef f ect of ascorbi c aci d i n ret i nal l i ght damage of rat s exposed t o i nt ermi t t ent
l i ght . I nvest . Opht hal mol . Vi s. Sci . , 31: 1195- 1202. 1990.
- Orgel , L. . Themai nt enance of prot ei n synt hesi s andi t s rel evance t o agei ng.
Proc. Nat. Acad. Sci . ,
49
: 517- 521.
1963.
- Osborne, N. N. , Cazevi ei l l e, C. , Pergande, G. , Wood, J. P. . I nduct i on of
apopt osi si ncul t ured humanret i nal pi gment epi t hel i al cel l s i s count eract ed by
f l upi rt i ne. I nvest . Opht hal mol . Vi s. Sci . ,
38
: 1390- 1400. 1997.
- Ozawa, T. , Tanaka, M. , Suzuki , H. , Ni shi ki mi , M. . St ruct ure andf unct i on
of mi t ochondri a: t hei r organi zat i on
anddi sorders. Brai nDev. , 9: 76- 81. 1987.
- Pal l ard6, EX. , Asensi , M. , Garci a de l a Asunci 6n, J. , et al . . Lat e onset
admi ni st rat i on of oral ant i oxi dant s
prevent s age- rel at ed l oss of mot or
coordi nat i on
andbrai n mi t ochondri al DNAdamage. FreeRadi c. Res. , 29: 617-
623. 1998.
- Pal mer, R. M. J . , Rees, D. D. , Asht on, D. S. , Moncada, S. .
L- argi ni ne i s t he
physi ol ogi cal precursor
f or t he f ormat i on of ni t ri c oxi de i n endot hel i um
dependent rel axat i on. Bi ochem. Bi ophys. Res. Commun. , 153
: 1251- 1256. 1988.
- Pal mero, M. ,
Bel l ot , J. L. , Cast i l l o, M. , Garci a- Cabanes, C. , Mi quel , J. ,
Ort s, A. . An i n vi t ro model of i schemi c- l i ke st ress i n ret i nal
pi gment ed
epi t hel i umcel l s:
Prot ect i ve ef f ect s of ant i oxi dant s. Mech. Agi ng Dev. , 114: 185-
190. 2000.
- Pal mero, M. , Bel l ot ,
J. L
. ,
Garci a- Cabanes, C. , Ort s. A. . Ani nvi t ro model of
oxi dat i ve st ress and hypoxi a i n ret i nal pi gment ed epi t hel i al cel l s" . J. Toxi col .
Cut. &Ocul ar Toxi col . , 20: 29- 38. 2001.
- Pal mero, M. . Caract eri zaci n
f unci onal del recept or di hi dropi ri di ni co
mi t ocondri al adrenomedul ar bovi no. Tesi s Doct oral . Facul t ad de Medi ci na.
Uni versi daddeAl i cant e. 1992.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Bi bl i ograf t
- Paradi es, G. , Ruggi ero, F. M. , Di noi , P. , Petrosi l l o, G. , Quagl i ari el l o, E. .
Decreased cytochrome
oxi dase acti vi ty andchanges i n phosphol i pi ds i n heart
mi tochondri a f rom hypothyroi d rats. Arch. Bi ochem. Bi ophys. ,
307
: 91-
95. 1993.
- Paschen, W. , Douthei l , J. . Di sturbance of endopl asmi c reti cul umf uncti ons:
a key mechani smunderl yi ng cel l
damage?. Acta Neurochi r. Suppl . , 73: 1- 5.
1999.
- Prez- Tur, J. . La genti ca y l a enf ermedad
de Al zhei mer. Rev. NeuroL,
30
: 161- 169
. 2000
.
- Porter,
R. S. . Therapeuti cs bi bl i ography f or cl i ni cal pharmaci sts:
respi ratory
di sease. Am. J. Hosp. Pharm. ,
37: 862- 864. 1980.
- Proctor,
P. H. , Reynol ds, E. S. . Free radi cal s and di sease i n man. Physi ol .
Chem. Phys. , 16: 175- 195. 1984.
- Pryor, W. . . Oxy- radi cal s
andrel ated speci es: thei r f ormati on, l i f e- ti mes, and
reacti ons. Annu. Rev. Physi ol . , 48: 657- 667.
1986.
- Pryor, W.
A. , Church, D. P. . Al dehi des, hydrogen peroxi de,
and organi c
radi cal as medi ators of oxygen toxi ci ty. FreeRad. Bi ol
. Chem. , 11 : 41- 46. 1991 .
- Pryor, W. . Free radi cal bi ol ogy, xenobi oti cs,
cancer andagi ng. Ann. N. Y. Acad.
Sci . , 393: 1- 16
. 1982.
- Pryor, W. . Free radi cal s i n bi ol ogy. I n: Free radi cal s
i n bi ol ogy. Vol 1 y 3.
Academi c
press,
New
York. 1976.
- Pryor, W. A. , Stone, K. . Oxi dant i n ci garette smoke
: radi cal s, hydrogen
peroxi de, peroxyni trate
andperoxyni tri te. Ann. N. Y. Acad. Sci . , 12- 27.
1993.
- Puerto, M. , Guayerbas, N. , V ctor, V. M. ,
De l a Fuente, M. . Ef f ect of N-
acetyl cystei ne
on macrophage andl ymphocyte f uncti ons i n amouse model
of
prematureagei ng. Pharmacol . Bi ochem. Behavi or, 73: 797- 804.
2002.
- Ramachandra, S. , Studzi nski , G. P. . Morphol ogi cal
and bi ochemi cal cri teri a
of apoptosi s. I n: Cel l
GrowthandApoptosi s. Apracti cal approach. Studzi nski ,
G. P
.
