Вам также может понравиться
- Floudas2016 Article NewInsightsIntoIL-10DependentA PDFДокумент9 страницFloudas2016 Article NewInsightsIntoIL-10DependentA PDFKleber FelipeОценок пока нет
- 2588 FullДокумент11 страниц2588 FullManuel SolisОценок пока нет
- Sle 3Документ8 страницSle 3Dianu GutiérrezОценок пока нет
- The Pathophysiology of Malarial Anaemia: Where Have All The Red Cells Gone?Документ4 страницыThe Pathophysiology of Malarial Anaemia: Where Have All The Red Cells Gone?rolland_arrizaОценок пока нет
- TLR Agonists and or IL-15 Adjuvanted Mucosal SIV-VaccineДокумент19 страницTLR Agonists and or IL-15 Adjuvanted Mucosal SIV-VaccineYuliLusianaSariОценок пока нет
- 3493 FullДокумент8 страниц3493 Fullgaa5Оценок пока нет
- Acquired Immune Deficiency Syndrome Case Study NotesДокумент2 страницыAcquired Immune Deficiency Syndrome Case Study NotesIan SibalОценок пока нет
- Immune Mechanisms in Type 1 Diabetes: Maja Wa Llberg and Anne CookeДокумент9 страницImmune Mechanisms in Type 1 Diabetes: Maja Wa Llberg and Anne CookeNiko Hizkia SimatupangОценок пока нет
- Modulation of Hos Responses by B-CellsДокумент15 страницModulation of Hos Responses by B-CellsAGLОценок пока нет
- بحث زهراءДокумент25 страницبحث زهراءمحمد عبدالواسع المسنيОценок пока нет
- CD8+ T-Cell Deficiency, Epstein-Barr Virus Infection, Vitamin D Deficiency, and Steps To Autoimmunity - A Unifying HypothesisДокумент16 страницCD8+ T-Cell Deficiency, Epstein-Barr Virus Infection, Vitamin D Deficiency, and Steps To Autoimmunity - A Unifying HypothesiselipoarchОценок пока нет
- (NO) B-Cells-In-The-Tumor-Microenvironment - Multi-FacetДокумент24 страницы(NO) B-Cells-In-The-Tumor-Microenvironment - Multi-FacetPabloОценок пока нет
- Cellular Immunology: Lorena Álvarez-Rodríguez, Marcos López-Hoyos, Pedro Muñoz-Cacho, Víctor Manuel Martínez-TaboadaДокумент9 страницCellular Immunology: Lorena Álvarez-Rodríguez, Marcos López-Hoyos, Pedro Muñoz-Cacho, Víctor Manuel Martínez-TaboadaFernanda Camargo NunesОценок пока нет
- Age and ImmunityДокумент6 страницAge and ImmunityGene Ryuzaki SeseОценок пока нет
- Abstract PDFДокумент36 страницAbstract PDFDhini Kartika SariОценок пока нет
- Tamburini 2012Документ9 страницTamburini 2012AycaОценок пока нет
- Dendritic Cells and Humoral Immunity in Humans: ReviewДокумент5 страницDendritic Cells and Humoral Immunity in Humans: ReviewCarlos GuerreroОценок пока нет
- 2010-Inhibition of B-Cell Activation by Retinal Pigment EpitheliumДокумент6 страниц2010-Inhibition of B-Cell Activation by Retinal Pigment EpitheliumFabricio CardosoОценок пока нет
- PHD Thesis Dendritic CellsДокумент5 страницPHD Thesis Dendritic Cellsashleyjonesmobile100% (1)
- Progression of A Model From The Severe As Well As Postponed Effects of Large Serving Radiation Publicity Within Fitzgibbons Diversity Outbred Rats Comparison For You To Inbred C57BL6 Micedhoci PDFДокумент2 страницыProgression of A Model From The Severe As Well As Postponed Effects of Large Serving Radiation Publicity Within Fitzgibbons Diversity Outbred Rats Comparison For You To Inbred C57BL6 Micedhoci PDFoakpart32Оценок пока нет
- IL-15 Participates in The Respiratory Innate Immune Response To Influenza Virus InfectionДокумент11 страницIL-15 Participates in The Respiratory Innate Immune Response To Influenza Virus InfectionBlack ThokОценок пока нет
- Royal JellyДокумент10 страницRoyal JellyninukОценок пока нет
- Immunology - 2008 - Montcuquet - Regulatory T Cell Expansion and Function Do Not Account For The Impaired Alloreactivity ofДокумент11 страницImmunology - 2008 - Montcuquet - Regulatory T Cell Expansion and Function Do Not Account For The Impaired Alloreactivity ofciara babyОценок пока нет
