Вам также может понравиться
- Hey Friends B TBДокумент152 страницыHey Friends B TBTizianoCiro CarrizoОценок пока нет
- GT-N7100-Full Schematic PDFДокумент67 страницGT-N7100-Full Schematic PDFprncha86% (7)
- DC 7 BrochureДокумент4 страницыDC 7 Brochures_a_r_r_yОценок пока нет
- 1983 - A Method For The Isolation of Longevity Mutants in The Nematode Caenorhabditis Elegans and Initial ResultsДокумент8 страниц1983 - A Method For The Isolation of Longevity Mutants in The Nematode Caenorhabditis Elegans and Initial ResultsRaymond LaBoyОценок пока нет
- The Dinoflagellate Dinophysis Norvegica Biological and Ecological Observations in The Baltic SeaДокумент10 страницThe Dinoflagellate Dinophysis Norvegica Biological and Ecological Observations in The Baltic SeaHabib Ben HabibОценок пока нет
- Microbiology EcologyДокумент13 страницMicrobiology Ecologynhanhnafi5Оценок пока нет
- The Survival of Infectious Bronchitis Ib Virus in WaterДокумент12 страницThe Survival of Infectious Bronchitis Ib Virus in WaterGanjar AlaydrussaniОценок пока нет
- Respiratory Rates of Migrating Zooplankton in The Natural, Habitat John W. Bishop?Документ5 страницRespiratory Rates of Migrating Zooplankton in The Natural, Habitat John W. Bishop?toboОценок пока нет
- Tmp541e TMPДокумент3 страницыTmp541e TMPFrontiersОценок пока нет
- Viability of EscherichiaДокумент10 страницViability of EscherichiagiuseppegnrОценок пока нет
- Desert Snails: Problems of Heat, Water and FoodДокумент14 страницDesert Snails: Problems of Heat, Water and FoodcoordinatedmultibeamsatelliteОценок пока нет
- Physiological Studies of Subtropical Mangrove ThraustochytridsДокумент8 страницPhysiological Studies of Subtropical Mangrove ThraustochytridsIsnaini PrihatiningsihОценок пока нет
- Deteccion of Helicobacter y Campilobacter en Mamiferos MarinosДокумент5 страницDeteccion of Helicobacter y Campilobacter en Mamiferos MarinosCamilo Huertas CamposОценок пока нет
- Tangaroa1109 PDFДокумент33 страницыTangaroa1109 PDFS.paniruban S.panirubanОценок пока нет
- González Et Al., 2002Документ11 страницGonzález Et Al., 2002cindy tatiana paba carreñoОценок пока нет
- Escherichia Coli and Total Coliforms in Water and Sediments at Lake MarinasДокумент8 страницEscherichia Coli and Total Coliforms in Water and Sediments at Lake MarinasAsfandОценок пока нет
- The Effects of Season, Temperature and Salinity On The Oxygen Consumption of Bivalve Gill TissueДокумент12 страницThe Effects of Season, Temperature and Salinity On The Oxygen Consumption of Bivalve Gill TissueineeddocОценок пока нет
- Legionella: Environmental Factors Affecting The Colonization of Cool-Ing Towers by Spp. in South AustraliaДокумент9 страницLegionella: Environmental Factors Affecting The Colonization of Cool-Ing Towers by Spp. in South AustraliaAliAliОценок пока нет
- The Accumulation of Low Molecular Weight Aromatic Hydrocarbons of Crude Off by Coho SalmonДокумент8 страницThe Accumulation of Low Molecular Weight Aromatic Hydrocarbons of Crude Off by Coho SalmonSaeed DelaramОценок пока нет
- Hershey Chase 1952Документ18 страницHershey Chase 1952moritheilОценок пока нет
- The Fate of 2-, 3-, and 4-Chlorobiphenyl Following Intravenous Administration To The Thorny Skate and The Winter SkateДокумент10 страницThe Fate of 2-, 3-, and 4-Chlorobiphenyl Following Intravenous Administration To The Thorny Skate and The Winter SkateSh1vaОценок пока нет
- Cryptosporidium Spp. and Giardia Duodenalis IsolatesДокумент8 страницCryptosporidium Spp. and Giardia Duodenalis IsolateswiwienОценок пока нет
- Warren, Benoit and Jessee, 1978Документ6 страницWarren, Benoit and Jessee, 1978Sajad MoradiОценок пока нет
- Lsochrysis Galbana: Optimal Growth Conditions ForДокумент12 страницLsochrysis Galbana: Optimal Growth Conditions ForHuy Hữu NguyễnОценок пока нет
