
Вам также может понравиться
- Structural Organisation in AnimalsДокумент49 страницStructural Organisation in AnimalsPukazhvanthen Paramanandhan100% (1)
- BioSci 101 Test 2 NotesДокумент12 страницBioSci 101 Test 2 NotesPianomanSuperman100% (1)
- Mitosis QuestionsДокумент10 страницMitosis QuestionscapremaeОценок пока нет
- Species Concept, Speciation Evolutionary Thought Before DarwinДокумент26 страницSpecies Concept, Speciation Evolutionary Thought Before DarwinmashedtomatoezedОценок пока нет
- Is This The Man Behind The Global Coronavirus PandemicДокумент6 страницIs This The Man Behind The Global Coronavirus PandemicMonte CarloОценок пока нет
- Topic 1 Cellular Respiration FS2014 VO Part 2Документ29 страницTopic 1 Cellular Respiration FS2014 VO Part 2宛仪Оценок пока нет
- 5905 Et 21-Biosystematics-EtДокумент10 страниц5905 Et 21-Biosystematics-EtAnjali Singh100% (1)
- Principles of Systematic Zoology TaxonomyДокумент19 страницPrinciples of Systematic Zoology TaxonomyKhadija IkramОценок пока нет
- 6.LECTURE 6 - TaxonomyДокумент35 страниц6.LECTURE 6 - TaxonomyMuhammad Danial HasanОценок пока нет
- Edexcel GCE Biology Unit 2 Exam Revision NotesДокумент18 страницEdexcel GCE Biology Unit 2 Exam Revision NotescolachauОценок пока нет
- Engler and Prantel System of ClassificationДокумент14 страницEngler and Prantel System of ClassificationEasha tu RaziaОценок пока нет
- PoriferaДокумент39 страницPoriferaabhishek.ajmaniОценок пока нет
- 1 - Principles of ClassificationДокумент42 страницы1 - Principles of ClassificationSandhya LakshmiОценок пока нет
- Lab Practical 1Документ21 страницаLab Practical 1randy_sanchez_10Оценок пока нет
- Zoology 2008Документ91 страницаZoology 2008Ajmal Khan0% (2)
- Porifera - EvolucionДокумент9 страницPorifera - EvolucionJuan David HPОценок пока нет
- Bio Zoology 11thДокумент247 страницBio Zoology 11thPradeep ChandarОценок пока нет
- Animals ClassificationДокумент10 страницAnimals ClassificationVivek JainОценок пока нет
- Lab+3+Taxonomy+Lab ACДокумент42 страницыLab+3+Taxonomy+Lab ACJustinSantostefanoОценок пока нет
- Assignment 1 Classification, Ecology and SoilДокумент2 страницыAssignment 1 Classification, Ecology and SoilLee-Anne LutchmanОценок пока нет
- Micro TaxonomyДокумент29 страницMicro TaxonomyGujulla RahulОценок пока нет
- Lab 10 Porifera & CnidariaДокумент25 страницLab 10 Porifera & Cnidariadraykid100% (1)
- Chapter 1 Classification SystemsДокумент18 страницChapter 1 Classification SystemsAsadОценок пока нет
- Systematic Approaches To Phylogeny)Документ26 страницSystematic Approaches To Phylogeny)Mhi IsmailОценок пока нет
- Kingdom Protista: Submitted ToДокумент57 страницKingdom Protista: Submitted ToAlleli AspiliОценок пока нет
- Biological Classification 2013Документ23 страницыBiological Classification 2013Anonymous xU5C5lОценок пока нет
- Clades - Research QuestionsДокумент3 страницыClades - Research QuestionsK KorukxuОценок пока нет
- Genetic TechnologyДокумент36 страницGenetic TechnologyAnggraeni Kusuma WardaniОценок пока нет
- Theories of Organic EvolutionДокумент15 страницTheories of Organic EvolutionAdityaОценок пока нет
- Membrane Models: Gorter and Grendel's Membrane Theory (1920)Документ5 страницMembrane Models: Gorter and Grendel's Membrane Theory (1920)Hafiz AhmadОценок пока нет