- 10 1155@2019@2473164Документ13 страниц10 1155@2019@2473164edwinsyahОценок пока нет
- Dendritic Cell Gene TherapyДокумент16 страницDendritic Cell Gene TherapyazharbattooОценок пока нет
- Mike Mast DissertationДокумент10 страницMike Mast DissertationPaperWritingServiceCollegeUK100% (1)
- Alternative Immunomodulatory Strategies For Xenotransplantation: CD80/CD86-CTLA4 Pathway-Modified Immature Dendritic Cells Promote Xenograft SurvivalДокумент7 страницAlternative Immunomodulatory Strategies For Xenotransplantation: CD80/CD86-CTLA4 Pathway-Modified Immature Dendritic Cells Promote Xenograft SurvivalTruc PhamОценок пока нет
- Nov 151Документ6 страницNov 151alexandre06frОценок пока нет
- Targeting BMI-1 in B Cells Restores Effective Humoral Immune Responses and Controls Chronic Viral InfectionДокумент33 страницыTargeting BMI-1 in B Cells Restores Effective Humoral Immune Responses and Controls Chronic Viral InfectionGiorgi BeroshviliОценок пока нет
- Hiv No VelhoДокумент20 страницHiv No VelhoSCIH HFCPОценок пока нет
- Identification and Characterization of Circulating Human Transitional B CellsДокумент9 страницIdentification and Characterization of Circulating Human Transitional B CellsLizeth Rincon DelgadoОценок пока нет
- Modeling Human Adaptive Immune Responses With Tonsil OrganoidsДокумент27 страницModeling Human Adaptive Immune Responses With Tonsil Organoidsissis alppОценок пока нет
- Common Variable Immunodeficiency: Epidemiology, Pathogenesis, Clinical Manifestations, Diagnosis, Classification, and ManagementДокумент21 страницаCommon Variable Immunodeficiency: Epidemiology, Pathogenesis, Clinical Manifestations, Diagnosis, Classification, and ManagementViviana LunaОценок пока нет
- B Cells As A Target of Immune ModulationДокумент6 страницB Cells As A Target of Immune ModulationAlex HydronОценок пока нет
- Effects of HIV-1-induced CD1c and CD1d Modulation and Endogenous Lipid Presentation On CD1c-restricted T-Cell ActivationДокумент10 страницEffects of HIV-1-induced CD1c and CD1d Modulation and Endogenous Lipid Presentation On CD1c-restricted T-Cell ActivationYodha PrasidyaОценок пока нет
- Nat Rev Immunol 2013 TangyeДокумент16 страницNat Rev Immunol 2013 TangyeMarcus GarroneОценок пока нет
- The Effectiveness of The Treatment of Idiopathic Thrombocytopenic Purpura With Modern DrugsДокумент6 страницThe Effectiveness of The Treatment of Idiopathic Thrombocytopenic Purpura With Modern DrugsIJRASETPublicationsОценок пока нет
- T-Cell Clonality in Immune Responses: ViewpointДокумент5 страницT-Cell Clonality in Immune Responses: Viewpointyanisa141Оценок пока нет
- A Follicular Regulatory Innate Lymphoid Cell Population Impairs Interactions Between Germinal Center TFH and B CellsДокумент13 страницA Follicular Regulatory Innate Lymphoid Cell Population Impairs Interactions Between Germinal Center TFH and B Cellsiva hullОценок пока нет
- Final Review Med.J of Hem and Inf DiseasesДокумент14 страницFinal Review Med.J of Hem and Inf Diseasesmgounari7293Оценок пока нет
- General Pathology Bimonthly Exam Compilation Updated 2Документ197 страницGeneral Pathology Bimonthly Exam Compilation Updated 2Cherry Rahima100% (1)
- Perspectives: A Brief History of T Cell Help To B CellsДокумент5 страницPerspectives: A Brief History of T Cell Help To B CellsAstynnia GanОценок пока нет
- JC 13:10:11Документ12 страницJC 13:10:11davdavdavdavdavdavdaОценок пока нет