- Kangatharalingam1993 PDFДокумент4 страницыKangatharalingam1993 PDFSmokeОценок пока нет
- Algae Nitrogen FixationДокумент10 страницAlgae Nitrogen FixationAshish KaushalОценок пока нет
- TMP EEF6Документ6 страницTMP EEF6FrontiersОценок пока нет
- Bio Unit Exam 2Документ12 страницBio Unit Exam 2api-216550327Оценок пока нет
- 1997-Riquemel Et AlДокумент5 страниц1997-Riquemel Et AlYendry VargasОценок пока нет
- BF00248485Документ7 страницBF00248485ahmed shawkyОценок пока нет
- In Situ Measurement of The Settling Velocity of Organic Carbon Particles and Species of PhytoplanktonДокумент10 страницIn Situ Measurement of The Settling Velocity of Organic Carbon Particles and Species of PhytoplanktonAnonymous d6vkxJОценок пока нет
- Viruses From Sulfolobus and Close Relatives Dpb5imДокумент8 страницViruses From Sulfolobus and Close Relatives Dpb5imywang9628Оценок пока нет
- Determination of PCB Congener-Specific First Order Absorption/Desorption Rate Constants Using Larvae (Insecta: Diptera: Chironomidae)Документ7 страницDetermination of PCB Congener-Specific First Order Absorption/Desorption Rate Constants Using Larvae (Insecta: Diptera: Chironomidae)Ufuoma AsagbaОценок пока нет
- Test UploadДокумент8 страницTest UploadAbi Diciembre BrionesОценок пока нет
- Left and Right Panel FinalДокумент2 страницыLeft and Right Panel Finalapi-281974254Оценок пока нет
- Effects of Acute Temperature Change On The Metabolism and Swimming Ability of SterletДокумент5 страницEffects of Acute Temperature Change On The Metabolism and Swimming Ability of SterletRaluca-Cristina GuriencuОценок пока нет
- The Effects of Zinc On The Mortality and Reproduction of The MinnowДокумент14 страницThe Effects of Zinc On The Mortality and Reproduction of The MinnowSh1vaОценок пока нет
- Su-Jung Chang, Su-Min Tseng, and Hsin-Yiu ChouДокумент9 страницSu-Jung Chang, Su-Min Tseng, and Hsin-Yiu ChouMirza ČelebičićОценок пока нет
- Kerb Rat 2010Документ7 страницKerb Rat 2010Roriscorpion RosydaОценок пока нет
- (Received April 1988 Revision 26 Sept. 1988) : Minerals Engineering, Vol. 2, No. 2, Pp. 217-227, 1989Документ11 страниц(Received April 1988 Revision 26 Sept. 1988) : Minerals Engineering, Vol. 2, No. 2, Pp. 217-227, 1989alinoriОценок пока нет
- J. Lipid Res.-2002-Damst・1641-51Документ11 страницJ. Lipid Res.-2002-Damst・1641-51ymiyazyОценок пока нет
- Sublethal Stress in Escherichia Coli A Function of SalinitytДокумент6 страницSublethal Stress in Escherichia Coli A Function of SalinitytgiuseppegnrОценок пока нет
- Journal Review: Utilization of Halogen Clor Elements in The Use of Chlorine Dioxide in Fruits and Vegetables For Salmonella (Biochemistry)Документ7 страницJournal Review: Utilization of Halogen Clor Elements in The Use of Chlorine Dioxide in Fruits and Vegetables For Salmonella (Biochemistry)DesiratnasariОценок пока нет
- Clostridium Hungatei Sp. Nov., A Mesophilic, N: - Fixing Cellulolytic Bacterium Isolated From SoilДокумент10 страницClostridium Hungatei Sp. Nov., A Mesophilic, N: - Fixing Cellulolytic Bacterium Isolated From SoilRajapriya Prabha KaranОценок пока нет
- Chromium-Induced Micronuclei in Fish: K. Al-Sabti,? M. Franko, B. Andrijanie, S. Knez and P. StegnarДокумент4 страницыChromium-Induced Micronuclei in Fish: K. Al-Sabti,? M. Franko, B. Andrijanie, S. Knez and P. StegnarAyelen Gonzalez NuñezОценок пока нет
- The Effect of Temperature On The Hatching Success of Brine ShrimpsДокумент15 страницThe Effect of Temperature On The Hatching Success of Brine ShrimpsTootsie82% (11)
- Efectos Del CO 2 Elevado en La Reproducción de Dos Copépodos CalanoideosДокумент7 страницEfectos Del CO 2 Elevado en La Reproducción de Dos Copépodos CalanoideosISABEL SANCHEZ RIVERAОценок пока нет
- tmp2F18 TMPДокумент13 страницtmp2F18 TMPFrontiersОценок пока нет
- Holm 1986 Aquacultural-EngineeringДокумент15 страницHolm 1986 Aquacultural-EngineeringJorge RodriguezОценок пока нет
