Вам также может понравиться
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (895)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- Azokeratin ProtocolДокумент8 страницAzokeratin ProtocolconcatamerОценок пока нет
- Chryseobacterium Library Construction: Restriction Endonuclease RangeДокумент2 страницыChryseobacterium Library Construction: Restriction Endonuclease RangeconcatamerОценок пока нет
- ASP13 (1) 2014 03-18 MaxДокумент16 страницASP13 (1) 2014 03-18 MaxconcatamerОценок пока нет
- A Rapid Aflatoxin B ELISA: Development and Validation With Reduced Matrix Effects For Peanuts, Corn, Pistachio, and SoybeansДокумент10 страницA Rapid Aflatoxin B ELISA: Development and Validation With Reduced Matrix Effects For Peanuts, Corn, Pistachio, and SoybeansconcatamerОценок пока нет
- Choudhary KeraДокумент8 страницChoudhary KeraconcatamerОценок пока нет
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (400)
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (345)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2259)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (121)
- Obesity - The Health Time Bomb: ©LTPHN 2008Документ36 страницObesity - The Health Time Bomb: ©LTPHN 2008EVA PUTRANTO100% (2)
- All You Need To Know About Egg YolkДокумент7 страницAll You Need To Know About Egg YolkGolden Era BookwormОценок пока нет
- Chemistry Form 4 Daily Lesson Plan - CompressДокумент3 страницыChemistry Form 4 Daily Lesson Plan - Compressadila ramlonОценок пока нет
- Read While Being Blind.. Braille's Alphabet: Be Aware and Active !Документ3 страницыRead While Being Blind.. Braille's Alphabet: Be Aware and Active !bitermanОценок пока нет
- LM74680 Fasson® Fastrans NG Synthetic PE (ST) / S-2050/ CK40Документ2 страницыLM74680 Fasson® Fastrans NG Synthetic PE (ST) / S-2050/ CK40Nishant JhaОценок пока нет
- Mcdonald 2016Документ10 страницMcdonald 2016Andrika SaputraОценок пока нет
- Marketing FinalДокумент15 страницMarketing FinalveronicaОценок пока нет
- Chapter 13 Exercises With AnswerДокумент5 страницChapter 13 Exercises With AnswerTabitha HowardОценок пока нет
- Gods Omnipresence in The World On Possible MeaninДокумент20 страницGods Omnipresence in The World On Possible MeaninJoan Amanci Casas MuñozОценок пока нет
- 3 ALCE Insulators 12R03.1Документ12 страниц3 ALCE Insulators 12R03.1Amílcar Duarte100% (1)
- Presentation 11Документ14 страницPresentation 11stellabrown535Оценок пока нет
- Principals' Leadership Styles and Student Academic Performance in Secondary Schools in Ekiti State, NigeriaДокумент12 страницPrincipals' Leadership Styles and Student Academic Performance in Secondary Schools in Ekiti State, NigeriaiqraОценок пока нет
- Nama: Yossi Tiara Pratiwi Kelas: X Mis 1 Mata Pelajaran: Bahasa InggrisДокумент2 страницыNama: Yossi Tiara Pratiwi Kelas: X Mis 1 Mata Pelajaran: Bahasa InggrisOrionj jrОценок пока нет
- Problem Set-02Документ2 страницыProblem Set-02linn.pa.pa.khaing.2020.2021.fbОценок пока нет
- Parliament of India: Rajya SabhaДокумент64 страницыParliament of India: Rajya SabhaSivapothuraju KonathalaОценок пока нет
- Quick Help For EDI SEZ IntegrationДокумент2 страницыQuick Help For EDI SEZ IntegrationsrinivasОценок пока нет
- PED003Документ1 страницаPED003ely mae dag-umanОценок пока нет
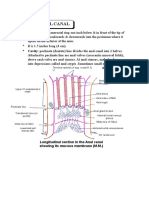
- Anatomy Anal CanalДокумент14 страницAnatomy Anal CanalBela Ronaldoe100% (1)
- The Person Environment Occupation (PEO) Model of Occupational TherapyДокумент15 страницThe Person Environment Occupation (PEO) Model of Occupational TherapyAlice GiffordОценок пока нет
- Analysis of Electric Machinery Krause Manual Solution PDFДокумент2 страницыAnalysis of Electric Machinery Krause Manual Solution PDFKuldeep25% (8)
- School of Mathematics 2021 Semester 1 MAT1841 Continuous Mathematics For Computer Science Assignment 1Документ2 страницыSchool of Mathematics 2021 Semester 1 MAT1841 Continuous Mathematics For Computer Science Assignment 1STEM Education Vung TauОценок пока нет
- KP Tevta Advertisement 16-09-2019Документ4 страницыKP Tevta Advertisement 16-09-2019Ishaq AminОценок пока нет
- Origami Oso HormigueroДокумент9 страницOrigami Oso HormigueroRogelio CerdaОценок пока нет
- Sindi and Wahab in 18th CenturyДокумент9 страницSindi and Wahab in 18th CenturyMujahid Asaadullah AbdullahОценок пока нет
- Ob NotesДокумент8 страницOb NotesRahul RajputОценок пока нет
- Comparitive Study ICICI & HDFCДокумент22 страницыComparitive Study ICICI & HDFCshah faisal100% (1)
- Ricoh IM C2000 IM C2500: Full Colour Multi Function PrinterДокумент4 страницыRicoh IM C2000 IM C2500: Full Colour Multi Function PrinterKothapalli ChiranjeeviОценок пока нет
- Lec 33 - Householder MethodДокумент11 страницLec 33 - Householder MethodMudit SinhaОценок пока нет
- BNF Pos - StockmockДокумент14 страницBNF Pos - StockmockSatish KumarОценок пока нет
- Review1 ScheduleДокумент3 страницыReview1 Schedulejayasuryam.ae18Оценок пока нет