- Immuno Senescence PublishedДокумент5 страницImmuno Senescence PublishedAcengHamudinОценок пока нет
- MF59 Mediates Its B Cell Adjuvanticity by Promoting T Follicular Helper Cells and Thus Germinal Center Responses in Adult and Early LifeДокумент11 страницMF59 Mediates Its B Cell Adjuvanticity by Promoting T Follicular Helper Cells and Thus Germinal Center Responses in Adult and Early LifeJoão Pedro NunesОценок пока нет
- Nutrients: Physical Activity and Diet Shape The Immune System During AgingДокумент17 страницNutrients: Physical Activity and Diet Shape The Immune System During AgingcarolinaОценок пока нет
- 260Документ8 страниц260Mohammad ArsidОценок пока нет
- Volk Imunitate in SepsisДокумент3 страницыVolk Imunitate in SepsisIna PoianaОценок пока нет
- T Cells in Celiac DiseaseДокумент11 страницT Cells in Celiac DiseaseRadwan AjoОценок пока нет
- 2020 Article 92Документ21 страница2020 Article 92LestiОценок пока нет
- Mol3005 H08Документ13 страницMol3005 H08DenizDerenОценок пока нет
- 07cell Cell InteractionsДокумент9 страниц07cell Cell InteractionsAhmed Goma'aОценок пока нет
- Dendritic Cell-Targeted VaccinesДокумент11 страницDendritic Cell-Targeted VaccinesRui FreitasОценок пока нет
- Soucre 9Документ23 страницыSoucre 9hesssaalsaeediОценок пока нет
- HEP-Adaptive Immune ResponsesДокумент7 страницHEP-Adaptive Immune ResponsesoomculunОценок пока нет
- Annals of The New York Academy of Sciences - 2009 - Bauer - The Role of Stress Factors During Aging of The Immune SystemДокумент14 страницAnnals of The New York Academy of Sciences - 2009 - Bauer - The Role of Stress Factors During Aging of The Immune SystemegrdfgОценок пока нет
- Antibodies Specificities, Isotypes, Receptor-2016Документ32 страницыAntibodies Specificities, Isotypes, Receptor-2016Holder PlaceОценок пока нет
- Dermatomyositis Case7Документ15 страницDermatomyositis Case7Hanina Macataman MetmugОценок пока нет
- CD48Документ10 страницCD48Jaky ShОценок пока нет
- Cerebral Blood FlowДокумент37 страницCerebral Blood FlowFetria MelaniОценок пока нет
- Malnutrition in Stroke PatientДокумент8 страницMalnutrition in Stroke PatientFetria MelaniОценок пока нет
- The Anatomy of A Great Poster: Richard Gerkin, M.D. Bridget Stiegler, D.OДокумент31 страницаThe Anatomy of A Great Poster: Richard Gerkin, M.D. Bridget Stiegler, D.OFetria MelaniОценок пока нет
- Nutrisi Ulseratif KolitisДокумент1 страницаNutrisi Ulseratif KolitisFetria MelaniОценок пока нет
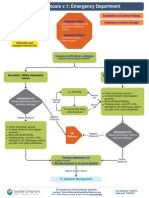
- Algorithm Pyloric StenosisДокумент15 страницAlgorithm Pyloric StenosisFetria MelaniОценок пока нет
- Full Liquid DietДокумент1 страницаFull Liquid DietFetria MelaniОценок пока нет
- Clear Liquid DietДокумент3 страницыClear Liquid DietFetria Melani100% (1)
- Health: Aradhna Kumar Bachelor of Health Science (Podiatry)Документ10 страницHealth: Aradhna Kumar Bachelor of Health Science (Podiatry)Fetria MelaniОценок пока нет
- Metabolisme Zat BesiДокумент33 страницыMetabolisme Zat BesiFetria MelaniОценок пока нет
- Chap 4 Lactation and Human Milk Part 1Документ12 страницChap 4 Lactation and Human Milk Part 1Fetria MelaniОценок пока нет
- Breakdown of Topics - Final Exam GenPath (2016-2017)Документ2 страницыBreakdown of Topics - Final Exam GenPath (2016-2017)Alfred ChowОценок пока нет
- Yogic Cure For ArthritiesДокумент59 страницYogic Cure For Arthritiesstar893190Оценок пока нет
- Seminar On Geriatric Consideration: Presented by Ajay Kumar Patel M.Sc. Nursing 2 SemДокумент68 страницSeminar On Geriatric Consideration: Presented by Ajay Kumar Patel M.Sc. Nursing 2 SemAjayPatelОценок пока нет