- Effects of Diet On Population Development of The Rotifer Brachion Us Plica Tiffs in CultureДокумент11 страницEffects of Diet On Population Development of The Rotifer Brachion Us Plica Tiffs in CultureEnrique MartinezОценок пока нет
- Influence of The Dipolar Magnetized Water On The Ecological Factors of Freshwater Ostracod Cypris Laevis (O. F. Müller, 1776)Документ7 страницInfluence of The Dipolar Magnetized Water On The Ecological Factors of Freshwater Ostracod Cypris Laevis (O. F. Müller, 1776)IOSRjournalОценок пока нет
- E ColiДокумент20 страницE ColiArabic TalentОценок пока нет
- Longzcornzs On An Algal Mat: Ecol.Документ9 страницLongzcornzs On An Algal Mat: Ecol.RicardoDennisMezaNiflaОценок пока нет
- Bacterial Transformation: The Lab Protocol Can Be Conceptualized As Four Major Steps. Pre-IncubationДокумент4 страницыBacterial Transformation: The Lab Protocol Can Be Conceptualized As Four Major Steps. Pre-IncubationGUIDO ERNESTO VILLOTA CALVACHIОценок пока нет
- Notes: Notes On Food, Beverages and Fomites Contaminated With Vibrio CholeraeДокумент10 страницNotes: Notes On Food, Beverages and Fomites Contaminated With Vibrio CholeraeMoonsoul ChildОценок пока нет
- KARI, Acta Chem. Scand., OSTEOX, Biochim. Biophys. Acta, Schryver, MUKHERJEE, Proc. Roy. Soc. (London)Документ9 страницKARI, Acta Chem. Scand., OSTEOX, Biochim. Biophys. Acta, Schryver, MUKHERJEE, Proc. Roy. Soc. (London)Rosa Elsy Puentes LondoñoОценок пока нет
- Assessment of Cryptophyceae Ingestion by Copepods Using Alloxanthin Pigment A CautionДокумент77 страницAssessment of Cryptophyceae Ingestion by Copepods Using Alloxanthin Pigment A CautionSamОценок пока нет
- Lahnsteiner 2012Документ10 страницLahnsteiner 2012Belay AbdissaОценок пока нет
- Effects of Bicarbonate and Carbon-Dioxide On The Competition-Between Chlorella and SpirulinaДокумент8 страницEffects of Bicarbonate and Carbon-Dioxide On The Competition-Between Chlorella and SpirulinaHitesh JethaniОценок пока нет
- The Effect of Temperature and Body Size On Filtration Rates of Limnoperna Fortunei (Bivalvia, Mytilidae) Under Laboratory ConditionsДокумент10 страницThe Effect of Temperature and Body Size On Filtration Rates of Limnoperna Fortunei (Bivalvia, Mytilidae) Under Laboratory ConditionsCarlos MeirellesОценок пока нет
- Nguyen T Final Research PaperДокумент20 страницNguyen T Final Research PaperTuong NguyenОценок пока нет
- Bacteriological Quality of The Sand Beach in Sopot (Gdansk Bay, Southern Baltic)Документ5 страницBacteriological Quality of The Sand Beach in Sopot (Gdansk Bay, Southern Baltic)Nazzalia Isabel Rodriguez CastroОценок пока нет
- Physiology and Behaviour of Marine Organisms: Proceedings of the 12th European Symposium on Marine Biology, Stirling, Scotland, September 1977От EverandPhysiology and Behaviour of Marine Organisms: Proceedings of the 12th European Symposium on Marine Biology, Stirling, Scotland, September 1977D.S. McLuskyОценок пока нет
- Detection and Enumeration of Salmonella and Pseudomonas AeruginosaДокумент10 страницDetection and Enumeration of Salmonella and Pseudomonas AeruginosagiuseppegnrОценок пока нет
- Impedance Microbiology-A Rapid Change For MicrobiologistsДокумент11 страницImpedance Microbiology-A Rapid Change For Microbiologistsgiuseppegnr100% (1)
- Incidence and Role of Salmonella in Seafood SafetyДокумент9 страницIncidence and Role of Salmonella in Seafood SafetygiuseppegnrОценок пока нет
- Selective Enumeration of LactobacillusДокумент4 страницыSelective Enumeration of LactobacillusgiuseppegnrОценок пока нет
- A Medium For The Cultivation of LactobacilliДокумент6 страницA Medium For The Cultivation of LactobacilligiuseppegnrОценок пока нет
- Sublethal Stress in Escherichia Coli A Function of SalinitytДокумент6 страницSublethal Stress in Escherichia Coli A Function of SalinitytgiuseppegnrОценок пока нет
