Вам также может понравиться
- Ditado de Palavras Com RRДокумент9 страницDitado de Palavras Com RRNivea Neves100% (6)
- Efeito imediato da acupuntura na lombalgia: Sistema Yamamura de Acupuntura do Osso Nasal e do ponto craniométrico lambdaОт EverandEfeito imediato da acupuntura na lombalgia: Sistema Yamamura de Acupuntura do Osso Nasal e do ponto craniométrico lambdaОценок пока нет
- Causa Emocionais Das DoençasДокумент8 страницCausa Emocionais Das DoençasAmiantusОценок пока нет
- #Lista de PalavrasДокумент18 страниц#Lista de PalavrasMarcelo Asensi Bordin100% (1)
- Fisiopatologia Da Dor - Aula CompletaДокумент55 страницFisiopatologia Da Dor - Aula CompletaHelio Pinheiro100% (1)
- Inglês - Volume 2Документ211 страницInglês - Volume 2lucaroxa23100% (2)
- Semiologia e Fisiopatologia Da DorДокумент44 страницыSemiologia e Fisiopatologia Da Dorlaura solettiОценок пока нет
- A cannabis nas patologias do sistema nervoso centralОт EverandA cannabis nas patologias do sistema nervoso centralОценок пока нет
- Phytoterapica Catalogo-Primavera PDFДокумент48 страницPhytoterapica Catalogo-Primavera PDFsadeyes_19Оценок пока нет
- Material Ergonomia e Ginástica LaboralДокумент18 страницMaterial Ergonomia e Ginástica LaboralalexmeioambienteОценок пока нет
- 4 - Ecologia - Relações EcológicasДокумент14 страниц4 - Ecologia - Relações EcológicasAlison JabesОценок пока нет
- Prova Prática GOДокумент9 страницProva Prática GOFabrício Fabri100% (1)
- 1 - Clinica Veterinaria e Pet Shop Ki LateДокумент30 страниц1 - Clinica Veterinaria e Pet Shop Ki LateMárcia LandiniОценок пока нет
- Atividade Formativa - Verbos Significativos e de Ligação - 7º Ano AmДокумент2 страницыAtividade Formativa - Verbos Significativos e de Ligação - 7º Ano AmYuri Pereira100% (1)
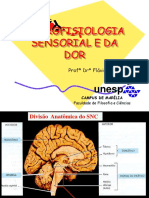
- Neuroanatomia Da DorДокумент5 страницNeuroanatomia Da DorRoberto HangleyОценок пока нет
- Instalação de Prótese TotalДокумент8 страницInstalação de Prótese TotalCristiane SantosОценок пока нет
- Atividade Avaliativa de Língua Portuguesa 5º Ano PDFДокумент3 страницыAtividade Avaliativa de Língua Portuguesa 5º Ano PDFBRUNA ROBERTA GOTTEMSОценок пока нет
- Avaliação do Controle Autonômico Baseada na Análise da Variabilidade da Frequência Cardíaca: Proposta de Método Otimizado para Ativação Vagal e Aplicação da Ferramenta para Estimativa da Dor em Intervenções ClínicasОт EverandAvaliação do Controle Autonômico Baseada na Análise da Variabilidade da Frequência Cardíaca: Proposta de Método Otimizado para Ativação Vagal e Aplicação da Ferramenta para Estimativa da Dor em Intervenções ClínicasОценок пока нет
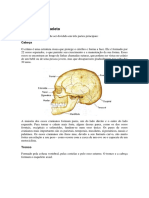
- Atlas de NeuroanatomiaДокумент97 страницAtlas de Neuroanatomiamanukense100% (3)
- Tecido MuscularДокумент32 страницыTecido MuscularJoselma Pinheiro100% (1)
- Farmacologia Da Dor e InflamaçãoETHOSFARMAДокумент54 страницыFarmacologia Da Dor e InflamaçãoETHOSFARMALuiz Gustavo Peron MartinsОценок пока нет
- Fisiopatologia Da Nocicepção e Da Supressão Da DorДокумент6 страницFisiopatologia Da Nocicepção e Da Supressão Da DoreduardaОценок пока нет
- Artigo Dor - Dr. Manoel e Dra. HelenaДокумент15 страницArtigo Dor - Dr. Manoel e Dra. HelenajusclОценок пока нет
- Fisiopatologia Da DorДокумент12 страницFisiopatologia Da DorSandro LimaОценок пока нет
- Dor e AnalgesiaДокумент7 страницDor e AnalgesiaLara TavaresОценок пока нет
- Trends ImmunologyДокумент31 страницаTrends ImmunologyRogerio MachadoОценок пока нет
- Resumo de DorДокумент11 страницResumo de DorLuciana100% (1)
- Semiologia Da DorДокумент12 страницSemiologia Da DorJoão PauloОценок пока нет
- Trilha Semiologia Da DorДокумент12 страницTrilha Semiologia Da DorCAMILA EUQUERES PARTATAОценок пока нет