- Budwig Cancer GuideДокумент110 страницBudwig Cancer GuideDejan Velkovski100% (2)
- Mechanisms of Bacterial PathogenesisДокумент34 страницыMechanisms of Bacterial PathogenesisMamta ShindeОценок пока нет
- Harris - DFW June 2013Документ172 страницыHarris - DFW June 2013Mahir DemirОценок пока нет
- 24 Lecture PresentationДокумент85 страниц24 Lecture PresentationWide PutinОценок пока нет
- BIO100 Exam 3 Review SheetДокумент8 страницBIO100 Exam 3 Review SheetSeanОценок пока нет
- Acute-Phase Protein - WikipediaДокумент4 страницыAcute-Phase Protein - WikipediaZACHARIAH MANKIRОценок пока нет
- Microbiology: Section IiДокумент40 страницMicrobiology: Section Iiparthibanb88100% (78)
- Chapter19 Transplantation ImmunologyДокумент77 страницChapter19 Transplantation Immunologymalesh123Оценок пока нет
- GXM GSHДокумент1 страницаGXM GSHGerardLum100% (1)
- How To Have Sex in An EpidemicДокумент27 страницHow To Have Sex in An EpidemicchbartsОценок пока нет
- Complement System: by Muhammad Azam Khan GPGC Mandian, AbbottabadДокумент11 страницComplement System: by Muhammad Azam Khan GPGC Mandian, AbbottabadsajidОценок пока нет
- Immunology - 2022 - Chen - New Onset Autoimmune Phenomena Post COVID 19 Vaccination-1Документ17 страницImmunology - 2022 - Chen - New Onset Autoimmune Phenomena Post COVID 19 Vaccination-1Ivan StojanovicОценок пока нет
- Pharmacology ToxicologyДокумент14 страницPharmacology ToxicologyBalwant JassalОценок пока нет
- (2021) - Nutraceuticals Transformation of Conventional Foods Into Health Disease PreventersДокумент28 страниц(2021) - Nutraceuticals Transformation of Conventional Foods Into Health Disease PreventersGuilherme LealОценок пока нет
- MeaslesДокумент23 страницыMeaslesSaad MotawéaОценок пока нет
- What Are The HLA Genes?: Humans Have Three Main MHC Class I Genes, Known As HLA-A, HLA-B, and HLA-C. TheДокумент2 страницыWhat Are The HLA Genes?: Humans Have Three Main MHC Class I Genes, Known As HLA-A, HLA-B, and HLA-C. ThepoojaОценок пока нет
- Covid 19Документ164 страницыCovid 19Dùtôivẫnnởnụcười Nhưngcuộcsốngcủatôi ChưabaogiờlàổnОценок пока нет
- Childrens FBC Reference Ranges PDFДокумент1 страницаChildrens FBC Reference Ranges PDFWarisatul Imam LubisОценок пока нет
- Ilovepdf Merged 2 PDFДокумент307 страницIlovepdf Merged 2 PDFAhmed ZidanОценок пока нет
- Christine Schaffner 2020 Essential Brain HygieneДокумент20 страницChristine Schaffner 2020 Essential Brain HygienereyshmeОценок пока нет
- Covid-19 Igm/Igg Rapid Test: Easy, Efficient, and Accurate Screening Device For Covid-19Документ3 страницыCovid-19 Igm/Igg Rapid Test: Easy, Efficient, and Accurate Screening Device For Covid-19German IzurietaОценок пока нет
- Immunomodulatory Activity of BhallatakДокумент35 страницImmunomodulatory Activity of BhallatakBalaji Kumar PanigrahiОценок пока нет
- King of Bitters (Andrographis Paniculata)Документ3 страницыKing of Bitters (Andrographis Paniculata)mateto_2014100% (1)
- Unit 3 Part 1 Introduction To Autoimmune DisordersДокумент14 страницUnit 3 Part 1 Introduction To Autoimmune DisordersReman AlingasaОценок пока нет
- Li2018 PDFДокумент9 страницLi2018 PDFRahmad SyamsulОценок пока нет
- Lymphatic System DiseasesДокумент1 страницаLymphatic System DiseasesRJ San JuanОценок пока нет
- Adaptogens: New Age Healing Gems For Physical Wellbeing: Rehaan Sharma, Pawan Sharma, Renu BhardwajДокумент10 страницAdaptogens: New Age Healing Gems For Physical Wellbeing: Rehaan Sharma, Pawan Sharma, Renu BhardwajajmrdОценок пока нет