( Eds) , I RLPress, pgs. 119- 142. 1995.
- Randerath, K. , ReddyR. , Danna, T.
P. , Watson, W. P. , et al . . Formati onof
ri bonucl eoti des i n DNA
modi f i ed by oxi dati ve damage i n vi tro and
i n vi vo.
Characteri zati onby32Ppostl abel i ng.
Mutat. Res. , 275: 355- 366. 1992.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Bi bl i ograf i a
- Rao, N. A. . Rol e of oxygen f ree radi cal s i n ret i nal damage associ at ed wi t h
experi ment al uvei t i s. Trans. Am. Opht hal mol .
Soc. , 88: 797- 850. 1990.
- Rapp, L. M. , Anderson, R. E. . Vi sual pi gment dynami cs and t he vi t ami n A
cycl e. I n: Bi ochemi st ry of t heEye, Ameri canAcademy
of Opht hal mol ogy, pgs.
189- 195. 1983.
- Rat an, R. R. , Murphy, T. H. , Baraban, J. M. .
Macromol ecul ar shynt hesi s
i nhi bi t ors
prevent oxi dat i ve st ress- i nduced apopt osi s i n embryoni c cort i cal
neurons by shunt i ng cyst ei ne f romprot ei n synt hesi s t o gl ut at hi one. J.
Neurosci . , 17: 4385- 4392. 1994(a) .
- Rat an, R. R. ,
Murphy,
T
. H. , Baraban, J. M. . Oxi dat i ve st ress i nduces
apopt osi s i n embryoni c cort i cal neurons. J. Neurochem. ,
62:
376- 379
. 1994(b) .
- Reddel , R. R. . Genes i nvol ved i n t he cont rol of cel l ul ar
prol i f erat i ve pot ent i al .
Ann. N. Y. Acad. Sci . , 854: 8- 19. 1998.
- Rego, A. C. , Ol i vei ra, C. R. . Dual ef f ect of
l i pi d peroxi dat i on ont hemembrane
order of ret i nal cel l s cul t ure. Arch. Bi ochem. Bi ophys. , 321: 127- 136. 1995.
- Rego, A. C. , Sant os, M. S. , Ol i vei ra,
C. R
.
. Adenosi ne t ri phosphat e
degradat i on product s af t er oxi dat i ve
st ress and met abol i c dysf unct i on i n
cul t ured ret i nal cel l s . J. Neurochem. , 69: 1228- 1235. 1997.
- Rego, A. C. , Sant os, M. S. , Ol i vei ra,
C. R. . Oxi dat i ve st ress, hypoxi a and
i schemi a- l i ke condi t i ons i ncrese t he rel ease of endogenous ami noaci ds
by
di st i nct mechani sms i n cul t ured ret i nal cel l s. J. Neurochem. ,
66: 2506- 2516.
1996.
- Ri ceevans, C. , Burdon, R. . Free radi cal l i pi d i nt eract i ons
and t hei r
pat hol ogi cal consequences. Prog. Li pi dRes. , 37: 37- 110. 1. 993.
- Ri cht er, C. . Do mi t ochondri al DNAf ragment s promot e cancer and agi ng?.
FEBSLet t . , 241: 1- 5. 1988
.
- Ri ckwood, D. , Wi l son, M. T. , Darl ey- Usmar, V. M. . Mi t ochondri a: Apract i cal
approach. I RLPress, Oxf ord Washi ngt on, D. C. 1987.
- Robi nson, N. C. . Funct i onal bi ndi ng of cardi ol i pi n
t o cyt ochrome c oxi dase. J.
Bi oenerg. Bi omembr. , 25: 153- 163. 1993.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Bi bl i ograf a
-Rooyackers, O. E. , Adey,
D. B. , Ades, P. A. , Nai r, K. S. . Ef f ect of ageoni nvi vo
rat es of mi t ochondri al prot ei nsynt hesi s i n humanskel et al
muscl e. Proc. Nat l .
Acad. Sci . , 93
: 15364-15369. 1996.
-Rose, M. R. . Canhumanagi ngbepost poned? Sci . Am. , 281
: 106-111. 1999.
-Rose, M. R. . Evol ut i onary
bi ol ogyof agi ng. Oxf ordUni versi t y Press. 1994.
-Rot t kamp,
C. A
. ,
Nunomura, A. , Hi rai , K. . Wi l l ant i oxi dant s f ul l f i l l t hei r
expect at i ons f or t he t reat ment of Al zhei mer di sease?. Mech. Agei ng
Dev. ,
116: 169-179. 2000.
-Rubbo, H. ,
Darl ey-Usmar, V. M. , Freeman, B. A. . Ni t ri c oxi de regul at i on
of
t i ssue f ree radi cal i nj ury. Chem. Res. Toxi col . , 9: 809-820
. 1996.
-Rust i n, P. , von
Kl ei st -Ret zow, J. , Vaj o, Z. , Rot i g, A. , Munni ch, A. .
For
debat e: def ect i ve mi t ochondri a, f ree radi cal s,
cel l deat h, agi ng real i t y or
mi t hochondri a?
. Mech. Agi ngDev. , 114: 201-206. 2000.
-Sagri st a, M. L. , Garci a, A. E. , Af ri ca de Madari aga, M. ,
Mora, M. .
Ant i oxi dant and pro-oxi dant ef f ect
of t he t hi ol i c compounds N-acet yl -L-
cyst ei ne and
gl ut at hi one agai ns f ree radi cal -i nduced l i pi d
peroxi dat i on. Free
Radi c. Res. , 36: 329-340. 2002.
