Вам также может понравиться
- Recent Advances in Polyphenol ResearchОт EverandRecent Advances in Polyphenol ResearchCelestino Santos-BuelgaОценок пока нет
- Ezymatic BrowningДокумент16 страницEzymatic BrowningCHIEMEZIEM NJOKUОценок пока нет
- Precursori LicopenДокумент12 страницPrecursori LicopenDiana Madalina MitrutОценок пока нет
- Positive Effects of Temperature and Growth Conditions On Enzymatic and Antioxidant Status in Lettuce PlantsДокумент6 страницPositive Effects of Temperature and Growth Conditions On Enzymatic and Antioxidant Status in Lettuce PlantsDiyar SaleemОценок пока нет
- PF 33 ClasificareДокумент22 страницыPF 33 Clasificaregiorgiana19Оценок пока нет
- 2009 Fowler KoffasДокумент11 страниц2009 Fowler KoffasJenny MarfàОценок пока нет
- Biosintesis Derivados FlavonoidesДокумент7 страницBiosintesis Derivados FlavonoidesFanny AlmeydaОценок пока нет
- Caracterizacion Polifenol UchuvaДокумент29 страницCaracterizacion Polifenol UchuvaLESLY JHANELA CRUZ ROMOОценок пока нет
- Chlorophyll Oxidative Metabolism During TheДокумент15 страницChlorophyll Oxidative Metabolism During Therafael wadniparОценок пока нет
- BMB 21083Документ7 страницBMB 21083ShintaОценок пока нет
- Sweet Oranges PeelsДокумент9 страницSweet Oranges PeelsMirella FerreiraОценок пока нет
- Léchaudel Et Al 2018Документ12 страницLéchaudel Et Al 2018Jandira CostaОценок пока нет
- Bioavailability of Anthocyanins and DerivativesДокумент13 страницBioavailability of Anthocyanins and DerivativesUno MendozaОценок пока нет
- Agronomy: The Effect of Foliar Application of K SO or KH PO On Skin Color of The Kyoho' GrapeДокумент14 страницAgronomy: The Effect of Foliar Application of K SO or KH PO On Skin Color of The Kyoho' Grapemashhoodullah samadiОценок пока нет
- Phenyl AlanДокумент9 страницPhenyl AlanAibhlinxxОценок пока нет
- Extracelular ProteinsДокумент11 страницExtracelular ProteinsKary TéllezОценок пока нет
- Enzymatic Browning in Fruit2Документ68 страницEnzymatic Browning in Fruit2Ruben MarquezОценок пока нет
- Production of Commercial and Amazonian Strains of PleurotusДокумент13 страницProduction of Commercial and Amazonian Strains of PleurotusJaqueline da Silva Coelho MoreiraОценок пока нет
- Substrate Specificity of Polyphenol Oxidase: Critical Reviews in Biochemistry and Molecular BiologyДокумент36 страницSubstrate Specificity of Polyphenol Oxidase: Critical Reviews in Biochemistry and Molecular BiologyAnh Phương TrầnОценок пока нет
- Yoruk 2003Документ62 страницыYoruk 2003Gerson OrtegaОценок пока нет
- Symposium Report Nadph - Cytochrome P450 Oxidoreductase: Roles in Physiology, Pharmacology, and ToxicologyДокумент12 страницSymposium Report Nadph - Cytochrome P450 Oxidoreductase: Roles in Physiology, Pharmacology, and ToxicologyNimra Naveed ShaikhОценок пока нет
- Biodegradation of Phenol ReviewДокумент60 страницBiodegradation of Phenol ReviewMigue HerreraОценок пока нет
- Flavonoids Analgesic AgentДокумент17 страницFlavonoids Analgesic AgentStella AguirreОценок пока нет
- 2mtblts Extraction2Документ28 страниц2mtblts Extraction2Vijay KumarОценок пока нет