- Pieris Homo Lathyrus Sapiens OdoratusДокумент1 страницаPieris Homo Lathyrus Sapiens OdoratusjennieОценок пока нет
- Zoology Notes PDFДокумент6 страницZoology Notes PDFAdarsh jainОценок пока нет
- Gen Bio Mod 6Документ24 страницыGen Bio Mod 6GMusa, Dexie Heart U.Оценок пока нет
- Cell DifferentiationДокумент16 страницCell DifferentiationHarika KeshavОценок пока нет
- Zoology NotesДокумент42 страницыZoology NotesNolour de los SantosОценок пока нет
- Chapter 4 Animal KingdomДокумент16 страницChapter 4 Animal KingdomYashiОценок пока нет
- Classification, Morphology, Life Cycle of ProtozoaДокумент15 страницClassification, Morphology, Life Cycle of ProtozoaFaris Muhammad100% (1)
- Heredity and VariationДокумент25 страницHeredity and VariationDre ParkerОценок пока нет
- Replication in EukaryotesДокумент8 страницReplication in EukaryotesnaliniОценок пока нет
- Introduction To ZoologyДокумент51 страницаIntroduction To Zoologyirish x100% (1)
- 01 The Living WorldДокумент5 страниц01 The Living WorldHerick RobinОценок пока нет
- +1 Zoology Focus Area NotesДокумент85 страниц+1 Zoology Focus Area NotesKhatarnakОценок пока нет
- 120-11th Bio Zoology - Unit 1-6 Important Questions - English Medium PDF DownloadДокумент6 страниц120-11th Bio Zoology - Unit 1-6 Important Questions - English Medium PDF Downloadtamilan storesОценок пока нет
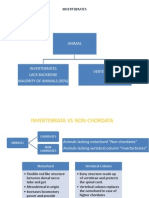
- ZOOLOGY (First PAPER) (Structure and Functions of Invertebrate Types)Документ2 страницыZOOLOGY (First PAPER) (Structure and Functions of Invertebrate Types)GuruKPO0% (1)
- Cytoskeleton DAДокумент35 страницCytoskeleton DAAnonymous NVBCqKWHОценок пока нет
- Species InteractionДокумент19 страницSpecies InteractionStyrich Nyl AbayonОценок пока нет
- Phylum PoriferaДокумент19 страницPhylum PoriferaRen Zo ImpelidoОценок пока нет
- Entomology Lab HandoutsДокумент13 страницEntomology Lab Handoutshumanupgrade100% (1)
- Class: 8 Science Chapter - 8 Cell - Structure and FunctionsДокумент10 страницClass: 8 Science Chapter - 8 Cell - Structure and FunctionsAmiteshwar SinghОценок пока нет
- Cell - The Unit of LifeДокумент71 страницаCell - The Unit of LifeGanesh PatilОценок пока нет
- NomenclatureДокумент54 страницыNomenclatureDestha RamadantyОценок пока нет
- Zoology 100 Notes 1Документ8 страницZoology 100 Notes 1Bethany Jane Ravelo IsidroОценок пока нет
- Zoology Notes: 009 Chapter 5Документ4 страницыZoology Notes: 009 Chapter 5humanupgrade100% (1)
- Animal KingdomДокумент22 страницыAnimal KingdomdellfrogОценок пока нет
- Chapter-2 - Biological Classification NotesДокумент7 страницChapter-2 - Biological Classification NotesAnita TandonОценок пока нет
- The Pyridine Nucleotide CoenzymesОт EverandThe Pyridine Nucleotide CoenzymesJohannes EverseОценок пока нет
- Purpose of Taxonomy in BiologyДокумент7 страницPurpose of Taxonomy in BiologytboylingweОценок пока нет
- Taxonomy PDFДокумент15 страницTaxonomy PDFSwarup MondalОценок пока нет
- PDF Document 4Документ9 страницPDF Document 4Sheena PajoОценок пока нет
- Plant TaxonomyДокумент7 страницPlant TaxonomySiaОценок пока нет
- European Journal of BiomedicalДокумент7 страницEuropean Journal of BiomedicalDr Swapna GurrapuОценок пока нет