-Samper, E. , Goyt i sol o, F. , Sl i j epcevi c,
P. , vanJul , P. , et al . . Mammal i an
Ku86 prevent s
t el omeri c f usi ons i ndependent l y of t he l engt h
of TTAGGG
repeat s andt he G-st randoverhang. EMBO
Report s, 1: 244-252. 2000.
-Saran,
M. , Bors, W. . Radi cal react i ons i n vi vo. An
overvi ew. Radi at . Envi ron.
Bi ophys. , 29: 249-262
. 1990.
-Sast re, J. ,
Pal l ard, F. V. , Garci a de l a Asunci 6n, J. , Vi a,
J. . Mi t o-
chondri a,
oxi dat i ve st ress andagi ng. FreeRad.
Res. , 32: 189-198. 2000a.
-Sast re, J . , Pal l ard, F. V. ,
Vi a, J. . Mi t ochondri al oxi dat i ve st ress pl ays
a
keyrol e i n
agi ng andapopt osi s. IUBMBLi f e, 49: 427-435.
2000b.
-Sast ry, P. S. , Rao, K. S. . Apopt osi s andt he nervous
syst em. J. Neurochem. ,
74: 1-20. 2000.
-Scadut o,
R. S. . Gl ut at hi one i n i schemi a and
reperf usi on-i nduced t i ssue
i nj ury. En:
Gl ut at hi one met abol i smandphysi ol ogi cal f unct i ons
. Vi a, J . ( Ed) ,
CRC
Press, Boca Rat on, Fl ori da, pgs. 227-235.
1990.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Bi bl i ograf i a
- Schaf er, K. A. . Thecel l cycl e: a
revi ew. Vet . Pathol . , 35: 461- 478. 1998.
- Schai ch, K. H. . Metal s and l i pi d oxi dati on.
Cotemporary i ssues. Li pi ds,
27: 209- 218. 1992.
- Schonei ch, C. . Reacti ve
oxygen speci es andbi ol ogi cal agi ng: a mechani sti c
approach. Exp.
Gerontol . , 34: 19- 34. 1999.
- Sevani an, A. , Stei n, R. A. ,
Mead, J. F. . Metabol i sm of epoxi di zed
phospati dyl chol i ne by phosphol i pase A2 and epoxi de
hydrol ase. Li pi ds,
16: 781- 788. 1981.
- Shaki b, M. ,
Cunha- Vaz, J. G. . Studi es onthepermeabi l i ty of
thebl ood- ocul ar
barri er . I V. Juncti onal compl exes of the reti nal
vessel s andthei r rol e i n the
permeabi l i ty of the
bl ood- reti nal barri er . Exp. EyeRes. , 5: 229- 234. 1966.
- Sharpe,
M. A
. ,
Cooper, C. E. . Reacti ons of ni tri c oxi de wi th
mi tochondri al
cytochrome c: a novel mechani smf or the
f ormati on of ni trosyl ani on and
peroxyni tri te. Bi ochem. J. ,
332: 9- 19. 1998.
- Si es,
H. . Oxi dati veStress. Si es H. ( ed)
Academi c Press,
London. 1. 985.
- Sl ater, T. F. S. . Free radi cal s
f act or f i cti on? . Br. J. Cancer, 55: 5- 10. 1987.
- Smi th, W. L. .
Prostagl andi n bi osynthesi s and i ts
compartmentati on i n
vascul ar smooth muscl e and endothel i al
cel l s. Annu. Rev. Physi ol . , 48: 251-
262
. 1986.
- Snodderl y, D. M. . Evi dence f or protecti on
agai nst age- rel ated macul ar
degenerati on by carotenoi ds and
anti oxi dant vi tami ns. Am. J. Cl i n. Nutr. ,
62
: 1448S- 1461S. 1995.
- Song, Z. , Stel l er, H. . Deathby desi gn: mechani sm
andcontrol of apoptosi s.
Trends Cel l Bi ol . ,
9: M49- M52. 1999.
- Southorn, P. A. , Powi s, G. . Free radi cal s i nmedi ci ne. I . Chemi cal
nature and
bi ol ogi c reacti ons. Cl i n.
Proc. , 63: 381- 389. 1988.
- Spal l hol z, J. E. , Boyl an, L. M. . Gl utathi one peroxi dasa: The
two sel eni um
enzymes. En: Peroxi dases i n
chemi stry andbi ol ogy. Everse, J . , Everse, K. E. ,
Gri sham, M. B. ( eds) , CRCPress, BocaRaton. 1991.
- Stadtman, E. R. . Protei noxi dati onandagi ng. Sci ence, 257: 1220- 1224.
1992.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Bi bl i ograf a
- Staml er, J. S. , Si ngl e, D. , Loscal zo, J. . Bi ochemi stry of ni tri c
oxi de and i ts
redox- acti vatedf orms. Sci ence, 258: 1898- 1902. 1992.
- Stei n- Behrens, B. A. , Spol sky, R. M. . Stress, gl ucocorti coi ds,
andagi ng. Agi ng
Cl i n. Exp. Res
. ,
4
: 197- 210. 1992.
- Stei nberg, R. H. , Mi l l er, S. H. . Transport and membrane properti es of the
reti nal pi gment epi thel i um. I n: The Reti nal Pi gment
Epi thel i um. Zi nn, K. M. ,
Marmor, M. F. , ( Eds) , Cambri dge,
Massachusetts, Harvard Uni versi ty Press,
pgs. 205- 225. 1979.
- Strol i n- Benedetti ,
M. ,
Dostert,
P. , Marrari , P. , Ci ni , M. . Ef f ect of agei ngon
ti ssue l evel s of ami no aci ds i nvol ved i n the ni tri c oxi depathway
i n rat brai n. J.