- Cabaton I In. 2009Документ10 страницCabaton I In. 2009Ola ZygmuntowiczОценок пока нет
- Antioxidant Activity of Vitamin E and Trolox: Understanding of The Factors That Govern Lipid Peroxidation Studies in VitroДокумент9 страницAntioxidant Activity of Vitamin E and Trolox: Understanding of The Factors That Govern Lipid Peroxidation Studies in VitroJoana FerreiraОценок пока нет
- Suratman 2015Документ15 страницSuratman 2015PatySA2Оценок пока нет
- Chung S.Yang (2008) - Bioavailability Issues in Studying The Health Efects of Plant Polyphenolic CompoundsДокумент13 страницChung S.Yang (2008) - Bioavailability Issues in Studying The Health Efects of Plant Polyphenolic CompoundsthanhktОценок пока нет
- Lucock 2011 Folic Acid Beyond MetabolismДокумент12 страницLucock 2011 Folic Acid Beyond Metabolismadinda.larastitiОценок пока нет
- AppleДокумент13 страницAppleAyishaОценок пока нет
- QTL and Candidate Gene Mapping For Polyphenolic Composition in Apple FruitДокумент16 страницQTL and Candidate Gene Mapping For Polyphenolic Composition in Apple FruitVianney CastaÂ¥Eda MonroyОценок пока нет
- Food Chemistry: Lei Zhou, Zhiqiang Xiong, Wei Liu, Liqiang ZouДокумент10 страницFood Chemistry: Lei Zhou, Zhiqiang Xiong, Wei Liu, Liqiang ZouQuenguan FranklinОценок пока нет
- Vol4 SPL (1) P 38-51Документ14 страницVol4 SPL (1) P 38-51Edris AslamiОценок пока нет
- Carotenoid Accumulation Affects Redox Status, Starch Metabolism, and Flavonoid/anthocyanin Accumulation in CitrusДокумент16 страницCarotenoid Accumulation Affects Redox Status, Starch Metabolism, and Flavonoid/anthocyanin Accumulation in CitrusAntonellaОценок пока нет
- Study of Phenolic Compounds As Natural Antioxidants by A Fluorescence MethodДокумент8 страницStudy of Phenolic Compounds As Natural Antioxidants by A Fluorescence MethodLavanya Priya SathyanОценок пока нет
- Enzymatic Determination of Catechol Oxidase and Protease From Fruits (Orange, Apple) and Vegetables (Carrot, Tomato)Документ7 страницEnzymatic Determination of Catechol Oxidase and Protease From Fruits (Orange, Apple) and Vegetables (Carrot, Tomato)International Organization of Scientific Research (IOSR)Оценок пока нет
- Polyphenols, Food Sources and BioavailabilityДокумент21 страницаPolyphenols, Food Sources and BioavailabilityMariaОценок пока нет
- Food Chemistry: Arzu Altunkaya, Vural GökmenДокумент5 страницFood Chemistry: Arzu Altunkaya, Vural Gökmenmr.andОценок пока нет
- Polyphenols Methods of ExtractionДокумент6 страницPolyphenols Methods of ExtractionWidayat WahyuОценок пока нет
- 1 s2.0 S0308814622010202 MainДокумент12 страниц1 s2.0 S0308814622010202 Mainborbala bereckiОценок пока нет
- Pro LinaДокумент14 страницPro LinaDaniela SepulvedaОценок пока нет
- Food and Chemical ToxicologyДокумент7 страницFood and Chemical ToxicologyDaniel Felipe Garzon GalloОценок пока нет
- Ferulic Acid and Its Therapeutic Potential As A HormetinДокумент12 страницFerulic Acid and Its Therapeutic Potential As A HormetinDaniel GrajalesОценок пока нет
- In Vitro Antioxidant Versus Metal Ion Chelating PRДокумент21 страницаIn Vitro Antioxidant Versus Metal Ion Chelating PRDr. Awanish MishraОценок пока нет