- 2.fisiopatologia Da Dor - Luiz FernandoДокумент5 страниц2.fisiopatologia Da Dor - Luiz FernandopmontanherОценок пока нет
- Tutoria 1.2Документ31 страницаTutoria 1.2juniorblack_7Оценок пока нет
- Dor ResumoДокумент4 страницыDor ResumoGee AndlerОценок пока нет
- ASHMAWI, H. A. FREIRE, G. N. G. Peripheral and Central Sensitization. São Paulo Revista Dor. v.17, n.3, p.31-34, 2016.Документ4 страницыASHMAWI, H. A. FREIRE, G. N. G. Peripheral and Central Sensitization. São Paulo Revista Dor. v.17, n.3, p.31-34, 2016.Fran WermannОценок пока нет
- Devolutiva Problema 1 DORДокумент8 страницDevolutiva Problema 1 DORMatheus NicolettiОценок пока нет
- Fisiologia Da Dor PDFДокумент3 страницыFisiologia Da Dor PDFGrisel Ibáñez100% (1)
- Neuralgia Do TrigêmeoДокумент3 страницыNeuralgia Do Trigêmeogiuliaoliveira98hotmail.comОценок пока нет
- FIBROMIALGIAДокумент17 страницFIBROMIALGIARodrigo RibeiroОценок пока нет
- Dor - Anatomofisiologia e MecanismosДокумент51 страницаDor - Anatomofisiologia e MecanismosWallberthy Linhares100% (1)
- Aspectos Neuropsicofisiológicos Da HipnoseДокумент6 страницAspectos Neuropsicofisiológicos Da Hipnoseanderson11066100% (1)
- Nociceptores e Hiperalgesia.Документ32 страницыNociceptores e Hiperalgesia.Kedma VitóriaОценок пока нет
- Aula 2 Parte I - Nocicepção e TemperaturaДокумент15 страницAula 2 Parte I - Nocicepção e TemperaturaPatrícia SilvaОценок пока нет
- Cespe 2009 Ms Medico Clinica Da Dor ProvaДокумент4 страницыCespe 2009 Ms Medico Clinica Da Dor ProvaCésar OliveiraОценок пока нет
- Capítulo 01 - Aspectos Anatômicos e FisiopatológicosДокумент7 страницCapítulo 01 - Aspectos Anatômicos e FisiopatológicosSandro PerilloОценок пока нет
- DORДокумент5 страницDORSoraia GonçalvesОценок пока нет
- Farmacologia Dos Anestesicos LocaisДокумент15 страницFarmacologia Dos Anestesicos Locaisiti_tur100% (2)
- HarrisonДокумент1 351 страницаHarrisonMaria LimaОценок пока нет
- Harisson DorДокумент113 страницHarisson DorRangellwlОценок пока нет
- 179-Textos Fonte-436-1-10-20150303Документ21 страница179-Textos Fonte-436-1-10-20150303me687020Оценок пока нет
- 02 - Aspectos Neurofisiologicos Da DorДокумент54 страницы02 - Aspectos Neurofisiologicos Da DorFlávio Martins100% (1)
- 9 Fisiologia Da DorДокумент8 страниц9 Fisiologia Da DorLarissa Sayuri YoshizawaОценок пока нет
- Vias AscendentesДокумент9 страницVias AscendentesLucianaОценок пока нет
- Sensibilização Central - GraduaçãoДокумент14 страницSensibilização Central - GraduaçãoHenrique MachadoОценок пока нет
- Nociceptor Interações Sensoriais Neurônio-Imune Na Dor e InflamaçãoДокумент42 страницыNociceptor Interações Sensoriais Neurônio-Imune Na Dor e InflamaçãoRogerio MachadoОценок пока нет
- Captura de Tela 2020-11-18 À(s) 11.41.33Документ47 страницCaptura de Tela 2020-11-18 À(s) 11.41.33Angelo TorresОценок пока нет
- Fisiopatologia Da Dor PDFДокумент23 страницыFisiopatologia Da Dor PDFSérgioD'LimaОценок пока нет
- A Fisiologia Da Dor: IntroduçãoДокумент12 страницA Fisiologia Da Dor: Introduçãoesmael franciscoОценок пока нет
- Semiologia Da DorДокумент13 страницSemiologia Da DorEduardo TavaresОценок пока нет
- Visao Geral Sistema SensitivoДокумент38 страницVisao Geral Sistema SensitivoPriscilla Santos100% (1)
- Resenha DOR - Julho 1 OkДокумент29 страницResenha DOR - Julho 1 OkRayan Orlando KinalskiОценок пока нет
- Neurofisiologia Do Processo de DorДокумент91 страницаNeurofisiologia Do Processo de DorLu Nogueira100% (1)
- Aula DorДокумент20 страницAula DorRafel FisioОценок пока нет
- P1 TutoriaДокумент6 страницP1 TutoriaMaria Eduarda GeneraliОценок пока нет
- Artigo DorДокумент12 страницArtigo DorScientifica EducacionalОценок пока нет