Neural Transm. Gen.
Sect. ,
94
: 21- 30. 1993.
- Sugrue,
M
. M. , Wang, Y. , Chal mers- Redman, R. M. , et
al
. .
Reduced
mi tochondri a) membranes potenti al and al tered
responsi veness of a
mi tochondri a) membrane megachannel i n
p53- i nduced senescence. Bi ochem.
Bi ophys.
Commun. , 261: 123- 130. 1999.
- Szabo, C. , Zi ngarel l i , B. , O' Connor, M. , Sal zman,
A. . DNAstrandbreakage,
acti vati on of
pol y( ADP- ri bose) Synthetase, and cel l ul ar energy depl eti on are
i nvol ved i n the cytotoxi ci ty i n macrophages and smooth
muscl e cel l s exposed
to peroxyni tri te.
Proc. Natl . Acad. Sci . , 93: 1753- 1758. 1996.
- Szent- Gyorgi A. . Observati ons on the f uncti ons
of peroxi dase systems and
thechemi stryof the
adrenal cortex. Bi ochemJ. , 22: 1387- 1394. 1928.
- Tager, J. M. ,
Wanders, R. J . A. , Groen, A. K. , Kunz, W. , Bohnensack, R. ,
Kster, U. , et al . . Control of mi tochondri al respi rati on.
FEBSLett. , 151: 1- 9.
1983.
- Tagl i al atel a,
G. , P6rez- Pol o,
J. R. , Rassi n, D. K. . I nducti on
of apoptosi s i n
theCNSduri ngdevel opment by the
combi nati onof hyperoxi aandi nhi bi ti on of
gl utathi one
synthesi s. FreeRadi c. Bi ol . Med. , 15: 936- 942. 1998.
- Takasawa, M. , Hayakawa, M. , Sugi yama,
S. , Hattori , K. , I to, T. , Ozawa,
T. .
Age- associ ated damage i n mi tochondri a) f uncti on i n
rat hearts. Exp.
Gerontol . , 28: 269- 280. 1993.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Bi bl i ograf i a
-Tamatani , M. , Ogawa, S. , Ni i tsu, Y. , Tohyama,
M. . I nvol vement of Bcl -2
f ami l y and caspase-3-l i ke protease i n NO-medi ated neuronal apoptosi s.
J.
Neurochem. , 71: 1588-1596. 1998.
-Tappel ,
A. L. , Brown,
V
. D. , Zal ki n, H. , Mai er, V. P. . Unsaturated l i pi d
peroxi dati oncatal yzedbyhemati ncompounds andi ts i nhi bi ti onbyvi tami nE.
J. Am. Oi l . Chem. Soc. , 38: 5-10. 1961.
-Tappel ,
A. L. . Free-radi cal l i pi d peroxi dati on damage andi ts i nhi bi ti on by
vi tami nEandsel eni um. Fed. Proc. , 24: 73-78. 1965.
-Tate, D. J . , Mi cel i , M. V. ,
Newsome, D. A. , Al cock, N. W. , Ol i ver, P. D. .
I nf l uence of zi nc on sel ected cel l ul ar f uncti ons
of
cul tured
human reti nal
pi gment
epi thel i um.
Curr. Eye
Res. , 14: 897-903. 1995
-Terry,
R. D. , DeTeresa, T. , Hansen, L. A. . Neocorti cal cel l count i n
normal
humanadul t agi ng. Ann. Neurol . , 21: 530-539
. 1987.
-Thurnham, D. L. Anti oxi dants and
prooxi dant i n mal nouri shedpopul ati ons.
Proc. Nutr. Soc. , 49: 247-259. 1990.
-Ti an, L. , Cai , Q. , Wei , H. . Al terati ons
of anti oxi dant enzymes andoxi dati ve
damageto
macromol ecul es
i n
di f f erent organs of rats duri ng agi ng. FreeRadi c
.
Bi ol . Med. , 24: 1477-1484. 1998.
-Tosaki ,
A. , Droy-Lepai x, M. T. , Pal i , T. , Das, D. K. . Ef f ects
of SOD, catl ase,
and a novel semi arrhythmi c drug,
EGB 761, on reperf usi on-i nduced
arrhythmi as i n
i sol ated
rat
hearts. FreeRad. Bi ol . Med. , 14: 361-370. 1993.
-Trounce, I . , Byrne, E. , Marzuki , S. . Decl i ne i n
skel etal muscl e mi to-
chondri al respi ratorychai n f uncti on
: possi bl e f actor i n agei ng. Lancet, 1: 637-
639.
1989.
-Trush, M. A. , Mi mnaugh, E. G. , Gram, T. E. . Acti vati on of pharmacol ogi c
agents to radi cal i ntermedi ates.
I mpl i cati ons f or the rol es of f ree radi cal s i n
drug acti onandtoxi ci ty. Bi ochem. Pharmac. , 31: 3335-3343. 1982.
-Turrens,
J. F. , Crapo,
J. D. ,
Freeman, B. A. . Protecti on agai nst oxygen
toxi ci ty by i ntravenous i nj ecti ons of l i posome-entrapped catal ase and
superoxi de di smutasa. J. Cl i n. I nvest. , 73: 87-95.
1984.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Bi bl i ograf a
- Val enzuel a, A. , Vi del a, L. A. . Formas act i vas del oxi geno, est rs oxi dat i vo y
suproyecci n pat ol gi ca. Rev. Med. Chi l e, 117: 60- 67. 1989.
- Val enzuel a, A. .