- 559 350 1 PBДокумент5 страниц559 350 1 PBSabar SupendiОценок пока нет
- Comparative Analysis of Antioxidant Properties of Water Extract of Centepede Grass and FestucaДокумент3 страницыComparative Analysis of Antioxidant Properties of Water Extract of Centepede Grass and FestucaInternational Journal of Application or Innovation in Engineering & ManagementОценок пока нет
- Moduladores Da Paraoxonase 2011 PDFДокумент8 страницModuladores Da Paraoxonase 2011 PDFMohamedElshbinyОценок пока нет
- Phenolic CompoundsДокумент16 страницPhenolic CompoundsLucíaОценок пока нет
- Antioxidants: How Does The Phenol Structure Influence The Results of The Folin-Ciocalteu Assay?Документ13 страницAntioxidants: How Does The Phenol Structure Influence The Results of The Folin-Ciocalteu Assay?Youcef BenamorОценок пока нет
- Biomedicines 08 00342Документ19 страницBiomedicines 08 00342Franco CibroОценок пока нет
- Mechanism of Fruit RipeningДокумент28 страницMechanism of Fruit RipeningDivya DivyachilaОценок пока нет
- Zhang 2013Документ8 страницZhang 2013Jeff Willian Rosas QuinaОценок пока нет
- Rocha 2001Документ7 страницRocha 2001ChristopherОценок пока нет
- Oliveira 2009Документ6 страницOliveira 2009metabolismeproteinОценок пока нет
- Thesis On Antioxidant Activity of Medicinal Plants PDFДокумент6 страницThesis On Antioxidant Activity of Medicinal Plants PDFafazapfjl100% (2)
- Effect of Flavonoids On Glutathione Level, Lipid Peroxidation and Cytochrome P450 CYP1A1 Expression in Human Laryngeal Carcinoma Cell LinesДокумент11 страницEffect of Flavonoids On Glutathione Level, Lipid Peroxidation and Cytochrome P450 CYP1A1 Expression in Human Laryngeal Carcinoma Cell LinesEric GibsonОценок пока нет
- Phenol 3 PDFДокумент10 страницPhenol 3 PDFsegОценок пока нет
- Review - Flavonoids A Powerful and Abundant Source of AntioxidantsДокумент4 страницыReview - Flavonoids A Powerful and Abundant Source of AntioxidantsPencari Ilmu100% (1)
- Extraction, Separation, Detection, and Structural Analysis of FlavonoidsДокумент26 страницExtraction, Separation, Detection, and Structural Analysis of FlavonoidsHusnatul AyniahОценок пока нет
- 639 FullДокумент12 страниц639 Fullahmed salahОценок пока нет
- CPVC SpecificationДокумент2 страницыCPVC Specificationviralnanobio_4150420Оценок пока нет
- FinishingSchool EED 25-8-2015Документ2 страницыFinishingSchool EED 25-8-2015viralnanobio_4150420Оценок пока нет
- Advanced Quality Engineering Techniques (AQET 2015) : Teqip - II Sponsored Short Term Training Programme OnДокумент2 страницыAdvanced Quality Engineering Techniques (AQET 2015) : Teqip - II Sponsored Short Term Training Programme Onviralnanobio_4150420Оценок пока нет
- Conf Seminar Brochure 2016Документ9 страницConf Seminar Brochure 2016viralnanobio_4150420Оценок пока нет
- Cotton CoatingДокумент3 страницыCotton Coatingviralnanobio_4150420Оценок пока нет
- Maintenance Manual For Laboratory EquipmentДокумент77 страницMaintenance Manual For Laboratory Equipmentjonathan_calixto_4Оценок пока нет