- Marine Bacteria Which Produce TetrodotoxinДокумент2 страницыMarine Bacteria Which Produce TetrodotoxingiuseppegnrОценок пока нет
- Serial Dilution Simulation LabДокумент3 страницыSerial Dilution Simulation LabgiuseppegnrОценок пока нет
- Viability of EscherichiaДокумент10 страницViability of EscherichiagiuseppegnrОценок пока нет
- Introduction-: Microprocessor 68000Документ13 страницIntroduction-: Microprocessor 68000margyaОценок пока нет
- How Drugs Work - Basic Pharmacology For Healthcare ProfessionalsДокумент19 страницHow Drugs Work - Basic Pharmacology For Healthcare ProfessionalsSebastián Pérez GuerraОценок пока нет
- Global Geo Reviewer MidtermДокумент29 страницGlobal Geo Reviewer Midtermbusinesslangto5Оценок пока нет
- Arnold Ventures Letter To Congressional Social Determinants of Health CaucusДокумент7 страницArnold Ventures Letter To Congressional Social Determinants of Health CaucusArnold VenturesОценок пока нет
- Test Your Knowledge - Study Session 1Документ4 страницыTest Your Knowledge - Study Session 1My KhanhОценок пока нет
- The Mantel Colonized Nation Somalia 10 PDFДокумент5 страницThe Mantel Colonized Nation Somalia 10 PDFAhmad AbrahamОценок пока нет
- 15.053/8 February 7, 2013: More Linear and Non-Linear Programming ModelsДокумент42 страницы15.053/8 February 7, 2013: More Linear and Non-Linear Programming ModelsShashank SinglaОценок пока нет
- MASONRYДокумент8 страницMASONRYJowelyn MaderalОценок пока нет
- Business Analytics Emphasis Course GuideДокумент3 страницыBusiness Analytics Emphasis Course Guidea30000496Оценок пока нет
- Environmental Economics Pollution Control: Mrinal Kanti DuttaДокумент253 страницыEnvironmental Economics Pollution Control: Mrinal Kanti DuttashubhamОценок пока нет
- Deal Report Feb 14 - Apr 14Документ26 страницDeal Report Feb 14 - Apr 14BonviОценок пока нет
- Snapdragon 435 Processor Product Brief PDFДокумент2 страницыSnapdragon 435 Processor Product Brief PDFrichardtao89Оценок пока нет
- Waterstop TechnologyДокумент69 страницWaterstop TechnologygertjaniОценок пока нет
- Oracle - Prep4sure.1z0 068.v2016!07!12.by - Lana.60qДокумент49 страницOracle - Prep4sure.1z0 068.v2016!07!12.by - Lana.60qLuis AlfredoОценок пока нет
- CT SizingДокумент62 страницыCT SizingMohamed TalebОценок пока нет
- 2009 2011 DS Manual - Club Car (001-061)Документ61 страница2009 2011 DS Manual - Club Car (001-061)misaОценок пока нет
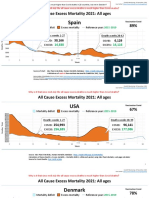
- Countries EXCESS DEATHS All Ages - 15nov2021Документ21 страницаCountries EXCESS DEATHS All Ages - 15nov2021robaksОценок пока нет
- Assessment of Students' Oral Communication in English ClassДокумент10 страницAssessment of Students' Oral Communication in English ClassKeebeek S ArbasОценок пока нет
- Report Card Grade 1 2Документ3 страницыReport Card Grade 1 2Mely DelacruzОценок пока нет
- Developing Paragraphs: Test Section - Writing Task 2Документ11 страницDeveloping Paragraphs: Test Section - Writing Task 2Cengizhan AkdağОценок пока нет
- NDY 9332v3Документ8 страницNDY 9332v3sulphurdioxideОценок пока нет
- Biological Beneficiation of Kaolin: A Review On Iron RemovalДокумент8 страницBiological Beneficiation of Kaolin: A Review On Iron RemovalValentin GnoumouОценок пока нет
- The Magic DrumДокумент185 страницThe Magic Drumtanishgiri2012Оценок пока нет
- Opc PPT FinalДокумент22 страницыOpc PPT FinalnischalaОценок пока нет
- Roleplayer: The Accused Enchanted ItemsДокумент68 страницRoleplayer: The Accused Enchanted ItemsBarbie Turic100% (1)
- Outdoor Air Pollution: Sources, Health Effects and SolutionsДокумент20 страницOutdoor Air Pollution: Sources, Health Effects and SolutionsCamelia RadulescuОценок пока нет
- Elements of ArtДокумент1 страницаElements of Artsamson8cindy8louОценок пока нет