The bi ol ogi cal si gni fi cance of
mal ondi al dehi de det ermi nat i on
i n t heassessment of t i ssue oxi dat i ve st ress. Li fe sci . , 48: 301- 309. 1991.
- Van St eensel , B. , Smogorzawska, A. , de Lange, T. . TRF2prot ect s human
t el omers fromend- t o- end fusi ons. Cel l , 92: 401- 413. 1998.
- Vanfl et eren, J. R. , Braeckman, B. P. . Mechani sms of l i fe span det ermi nat i on
i n Caenorhabdi t i s el egans. Neurobi ol . Agi ng, 20: 487- 502. 1999.
- Vi j g, J. , Mul l er, W. E. G. . The sci ence of agi ngandt heneed for amechani st i c
approach.
Mech.
Agei ngDev. , 114: 1- 3. 2000.
- Vi k, S. B. , Georgevi ch, G. , Capal di , R. A. . Di phosphat i dyl gl ycerol i s requi red
for opt i mal act i vi t y of beef heart cyt ochrome c oxi dase. Proc. Nat l . Acad.
Sci . ,
78: 1456- 1460. 1980.
- Vi a J . (Ed. ) . Gl ut at hi one: Met abol i smand Physi ol ogi cal Funct i ons. CRC
Press, BocaRat on. 1990.
- Vi rag, L. , Sal zman, A. L. , Szabo, C. . Pol y(ADP- ri bose) synt het ase act i vat i on
medi at es mi t ochondri al
i nj uryduri ngoxi dant - i nduced cel l deat h. J. I nmunol . ,
161
: 3753- 3759. 1998.
- Vi t ori ca, J. , Cano, J. , Sat rust egui , J. , Machado, A. .
Compari son bet ween
devel opment al and
senescent changes i n enzyme act i vi t i es l i nked t o energy
met abol i smi n rat heart . Mech. Agei ngDev. , 16: 105- 116. 1981.
- Webst er, N. R. , Nunn, J. P. . Mol ecul ar st ruct ure of free radi cal s and
t hei r
i mport ance i n
bi ol ogi cal react i ons. Br. J. Anaest h. , 60: 98- 108. 1988.
- Wei ss, H. , Li nke, P. , Hai ker, H. , Leonard, K. . St ruct ure
andfunct i on of t he
mi t ochondri al
ubi qui nol : cyt ochrome c reduct asa and NADH: ubi qui none
reduct asa. Bi ochem. Soc. Trans. , 15: 100- 102
.
1987.
- Wei ssman, A. .
Essays uponheredi t yandki ndred bi ol ogi cal probl ems. Oxford
Uni versi t y- Cl aredon (Eds) , London- NewYork. 1891.
- Wel l s, W. W. , Jung, C. . Regenerat i on
of vi t ami n C. En: Vi t ami n Ci n heal t h
anddi sease. Packer,
L. , Fuchs, J. , (Eds) , Marcel Dekker, I nc. NewYork, pgs.
109- 121.
1997
.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Bi bl i ograf i a
- Wel sh, O. . Studyof af ami l ywi th atypi c progeri a. Mod. Probl . Pedi atr. , 17: 44-
58. 1975.
- Wendel , A. . Gl utathi one peroxi dasa. En: Enzymati c basi s of detoxi cati on
.
J acoby, W. B. , Bend, J . R. , Cal dwel l , J . ( eds) ,
Academi c Press, NewYork. 1980.
- Wharton, D. C. , Gri f f i th,
D. E
. .
Studi es of the el ectron transport system.
Assayof cytochrome oxi dase. Ef f ect of phosphol i pi ds and
other f actors. Arch.
Bi ochem. Bi ophys. , 96: 103- 114. 1965.
- Wharton, D
. C
. ,
Tzagal of f , A. . Cytochrome oxi dase f rom beef heart
mi tochondri a. Methodsi n enzymol ogy, 10: 245- 250. 1967.
- Whi tehouse, P. J . , Pri ce, D. L. , Strubl e,
R. G. , Cl ark, A. , et al . . Al zhei mer
di sease and seni l e dementi a: l oss of neurons i n basal
f orebrai n. Sci ence,
215: 1237- 1239. 1982
.
- Wi l l i ams, A. J . , Coakl ey, J . , Chri stodoul ou, J . . Automated
anal ysi s of
mi tochondri al enzymes i n cul turedski n
f i brobl asts. Anal . Bi ochem. , 259: 2176-
2180. 1998.
- Wi l l i ams, D. L. H. . Ni trosati on. Cambri dge Uni versi ty
Press. Cambri dge, 1. 988.
- Wi nkl er, B. S. , Boul ton, M. E. , Gottsch, J . D. ,
Sternberg, P. . Oxi dati ve
damageandage- rel atedmacul ar
degenerati on. Mol . Vi s. , 3: 32- 38. 1999.
- Wi nkl er, B. S. . Gl ycol yti c and oxi dati ve
metabol i sm i n rel ati on to reti nal
f uncti on. J . Gen. Physi ol
. , 77: 667- 692. 1981.
- Wi nkl er, B. S. . Rel ati ve i nhi bi tory ef f ects of
ATP depl eti on, ouabai n and
cal ci umonreti nal
photoreceptors . Exp. EyeRes. , 36: 581- 594. 1983.
- Wi sl ocki , G. B. , Ladman, A. J . . Thedemonstrati on
of abl ood- ocul ar barri er i n
the al bi no rat by means of the
i ntravi tamdeposi ti on of si l ver . J. Bi ophys.
Bi ochem. Cytol . , 1: 501- 510. 1955.