- Plant Innate Immunity: Sub. - Biochem-506, IMMUNO CHEMISTRYДокумент22 страницыPlant Innate Immunity: Sub. - Biochem-506, IMMUNO CHEMISTRYviralnanobio_4150420Оценок пока нет
- EE130: Integrated Circuit Devices: Course OutlineДокумент12 страницEE130: Integrated Circuit Devices: Course Outlineviralnanobio_4150420Оценок пока нет
- 27membrane ScienceДокумент16 страниц27membrane Scienceviralnanobio_4150420Оценок пока нет
- Agriculture WasteДокумент20 страницAgriculture Wastemohammadyunus1992Оценок пока нет
- PHD ThesisДокумент154 страницыPHD Thesisviralnanobio_4150420Оценок пока нет
- The Hydrogen EconomyДокумент21 страницаThe Hydrogen Economyviralnanobio_4150420Оценок пока нет
- Targeting Signal Transduction: George Weber, Fei Shen, Tam !as I. Orb!an, Szabolcs K .Okeny, Edith OlahДокумент10 страницTargeting Signal Transduction: George Weber, Fei Shen, Tam !as I. Orb!an, Szabolcs K .Okeny, Edith Olahviralnanobio_4150420Оценок пока нет
- Pukrushpan ThesisДокумент147 страницPukrushpan Thesisviralnanobio_4150420Оценок пока нет
- CellCell Webdez13Документ37 страницCellCell Webdez13viralnanobio_4150420Оценок пока нет
- Full Text 01Документ53 страницыFull Text 01viralnanobio_4150420Оценок пока нет
- Phytochemical Analysis and Antioxidant Potential of Cucumis Melo SeedsДокумент5 страницPhytochemical Analysis and Antioxidant Potential of Cucumis Melo SeedsSSR-IIJLS JournalОценок пока нет
- Antioxidant and Antiradical Activities of Rosa Canina PDFДокумент8 страницAntioxidant and Antiradical Activities of Rosa Canina PDFMarina DintiuОценок пока нет
- CH - 11 Alcohols Phenols and EthersДокумент34 страницыCH - 11 Alcohols Phenols and Ethersankitpaul5410Оценок пока нет
- Oxalate Ion in GuavaДокумент9 страницOxalate Ion in GuavaCiandeb GracianoОценок пока нет
- ThymeДокумент6 страницThymeAnonymous 4PTbXjPuZОценок пока нет
- SCNC 2015 Abstract BookДокумент307 страницSCNC 2015 Abstract BookNuray NiyeОценок пока нет
- Enzymatic and Bacterial Conversions During Sourdough FermentationДокумент9 страницEnzymatic and Bacterial Conversions During Sourdough FermentationroadrunnerОценок пока нет
- Colorimetry of Total Phenolics With Phosphomolybdic-Phosphotungstic Acid Reagents PDFДокумент1 страницаColorimetry of Total Phenolics With Phosphomolybdic-Phosphotungstic Acid Reagents PDFMygame50% (2)
- Chirinos Et Al., 2007 PDFДокумент7 страницChirinos Et Al., 2007 PDFwitaОценок пока нет
- Pharmacological and Therapeutic Effects of Jasminum Sambac - A ReviewДокумент13 страницPharmacological and Therapeutic Effects of Jasminum Sambac - A ReviewBaru Chandrasekhar RaoОценок пока нет
- Phenolic Profile and Antioxidant Activity of Melon (Cucumis Melo L.) Seeds From PakistanДокумент7 страницPhenolic Profile and Antioxidant Activity of Melon (Cucumis Melo L.) Seeds From PakistanGalinaОценок пока нет
- Phytochemicals and Biological Activities of Pulicaria Species.Документ23 страницыPhytochemicals and Biological Activities of Pulicaria Species.radovanborovcaninОценок пока нет