- A novel library of -arylketones as potential inhibitors of α-glucosidase: Their design, synthesis, in vitro and in vivo studiesДокумент13 страницA novel library of -arylketones as potential inhibitors of α-glucosidase: Their design, synthesis, in vitro and in vivo studiesEstari MamidalaОценок пока нет
- In Vitro Anti Bacterial Activity of Phytosterol Isolated From Aerva Lanata RootsДокумент5 страницIn Vitro Anti Bacterial Activity of Phytosterol Isolated From Aerva Lanata RootsDr Swapna GurrapuОценок пока нет
- A novel library of -arylketones as potential inhibitors of α-glucosidase: Their design, synthesis, in vitro and in vivo studiesДокумент13 страницA novel library of -arylketones as potential inhibitors of α-glucosidase: Their design, synthesis, in vitro and in vivo studiesEstari MamidalaОценок пока нет
- Life SciencesДокумент6 страницLife SciencesDr Swapna GurrapuОценок пока нет
- 2018 Satish PoojariДокумент4 страницы2018 Satish PoojariDr Estari MamidalaОценок пока нет
- International Journal of Pharma Research and Health Sciences PDFДокумент5 страницInternational Journal of Pharma Research and Health Sciences PDFDr Swapna GurrapuОценок пока нет
- Hiv-1 Reverse Transcriptase Inhibition by Phenolic Compounds Isolated From Acalypha Indica (L.) Plant Leaves ExtractДокумент5 страницHiv-1 Reverse Transcriptase Inhibition by Phenolic Compounds Isolated From Acalypha Indica (L.) Plant Leaves ExtractDr Swapna GurrapuОценок пока нет
- Synthesis, anticancer and antibacterial evaluation of novel (isopropylidene) uridine-[1,2,3]triazole hybrids Ranjith kumar Thatipamula a, Sirassu Narsimha a, Kumaraswamy Battula a, V. Rajendra Chary b, Estari Mamidala b, Nagavelli Vasudeva ReddyДокумент8 страницSynthesis, anticancer and antibacterial evaluation of novel (isopropylidene) uridine-[1,2,3]triazole hybrids Ranjith kumar Thatipamula a, Sirassu Narsimha a, Kumaraswamy Battula a, V. Rajendra Chary b, Estari Mamidala b, Nagavelli Vasudeva ReddyEstari Mamidala100% (1)
- IN VITRO ANTIBACTERIAL ACTIVITY OF PHYSAGULIN ISOLATED FROM FRUITS OF PHYSALIS ANGULATA AGAINST PATHOGENIC CLINICALLY IMPORTANT BACTERIA by Swapna Gurrapu and Estari MamidalaДокумент5 страницIN VITRO ANTIBACTERIAL ACTIVITY OF PHYSAGULIN ISOLATED FROM FRUITS OF PHYSALIS ANGULATA AGAINST PATHOGENIC CLINICALLY IMPORTANT BACTERIA by Swapna Gurrapu and Estari MamidalaEstari MamidalaОценок пока нет
- Publish Paper E.AДокумент5 страницPublish Paper E.ADr Swapna GurrapuОценок пока нет
- SJLS 31 1 4 PDFДокумент4 страницыSJLS 31 1 4 PDFDr Swapna GurrapuОценок пока нет
- In Vitro Hiv-1 Reverse Transcriptase Inhibition by Alkaloids Isolated From Leaves of Eclipta AlbaДокумент5 страницIn Vitro Hiv-1 Reverse Transcriptase Inhibition by Alkaloids Isolated From Leaves of Eclipta AlbaDr Swapna GurrapuОценок пока нет
- Spermine Phosphate Inhibits The SARS-CoV-2 Spike-ACE2 Protein-Protein Interaction-As An in Silico Approach Contribute To Its Antiviral Activity Against COVIDДокумент14 страницSpermine Phosphate Inhibits The SARS-CoV-2 Spike-ACE2 Protein-Protein Interaction-As An in Silico Approach Contribute To Its Antiviral Activity Against COVIDRakesh DavellaОценок пока нет
- In-Silico MOLECULAR DOCKING ANALYSIS OF ANDROGRAPHOLIDE DERIVED FROM Andrographis Paniculata AS POTENTIAL ANTI-HIV AGENT TARGETING HIV-1 REVERSE TRANSCRIPTASEДокумент7 страницIn-Silico MOLECULAR DOCKING ANALYSIS OF ANDROGRAPHOLIDE DERIVED FROM Andrographis Paniculata AS POTENTIAL ANTI-HIV AGENT TARGETING HIV-1 REVERSE TRANSCRIPTASERakesh DavellaОценок пока нет