- Wol f f , S. P. , Dean R. T. . Fragmentati onof protei n by
f ree radi cal s andi ts
ef f ect on thei r suscepti bi l i ty to enzymi c hydrol ysi s
. Bi ochem. J. , 2234: 399- 403.
1986.
- Wol pp, S. P. , Garner A. , Dean, R. T. .
Free radi cal s l i pi ds and protei n
degradati on. Trends Bi ochem. Sci . , 11: 27- 31. 1986.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Bi bl i ograf a
-Wong, K. K. , Chang, S. , Wei l er, S. R. , Ganesan, S. , et al . . Tel omere
dysf uncti oni mpai rs DNArepai r and
enhances sensi ti vi ty to i oni si ng radi ati on.
Nat
. Genet. , 1 : 85-88. 2000.
-Wood, J. P. , Osborne, N. N. . I nducti on
of apoptosi s i ncul tured humanreti nal
pi gmented epi thel i al cel l s: the ef f ect of protei n ki nase C acti vati on and
i nhi bi ti on. Neurochem. I nt. , 31 : 261- . 1997.
-Wul l ner, U. , Kornhuber, J. , Wel l er, M. , Schul z, J. B. , et al . . Cel l death
and
apoptosi s regul ati ng protei ns i n Parki nson' s di sease-a
cauti onary note. Acta
Neuropathol . , 97: 408-412
. 1999.
-Yagi , T. , Hatef i , Y. .
Thi ol s i n oxi dati ve phosphoryl ati on: i nhi bi ti on and
energy-potenti ateduncopl i ngby monothi ol and
di thi ol modi f i ers. Bi ochemi stry,
23: 2449-2455. 1984.
-Yan, C. Y. I . , Greene, L. A. . Preventi on of PC12 cel l death
by N-acetyl cystei ne
requi res acti vati onof theRas pathway.
J. Neurosci . , 18: 4042-4049. 1998.
-Yen,
T, C. , Chen, Y. S. , Ki ng, K. L. , Yeh, S. H. , Wei , Y. H. .
Li ver mi tochondri al
respi ratory f uncti ons
decl i ne wi th age. Bi ochem. Bi ophys. Res.
Commun. ,
165
: 944-1003. 1989.
-Young, R. W. , Bok, D. . Metabol i smof thereti nal pi gment
epi thel i um. En: The
Reti nal Pi gment Epi thel i um. Zi nn,
K. M. , Marmor, M. F . , ( Eds) , Cambri dge,
Massachusetts,
HarvardUni versi ty Press, pgs. 103-123. 1979
.
-Yu, B. P. , Yang, R. . Cri ti cal eval uati on
of the f ree radi cal theory of agi ng: A
proposal f or
the oxi dati ve stress hypothesi s. Ann. N. Y. Acad. Sci . ,
786: 1-11.
1996.
-Yu, B. P. .
Approaches to anti -agi ng i nterventi on: the
promi ses and the
uncertai nti es. Mech. Agei ngDev. , 111
: 279-285. 1999.
-Zarbi n, M. A. .
Age-rel ated macul ar degenerati on: revi ewof pathogenesi s
. Eur.
J. Ophthal mol . ,
8: 199-206. 1998.
-Zhang, L. , Kokkonen, G. , Roth,
G. S
. .
I denti f i cati on of programmed cel l
deathi n si tu i n the stri atum
of normal adul t rat brai nandi ts
rel ati onshi p to
neuronal deathduri ngagi ng. Brai nRes. , 677: 177-179
. 1995.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Bi bl i ograf i a
-Zhang, Y. , Marci l l at , O. , Gi ul i vi , C. , Ernst er, L. , Davi es, K. J . A. . The
oxi dat i ve i nact i vat i on of mi t ochondri al el ect ron t ransport chai n component s
andATPase. J . Bi ol . Chem. , 265: 16330-16336. 1990.
-Zi gl er, J . S. , J erni gan, H. M. J r . , Garl and, D. , Reddy, V. N. .
The ef f ect s of
"oxygen
radi cal s" generat ed i n t he medi umon l enses i n organ cul t ure:
i nhi bi t i on of damage bychel at ed i ron. Arch. Bi ochem. Bi ophys. , 241: 163-172
.
1985.