- fISIOPATOLOGIA DA DOR PDFДокумент12 страницfISIOPATOLOGIA DA DOR PDFFisiologia IОценок пока нет
- TeoricoДокумент22 страницыTeoricoAlicia TavaresОценок пока нет
- Módulo I - Dor Problema 2 - Assombração Objetivos: Lorena de Souza Santos Medicina - Turma XX 5 Etapa - 2º Semestre 2015Документ6 страницMódulo I - Dor Problema 2 - Assombração Objetivos: Lorena de Souza Santos Medicina - Turma XX 5 Etapa - 2º Semestre 2015Anuah Mansur AbudОценок пока нет
- Dor CronicaДокумент17 страницDor CronicaJéssica MendesОценок пока нет
- Temática - Ciências da Vida - Relação e CoordenaçãoОт EverandTemática - Ciências da Vida - Relação e CoordenaçãoОценок пока нет
- Sindromes Extrapiramidais 2013 PDFДокумент43 страницыSindromes Extrapiramidais 2013 PDFKelly SchneiderОценок пока нет
- Antimicrobiano - MEDCELДокумент12 страницAntimicrobiano - MEDCELKelly SchneiderОценок пока нет
- Assistencia Pre Natal PDFДокумент140 страницAssistencia Pre Natal PDFKelly SchneiderОценок пока нет
- Diretrizes e HDA e HDB 2013Документ14 страницDiretrizes e HDA e HDB 2013ericanurseОценок пока нет
- Afetividade PDFДокумент3 страницыAfetividade PDFKelly SchneiderОценок пока нет
- Cefaleia Cronica DiariaДокумент9 страницCefaleia Cronica DiariaKelly SchneiderОценок пока нет
- Dor Cronica Persistente SBEDДокумент11 страницDor Cronica Persistente SBEDKelly SchneiderОценок пока нет
- Fisiologia CoagulacaoДокумент9 страницFisiologia CoagulacaoaxsantosОценок пока нет
- Anatomia Das Vias NociceptivasДокумент6 страницAnatomia Das Vias NociceptivasKelly SchneiderОценок пока нет
- Reparação TecidualДокумент5 страницReparação TecidualKelly SchneiderОценок пока нет
- Abc Do SusДокумент10 страницAbc Do Susgdr01Оценок пока нет
- Acolhimento Aos UsuariosДокумент3 страницыAcolhimento Aos UsuariosKelly SchneiderОценок пока нет
- SUSДокумент1 страницаSUSKelly SchneiderОценок пока нет
- Guia PassaroДокумент8 страницGuia PassaroceuvazОценок пока нет
- Cards Deformidades CongênitasДокумент11 страницCards Deformidades CongênitasAndré Costa Jr.Оценок пока нет
- Atividade HalloweenДокумент13 страницAtividade Halloweensandroneca100% (1)
- Desenvolvimento Da OclusãoДокумент14 страницDesenvolvimento Da OclusãoEstudos VitóriaОценок пока нет
- Aula 6 (1) (1) - MamiferosДокумент64 страницыAula 6 (1) (1) - MamiferosJoel CordovaОценок пока нет
- EsqueletoДокумент3 страницыEsqueletoJoaquim Nhama Manuel ChirariroОценок пока нет
- Termos Técnicos Zoologia m3 QualquercoiusateamobrabulicoДокумент3 страницыTermos Técnicos Zoologia m3 QualquercoiusateamobrabulicoRodrigo Cesário Pereira SilvaОценок пока нет
- Músculos Da Mímica Facial 2024-1Документ56 страницMúsculos Da Mímica Facial 2024-1Lívia Bremenkamp CorsiniОценок пока нет
- Plural Dos Substantivos CompostosДокумент6 страницPlural Dos Substantivos CompostoskathyОценок пока нет
- Conto e Crônica Contos de Fadas e FábulaДокумент35 страницConto e Crônica Contos de Fadas e FábulaPéricles RoberthОценок пока нет
- Aula Musculatura Do Torax Do Equino (Comentada) - HiankaДокумент15 страницAula Musculatura Do Torax Do Equino (Comentada) - HiankaVanessa Sampaio AlvesОценок пока нет
- DND Magias DruidaДокумент7 страницDND Magias DruidamalkadarkОценок пока нет
- PR Tica 1Документ10 страницPR Tica 1DiogoFevereiroОценок пока нет
- Posso Orar Por VoceДокумент1 страницаPosso Orar Por VoceOdeton SilvaОценок пока нет
- Aura - Aula 2 Sistema EsqueléticoДокумент38 страницAura - Aula 2 Sistema EsqueléticoKayllanne GomesОценок пока нет
- Relatório de Zoologia de Vertebrados Nº2Документ6 страницRelatório de Zoologia de Vertebrados Nº2matluaОценок пока нет
- Provérbios Africanos - WikiquoteДокумент7 страницProvérbios Africanos - Wikiquotesamwellson ismaelОценок пока нет