- Food Chemistry: Sahameh Shafiee, Saeid Minaei, Nasrollah Moghaddam-Charkari, Mohsen BarzegarДокумент8 страницFood Chemistry: Sahameh Shafiee, Saeid Minaei, Nasrollah Moghaddam-Charkari, Mohsen BarzegarDiana Carolina Acosta OpayomeОценок пока нет
- Enzymatic Browning in Fruit2Документ68 страницEnzymatic Browning in Fruit2Ruben MarquezОценок пока нет
- Food Chemistry: Merve Tomas, Jules Beekwilder, Robert D. Hall, Osman Sagdic, Dilek Boyacioglu, Esra CapanogluДокумент8 страницFood Chemistry: Merve Tomas, Jules Beekwilder, Robert D. Hall, Osman Sagdic, Dilek Boyacioglu, Esra CapanogluDiego AiresОценок пока нет
- S C I e N C e SДокумент7 страницS C I e N C e SEdy TansilОценок пока нет
- Alelopatia en HorticulturaДокумент38 страницAlelopatia en HorticulturaAlfredo M LuckieОценок пока нет
- Chapter 14Документ51 страницаChapter 14Varalaxmi BandariОценок пока нет
- Antioxidan 1Документ43 страницыAntioxidan 1pardeepbthОценок пока нет
- ANTIMICROBIAL ACTIVITY OF OREGANO (Origanum Vulgare L.) AND BASILДокумент6 страницANTIMICROBIAL ACTIVITY OF OREGANO (Origanum Vulgare L.) AND BASILJaime TerrazoОценок пока нет
- Phytochemicals, Antioxidant Activity and Phenolic Profiling of Diplocyclos Palmatus (L.) C. JefferyДокумент6 страницPhytochemicals, Antioxidant Activity and Phenolic Profiling of Diplocyclos Palmatus (L.) C. JefferyJovanJiEunMiОценок пока нет
- Poster Presentation Ga2007Документ31 страницаPoster Presentation Ga2007Paula_CastilhoОценок пока нет
- Microwave-Assisted Extraction of Phenolic Compounds From Olive Leaves A Comparison With MacerationДокумент8 страницMicrowave-Assisted Extraction of Phenolic Compounds From Olive Leaves A Comparison With MacerationThu LeОценок пока нет
- HDEPДокумент5 страницHDEPkmsrajuОценок пока нет
- Antiproliferative Activities of Common Edible Nut SeedsДокумент8 страницAntiproliferative Activities of Common Edible Nut SeedsAzekeОценок пока нет
- Antioxidant Properties of Various Solvent Extracts of Total Phenolic Constituents From Three Different Agroclimatic Origins of Drumstick Tree (Moringa Oleifera Lam.) LeavesДокумент12 страницAntioxidant Properties of Various Solvent Extracts of Total Phenolic Constituents From Three Different Agroclimatic Origins of Drumstick Tree (Moringa Oleifera Lam.) LeavesMeiОценок пока нет
- Antioxidant Capacity of Phenolic Phytochemicals From Various Cultivars of PlumsДокумент6 страницAntioxidant Capacity of Phenolic Phytochemicals From Various Cultivars of PlumsSevcan ErsanОценок пока нет
- Journal Buah Ketapang (Ficus Lyrata)Документ5 страницJournal Buah Ketapang (Ficus Lyrata)MuhammadRifqiAbdurrahmanОценок пока нет
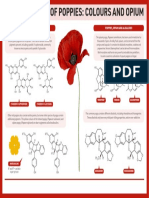
- The Chemistry of PoppiesДокумент1 страницаThe Chemistry of PoppiesShruti SinghalОценок пока нет
- Promociones Farmindustria - Dicar: Division Cod Sap Producto Escala Descuento Precio Final Codig O Precio RegularДокумент4 страницыPromociones Farmindustria - Dicar: Division Cod Sap Producto Escala Descuento Precio Final Codig O Precio RegularMerly CasasОценок пока нет
- University of Chicago Press Fall 2009 Distributed TitlesОт EverandUniversity of Chicago Press Fall 2009 Distributed TitlesРейтинг: 1 из 5 звезд1/5 (1)