Modelos de envejecimiento in vitro e in vivo, estrs oxidativo y proteccin antioxidante. Mara del Mar Castillo Paterna
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2002
Volver al ndice/Tornar a l'ndex
Вам также может понравиться
- Abascal Vicente, M DoloresДокумент659 страницAbascal Vicente, M DoloresAlba MarinaОценок пока нет
- Bioquimica IApoyo EductivoДокумент96 страницBioquimica IApoyo EductivodavidОценок пока нет
- PLAN CONSERVACION Bosques Pino-EncinoДокумент103 страницыPLAN CONSERVACION Bosques Pino-EncinoBerenice ListОценок пока нет
- Frontera Quechua en ApurímacДокумент0 страницFrontera Quechua en ApurímacMarcos van LukañaОценок пока нет
- Yianatos - Flotación de Minerales (UTFSM)Документ126 страницYianatos - Flotación de Minerales (UTFSM)Francisco Javier Alarcón Garrido100% (7)
- TS CampotElolaMariaДокумент49 страницTS CampotElolaMariaValeria BarreraОценок пока нет
- La Delgada Línea RosaДокумент39 страницLa Delgada Línea RosaJulio Cesar MacielОценок пока нет
- Tesis PCДокумент97 страницTesis PCJherson Ariza GarciaОценок пока нет
- Tecnología de Los Alimentos Vol I Componentes de Los Alimentos y Procesos - Juan OrdoñezДокумент357 страницTecnología de Los Alimentos Vol I Componentes de Los Alimentos y Procesos - Juan OrdoñezSol Luis100% (3)
- Bioestimulantes en CacaoДокумент85 страницBioestimulantes en CacaoAnonymous ABvPjp100% (1)
- Fernando Favela - Viaje A Las Estrellas en Agujeros de GusanoДокумент73 страницыFernando Favela - Viaje A Las Estrellas en Agujeros de GusanoALEISTERTHОценок пока нет
- Metodosdeestudiodevidadeanaqueldelosalimentos PDFДокумент106 страницMetodosdeestudiodevidadeanaqueldelosalimentos PDFdanny elisetОценок пока нет
- Fisica Comun PsuДокумент226 страницFisica Comun PsuMartín Soria ElsnerОценок пока нет
- Wmo 327-s Climatologia PDFДокумент179 страницWmo 327-s Climatologia PDFwalterdryОценок пока нет
- Biologia - Teoria y Metodos en Geografia FisicaДокумент152 страницыBiologia - Teoria y Metodos en Geografia FisicaCristián Monzon100% (1)
- Estimacion de La Resistencia Al Corte de M - Alb - ArmadaДокумент286 страницEstimacion de La Resistencia Al Corte de M - Alb - ArmadaCalculistaОценок пока нет
- 17MIBI - Medicina Regenerativa - ManualДокумент82 страницы17MIBI - Medicina Regenerativa - ManualAnonymous UAPowbM50Оценок пока нет
- Propuesta Metodólogica para Evaluación Fisioterapeútica de Transtornos Motores en Secuelas de HemДокумент125 страницPropuesta Metodólogica para Evaluación Fisioterapeútica de Transtornos Motores en Secuelas de HemNaike SakaeОценок пока нет
- Plagas y Enfermedades Del OlivoДокумент94 страницыPlagas y Enfermedades Del OlivoGregorio Rozas Fernández100% (2)
- Teoria de Bioquimica UamДокумент95 страницTeoria de Bioquimica UamRodrigo Martinez50% (2)
- Rosceliavitiamartell Diseño2 Casamundointerno Libreto-CombinadoДокумент44 страницыRosceliavitiamartell Diseño2 Casamundointerno Libreto-CombinadoKevin SolisОценок пока нет
- El Maltrato Infantil Un Análisis Ecológico de Los Factores de Riesgo PDFДокумент495 страницEl Maltrato Infantil Un Análisis Ecológico de Los Factores de Riesgo PDFRubenHernandoОценок пока нет
- Aceites Gildardo PDFДокумент174 страницыAceites Gildardo PDFSantiago PossoОценок пока нет
- HugoA LeónGarzón 2017Документ98 страницHugoA LeónGarzón 2017Heidy PCОценок пока нет
- Diferenciacion SexualДокумент90 страницDiferenciacion SexualJason ChávezОценок пока нет
- Informe Incinerador Doble CamaraДокумент18 страницInforme Incinerador Doble CamaraHitler Ivan Chacon QuispeОценок пока нет
- Atlas de HelmintosДокумент153 страницыAtlas de HelmintosGutierrez AleОценок пока нет
- Atlas Helmintos PDFДокумент182 страницыAtlas Helmintos PDFObarduckОценок пока нет
- PDF Civic A CompressДокумент140 страницPDF Civic A Compressssjcherub58Оценок пока нет
- Manual UPV Abusos Sexuales Contra Menores de EdadДокумент206 страницManual UPV Abusos Sexuales Contra Menores de EdadNicolás Pereira ElguetaОценок пока нет
- AO de ColumnaДокумент244 страницыAO de ColumnadafneferradaОценок пока нет
- 05MBIN MdeToroДокумент159 страниц05MBIN MdeToroJavier F Rosero MОценок пока нет
- Universidad Nacional Agraria: Facultad de AgronomiaДокумент130 страницUniversidad Nacional Agraria: Facultad de AgronomiaJEAN CARLOS AVILES TOLEDOОценок пока нет
- Notas ActuarialesДокумент47 страницNotas ActuarialesPavel PachecoОценок пока нет
- Apuntes Fluidos12Документ99 страницApuntes Fluidos12Francisca Sánchez DonosoОценок пока нет
- Manual Modulo 3 MA y MN 13Документ60 страницManual Modulo 3 MA y MN 13JOSE ARMANDO REYES OCHOAОценок пока нет
- Biotipo Facial y Patrón Esqueletal Predominante en Pobladores de 18 - 25 AñosДокумент58 страницBiotipo Facial y Patrón Esqueletal Predominante en Pobladores de 18 - 25 Añoshector rinconОценок пока нет
- PDF Practica Determinacion Del Contenido de Saponinas en La Quinua - CompressДокумент18 страницPDF Practica Determinacion Del Contenido de Saponinas en La Quinua - CompressSantiago DangerОценок пока нет
- Cianobacterias Planctonicas en Embalses Españoles - Carrasco - DДокумент320 страницCianobacterias Planctonicas en Embalses Españoles - Carrasco - DAlfredo SuárezОценок пока нет
