Вам также может понравиться
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- AMI December 2017 Digital PDFДокумент48 страницAMI December 2017 Digital PDFminteiluminataОценок пока нет
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (895)
- 2004 Yucha-Gilbert Evidence Based Practice BFДокумент58 страниц2004 Yucha-Gilbert Evidence Based Practice BFdiego_vega_00Оценок пока нет
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- Toroidal Structure of The Assemblage Point AppendixДокумент5 страницToroidal Structure of The Assemblage Point AppendixminteiluminataОценок пока нет
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (399)
- Exploration of Quantum Consciousness - Enlighten Next - 2010 PDFДокумент7 страницExploration of Quantum Consciousness - Enlighten Next - 2010 PDFraduОценок пока нет
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (266)
- Chapter 6 Motivating EmployeesДокумент38 страницChapter 6 Motivating EmployeesCabdiqani XasanОценок пока нет
- What Is AphasiaДокумент4 страницыWhat Is AphasiaValentina DubОценок пока нет
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- HIGH CENTERS DR. BARBON and DR VILA 1Документ15 страницHIGH CENTERS DR. BARBON and DR VILA 1Yami C.Оценок пока нет
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- LSP1501 Assessment 13Документ15 страницLSP1501 Assessment 13Annemarie RossouwОценок пока нет
- Executive Function Skills Questionnaire PDFДокумент2 страницыExecutive Function Skills Questionnaire PDFNikaelly LagesОценок пока нет
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- Questionnaire On Compulsive BuyingДокумент5 страницQuestionnaire On Compulsive Buyingsiddhi jainОценок пока нет
- MODULE 5 (Understanding My Learners, Their Strengths, Needs, Interests and Experiences)Документ8 страницMODULE 5 (Understanding My Learners, Their Strengths, Needs, Interests and Experiences)Jayrald Mendoza100% (1)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- Depression PDFДокумент16 страницDepression PDFMakmur SejatiОценок пока нет
- 2006 Management of Peripheral Neuropathic Pain. Integrating Neurobiology, Neurodynamics and Clinical EvidenceДокумент14 страниц2006 Management of Peripheral Neuropathic Pain. Integrating Neurobiology, Neurodynamics and Clinical EvidenceDiego PaezОценок пока нет
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2259)
- Mental Disorders-A MindHeal Homeopathy CaseStudyДокумент2 страницыMental Disorders-A MindHeal Homeopathy CaseStudyDr. Anita SalunkheОценок пока нет
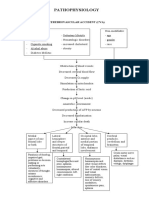
- PathophysiologyДокумент1 страницаPathophysiologynitlihpОценок пока нет
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (344)
- Face Ur Fears-Silva - Graff-2023-TICSДокумент13 страницFace Ur Fears-Silva - Graff-2023-TICSWinter Koidun FalgunОценок пока нет
- Models of CommunicationДокумент5 страницModels of CommunicationJoel PagkalinawaОценок пока нет
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- Cesario Johnson Eisthen IN PRESS CDPS Your Brain Is Not An OnionДокумент24 страницыCesario Johnson Eisthen IN PRESS CDPS Your Brain Is Not An OnionAna ChicasОценок пока нет
- Harvard Linguistics 130 SyllabusДокумент3 страницыHarvard Linguistics 130 SyllabusJ100% (4)
- 10 CS1AC16 Artificial Intelligence SummaryДокумент3 страницы10 CS1AC16 Artificial Intelligence SummaryAnna AbcxyzОценок пока нет
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- Instructions:: Sincerely! TrueДокумент40 страницInstructions:: Sincerely! TrueKeith RiveraОценок пока нет
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- An Overview of Popular Deep Learning Methods PDFДокумент12 страницAn Overview of Popular Deep Learning Methods PDFOussama El B'charriОценок пока нет
- Geronaga Elizabeth A. Module 10Документ4 страницыGeronaga Elizabeth A. Module 10elizabeth2geronagaОценок пока нет
- 7 Mind-Blowing Psychological Facts You Didnt Know About YourselfДокумент4 страницы7 Mind-Blowing Psychological Facts You Didnt Know About YourselfAsfaq SumraОценок пока нет
- Mind ReadingДокумент21 страницаMind ReadingMukund Varshney100% (1)
- The Preoperational Stage of Cognitive DevelopmentДокумент4 страницыThe Preoperational Stage of Cognitive DevelopmentJane QuintosОценок пока нет
- Color Consciouness: Beyond The GateДокумент4 страницыColor Consciouness: Beyond The GateMoMo25% (4)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (121)
- NACINOPA - FS 1 Learning Episode 9Документ11 страницNACINOPA - FS 1 Learning Episode 9Rolando Nacinopa Jr.Оценок пока нет
- Learning Task 4Документ6 страницLearning Task 4Jomar DiegoОценок пока нет
- Psychology: (8th Edition, in Modules) David MyersДокумент27 страницPsychology: (8th Edition, in Modules) David Myersmrchubs100% (2)
- The Hierarchical Model of Approach-Avoidance MotivationДокумент6 страницThe Hierarchical Model of Approach-Avoidance MotivationRhix RheeОценок пока нет
- Verbal Behavior Assessment FormДокумент7 страницVerbal Behavior Assessment FormMaura LangeОценок пока нет
- Topic 4 Schematic ConventionsДокумент18 страницTopic 4 Schematic ConventionsVasneas NewОценок пока нет
- Aphasia and Kindred Disorders of SpeechДокумент576 страницAphasia and Kindred Disorders of SpeechAntonio MadrigalОценок пока нет