- Swapna Gurrapu and Estari Mamidala - Medicinal Plants Used by Traditional Medicine Practitioners in The Management of HIV/AIDS-Related Diseases in Tribal Areas of Adilabad District, Telangana RegionДокумент7 страницSwapna Gurrapu and Estari Mamidala - Medicinal Plants Used by Traditional Medicine Practitioners in The Management of HIV/AIDS-Related Diseases in Tribal Areas of Adilabad District, Telangana RegionEstari MamidalaОценок пока нет
- Web of Science Int J Ls and Pharma ResДокумент16 страницWeb of Science Int J Ls and Pharma ResDr Estari MamidalaОценок пока нет
- An in Silico Approach For Identification of Inhibitors As A Potential Therapeutics Targeting SARS-Cov-2 ProteaseДокумент7 страницAn in Silico Approach For Identification of Inhibitors As A Potential Therapeutics Targeting SARS-Cov-2 ProteaseRakesh DavellaОценок пока нет
- Antihiv Phyll EmplicaДокумент6 страницAntihiv Phyll EmplicaDr Estari MamidalaОценок пока нет
- 2018 Satish PoojariДокумент4 страницы2018 Satish PoojariDr Estari MamidalaОценок пока нет
- 1 s2.0 S1018364720300458 MainДокумент6 страниц1 s2.0 S1018364720300458 MainDr Estari MamidalaОценок пока нет
- Spermine Phosphate Inhibits The SARS-CoV-2 Spike-ACE2 Protein-Protein Interaction-As An in Silico Approach Contribute To Its Antiviral Activity Against COVIDДокумент14 страницSpermine Phosphate Inhibits The SARS-CoV-2 Spike-ACE2 Protein-Protein Interaction-As An in Silico Approach Contribute To Its Antiviral Activity Against COVIDRakesh DavellaОценок пока нет
- 1 s2.0 S1319562X19303079 MainДокумент7 страниц1 s2.0 S1319562X19303079 MainDr Estari MamidalaОценок пока нет
- Inhibition of Human Immunodeficiency Virus HIV 1 Reverse TranscriptaseДокумент4 страницыInhibition of Human Immunodeficiency Virus HIV 1 Reverse TranscriptaseDr Estari MamidalaОценок пока нет
- 2012 Ijabpt 2012Документ4 страницы2012 Ijabpt 2012Dr Estari MamidalaОценок пока нет
- 3780 Speaker PDFДокумент1 страница3780 Speaker PDFEstari MamidalaОценок пока нет
- J Antivir AntiretrovirДокумент1 страницаJ Antivir AntiretrovirDr Estari MamidalaОценок пока нет
- In Vitro anti-HIV Activity of Crude Extracts From Tinospora CordifoliaДокумент1 страницаIn Vitro anti-HIV Activity of Crude Extracts From Tinospora CordifoliaDr Estari MamidalaОценок пока нет
- 2012 Eco Scan SPL IssueДокумент6 страниц2012 Eco Scan SPL IssueDr Estari MamidalaОценок пока нет
- BMC Infectious Diseasess PaperДокумент1 страницаBMC Infectious Diseasess PaperDr Estari MamidalaОценок пока нет
- The Effectiveness of Stem Cells Grafting On Regeneration of Dental Pulp: A Systematic Review and Meta-AnalysisДокумент11 страницThe Effectiveness of Stem Cells Grafting On Regeneration of Dental Pulp: A Systematic Review and Meta-AnalysisIJAR JOURNALОценок пока нет
- Bases Moleculares Del CáncerДокумент15 страницBases Moleculares Del CáncerMairim Adn-a JóasОценок пока нет
- Lab 2 Upload DocumentДокумент3 страницыLab 2 Upload DocumentgenevivaОценок пока нет
- Cinética Dos Granulócitos Na Saúde e Na Doença PDFДокумент260 страницCinética Dos Granulócitos Na Saúde e Na Doença PDFJorgeОценок пока нет
- Jones and KompalaДокумент27 страницJones and KompalaPramita SenОценок пока нет