- Quispe Tito Maycool YonnathanДокумент264 страницыQuispe Tito Maycool YonnathanLucero Vasquez CamposОценок пока нет
- Introducción A La Teoría de NudosДокумент57 страницIntroducción A La Teoría de NudosFavio90100% (1)
- UntitledДокумент117 страницUntitledRenzo Alan Santana BilbaoОценок пока нет
- MORENO NORBERTO. Relaciones Semánticas.Документ36 страницMORENO NORBERTO. Relaciones Semánticas.Mer Fernandez LunaОценок пока нет
- Teoria de GaloisДокумент142 страницыTeoria de Galoisjhorckham100% (1)
- Tesisi de Grado Las PymesДокумент117 страницTesisi de Grado Las PymesGabriel Antonio CanalesОценок пока нет
- Geografi LimitrofeДокумент123 страницыGeografi LimitrofeGrover LimaninОценок пока нет
- Tesis Isabella AquiqueДокумент144 страницыTesis Isabella AquiquetesisdaniyuliОценок пока нет
- Conservación Del Jugo de Naranja Mediante 3 ProtocoloДокумент61 страницаConservación Del Jugo de Naranja Mediante 3 ProtocoloVivian LMОценок пока нет
- Libro LeishmanДокумент200 страницLibro LeishmanDorina Rom-SalОценок пока нет
- Comportamiento Humano: El recurso básico de las organizaciones empresarialesОт EverandComportamiento Humano: El recurso básico de las organizaciones empresarialesОценок пока нет
- Medio ambiente y su protección jurídica: Diálogos doctrinarios y jurisprudenciales en el siglo XXIОт EverandMedio ambiente y su protección jurídica: Diálogos doctrinarios y jurisprudenciales en el siglo XXIОценок пока нет
- La acción merodeclarativa y su importancia en materia de concubinatoОт EverandLa acción merodeclarativa y su importancia en materia de concubinatoОценок пока нет
- Métodos y Técnicas de Investigación Social: Manual para principiantesОт EverandMétodos y Técnicas de Investigación Social: Manual para principiantesОценок пока нет
- 2do Parcial GuiaДокумент5 страниц2do Parcial GuiaGuillermo ValdezОценок пока нет
- 4to de Secundaria (Sebastian Balcazar Montano)Документ28 страниц4to de Secundaria (Sebastian Balcazar Montano)Sebastian Balcazar Montaño100% (1)
- MembranaДокумент35 страницMembranaIsa GarciaОценок пока нет
- Examen Farmaco 1 Fase PDFДокумент181 страницаExamen Farmaco 1 Fase PDFmarisol andrea ito ytoОценок пока нет
- Fisiología HepáticaДокумент17 страницFisiología Hepáticamkarla23Оценок пока нет
- Carbohidratos Informe CientificoДокумент6 страницCarbohidratos Informe Cientificojtcp27031Оценок пока нет
- Fluorocromos 123Документ4 страницыFluorocromos 123Adrees CastroОценок пока нет
- DisacaridosДокумент6 страницDisacaridosCariiThoFlorОценок пока нет
- Capítulos 1, 2, 3, 4, 5, 6, 7 y 8 de AnatomiaДокумент171 страницаCapítulos 1, 2, 3, 4, 5, 6, 7 y 8 de AnatomiaLeo C GОценок пока нет
- La GenéticaДокумент28 страницLa GenéticaVirginia Bornachera LopezОценок пока нет
- HORMONASДокумент4 страницыHORMONASSofía VillaОценок пока нет
- Organitos Membranosos y No Membranosos e Inclusiones CelularesДокумент19 страницOrganitos Membranosos y No Membranosos e Inclusiones CelularesEktor Yvan HdezОценок пока нет
- Bibliografia Cel Procarioticas y Eucarioticas Clase 2Документ8 страницBibliografia Cel Procarioticas y Eucarioticas Clase 2Nancy BertucciОценок пока нет
- ABP 2 Recordando-AndoДокумент49 страницABP 2 Recordando-AndoMaholy CherrezОценок пока нет
- PlastidosДокумент18 страницPlastidosKenia GuzmánОценок пока нет
- Resumen Sobre BiopotencialesДокумент3 страницыResumen Sobre BiopotencialesLeonardo Andres Acevedo AvilaОценок пока нет
- Propiedades de La Vitamina BДокумент4 страницыPropiedades de La Vitamina BJonathan VasquezОценок пока нет
- Quimica L 1Документ8 страницQuimica L 1Diego Linero RamirezОценок пока нет
- Síndrome X FrágilДокумент12 страницSíndrome X Frágilthalia asencios aguirreОценок пока нет
- Lipidos y MetabolismoДокумент45 страницLipidos y Metabolismokona chanОценок пока нет
- 3 PruebaДокумент3 страницы3 PruebaFrancisca Lehandra Hernandez IturraОценок пока нет
- Metabolismo de Lípidos y ProteínasДокумент3 страницыMetabolismo de Lípidos y ProteínasGabriela Lizeth Escobar SosaОценок пока нет
- Fichas Tecnicas PhysiologyДокумент28 страницFichas Tecnicas PhysiologyAlexia jocelyne Flores fuerteОценок пока нет
- GeneticaДокумент1 страницаGeneticaSalo EstrellaОценок пока нет
- Farmacoterápia Resumen Segundo Parcial.Документ136 страницFarmacoterápia Resumen Segundo Parcial.Fernando FloresОценок пока нет
- Metabolismo de Nucleótidos y Aciduria OróticaДокумент20 страницMetabolismo de Nucleótidos y Aciduria OróticaCATALINA ANDREA ROJAS ARMAZAОценок пока нет
- LLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLДокумент30 страницLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLLdulce100% (1)
- Sistema Renina Angiotensina AldosteronaДокумент12 страницSistema Renina Angiotensina AldosteronaVicente Fernando Arizabal CarpioОценок пока нет
- Tipos de Receptores Celulares y Mecanismos de TransducciónДокумент38 страницTipos de Receptores Celulares y Mecanismos de TransducciónKariem Noelia Sherezade Siquero Vera100% (1)
- Medios de CultivoДокумент24 страницыMedios de CultivoJorge LambaréОценок пока нет