- Rguhs Mbbs SyllabusДокумент15 страницRguhs Mbbs SyllabusPrabzinОценок пока нет
- Regulacion TranscripcionalДокумент27 страницRegulacion TranscripcionalPaulina CisnerosОценок пока нет
- Genome Christopher Nicklin v4 Full 20161230121108Документ10 177 страницGenome Christopher Nicklin v4 Full 20161230121108Christopher NicklinОценок пока нет
- Biology 13Th Edition Sylvia Mader Full ChapterДокумент67 страницBiology 13Th Edition Sylvia Mader Full Chapteroscar.gokey720100% (2)
- Cellular Respiration DefinitionДокумент9 страницCellular Respiration Definitionmaria genioОценок пока нет
- Prova de Ingles 3 Ano Ensino MedioДокумент2 страницыProva de Ingles 3 Ano Ensino MedioJC Campos100% (1)
- Tumor Suppressor GeneДокумент3 страницыTumor Suppressor GeneZiedTrikiОценок пока нет
- s12 Biology Public Questions KCДокумент10 страницs12 Biology Public Questions KCrevamanianОценок пока нет
- Dna Replication: By: Group 3Документ26 страницDna Replication: By: Group 3Jesse Kate GonzalesОценок пока нет
- Cell Division: Mitosis: Ligaya M. Dimaano Science TeacherДокумент36 страницCell Division: Mitosis: Ligaya M. Dimaano Science TeacherAily Milambiling DimaanoОценок пока нет
- BS Biotechnology: By: Azra Naheed Medical and Dental CollegeДокумент8 страницBS Biotechnology: By: Azra Naheed Medical and Dental CollegeabubakurОценок пока нет
- Types of BiodiversityДокумент9 страницTypes of BiodiversityFaiz Syed MohammedОценок пока нет
- TNT - Terapia de Nutricion TotalДокумент6 страницTNT - Terapia de Nutricion TotalJuana Rosa Martinez BonillaОценок пока нет
- Retail Staff Licensed Pharmacist in Indianapolis IN Resume Christopher WeihlerДокумент2 страницыRetail Staff Licensed Pharmacist in Indianapolis IN Resume Christopher WeihlerChristopherWeihler2Оценок пока нет
- Disadvantages of CloningДокумент6 страницDisadvantages of CloningEmilyn Mata CastilloОценок пока нет
- 4th Quarter - Science 10 - Performance TaskДокумент3 страницы4th Quarter - Science 10 - Performance Taskmykyla aledonОценок пока нет
- Maynard Smith - BibliographyДокумент7 страницMaynard Smith - BibliographyMarkvN2Оценок пока нет
- Curriculum Vitae: Maria BeihaghiДокумент19 страницCurriculum Vitae: Maria BeihaghiSamuel Embiza TadesseОценок пока нет
- 5' Race System ManualДокумент48 страниц5' Race System ManualPrabu DhanasekaranОценок пока нет
- Under The Sea: Distance Learning SupportДокумент17 страницUnder The Sea: Distance Learning SupportZykeria Hunt HamiltonОценок пока нет
- Medical Genetics and Genomics - 2022 - FinalДокумент357 страницMedical Genetics and Genomics - 2022 - FinalCsaba SzalaiОценок пока нет
- World Cotton Research Conference-5Документ594 страницыWorld Cotton Research Conference-5Barkat2012100% (3)
- Tools in BioinformaticsДокумент17 страницTools in BioinformaticsJane Guiron AballaОценок пока нет



































































![Synthesis, anticancer and antibacterial evaluation of novel (isopropylidene) uridine-[1,2,3]triazole hybrids Ranjith kumar Thatipamula a, Sirassu Narsimha a, Kumaraswamy Battula a, V. Rajendra Chary b, Estari Mamidala b, Nagavelli Vasudeva Reddy](https://imgv2-1-f.scribdassets.com/img/document/363567748/149x198/9427f87f47/1510319787?v=1)














































