Вам также может понравиться
- Multiphoton Optical Image Guided Spectroscopy Method For Characterization of Collagen-Based Materials Modified by GlycationДокумент7 страницMultiphoton Optical Image Guided Spectroscopy Method For Characterization of Collagen-Based Materials Modified by GlycationRui PereiraОценок пока нет
- Characterization of Secondary and Tertiary Conformational Changes of Â-Lactoglobulin Adsorbed On Silica Nanoparticle SurfacesДокумент10 страницCharacterization of Secondary and Tertiary Conformational Changes of Â-Lactoglobulin Adsorbed On Silica Nanoparticle SurfacesAnonymous 1DQJYzQ3J1Оценок пока нет
- 284Документ12 страниц284SilvanaMedhatОценок пока нет
- Self-Assembly of Amphiphilic Perylene - Cyclodextrin Conjugate and Vapor Sensing For Organic AminesДокумент7 страницSelf-Assembly of Amphiphilic Perylene - Cyclodextrin Conjugate and Vapor Sensing For Organic AminesDiogomussumОценок пока нет
- Total Synthesis of WedelolactoneДокумент5 страницTotal Synthesis of WedelolactoneTommy BetyОценок пока нет
- Tong - Viral Capsid DNA Aptamer Conjugates As Multivalent Cell-Targeting VehiclesДокумент5 страницTong - Viral Capsid DNA Aptamer Conjugates As Multivalent Cell-Targeting VehiclesChris TranОценок пока нет
- Regulation of Macromolecular Synthesis By Low Molecular Weight MediatorsОт EverandRegulation of Macromolecular Synthesis By Low Molecular Weight MediatorsGebhard KochОценок пока нет
- Gold Nanoparticle Size Controlled by Polymeric Au (I) Thiolate Precursor SizeДокумент8 страницGold Nanoparticle Size Controlled by Polymeric Au (I) Thiolate Precursor SizeleonardoОценок пока нет
- Heteroaryl Cross-Coupling As An Entry Toward The Synthesis of Lavendamycin Analogues: A Model StudyДокумент10 страницHeteroaryl Cross-Coupling As An Entry Toward The Synthesis of Lavendamycin Analogues: A Model StudySilvanaMedhatОценок пока нет
- Progress in Biophysics and Molecular BiologyОт EverandProgress in Biophysics and Molecular BiologyJ. A. V. ButlerОценок пока нет
- Chemistry and Biology of Nucleosides and NucleotidesОт EverandChemistry and Biology of Nucleosides and NucleotidesRobert HarmonОценок пока нет
- Org. Lett., Vol. 15, No. 3, 2013, 432-435Документ4 страницыOrg. Lett., Vol. 15, No. 3, 2013, 432-435Ramdas BorhadeОценок пока нет
- Poly (Dimethylsiloxane) - Based Protein Preconcentration Using A Nanogap Generated by Junction Gap BreakdownДокумент6 страницPoly (Dimethylsiloxane) - Based Protein Preconcentration Using A Nanogap Generated by Junction Gap BreakdownHai Lai QuangОценок пока нет
- D Hayal 2009Документ7 страницD Hayal 2009Bahaaa ZeroОценок пока нет
- Complexation Between Dodecyl Sulfate SurДокумент4 страницыComplexation Between Dodecyl Sulfate Suryeqiang liОценок пока нет
- Zhou 2012Документ14 страницZhou 2012ZAIB KOHLIОценок пока нет
- Optical Fiber Sensor For BODДокумент6 страницOptical Fiber Sensor For BODeveltoncОценок пока нет
- Protides of the Biological Fluids: Proceedings of the Twenty-Fifth Colloquium, Brugge, 1977От EverandProtides of the Biological Fluids: Proceedings of the Twenty-Fifth Colloquium, Brugge, 1977H. PeetersОценок пока нет
- Easley 2009Документ10 страницEasley 2009AmirОценок пока нет
- Zno Nanoparticle ThesisДокумент8 страницZno Nanoparticle Thesismariaparkslasvegas100% (2)
- Multiple Processing Streams in Occipitotemporal Visual CortexДокумент1 страницаMultiple Processing Streams in Occipitotemporal Visual CortexViorelDiorducОценок пока нет
- Biochemical Factors Concerned in the Functional Activity of the Nervous System: First International Meeting of the International Society for Neurochemistry, Strasbourg, 1967От EverandBiochemical Factors Concerned in the Functional Activity of the Nervous System: First International Meeting of the International Society for Neurochemistry, Strasbourg, 1967D. RichterОценок пока нет
- The Use Of Radioautography in Investigating Protein SynthesisОт EverandThe Use Of Radioautography in Investigating Protein SynthesisC LeblondОценок пока нет
- Maximizing Gene ExpressionОт EverandMaximizing Gene ExpressionWilliam ReznikoffОценок пока нет
- Carbon-13 NMR Spectroscopy of Biological SystemsОт EverandCarbon-13 NMR Spectroscopy of Biological SystemsNicolau BeckmannОценок пока нет
- Surfactant Interactions With Zein ProteinДокумент6 страницSurfactant Interactions With Zein Proteinyeqiang liОценок пока нет
- Systems Biology and Synthetic BiologyОт EverandSystems Biology and Synthetic BiologyPengcheng FuОценок пока нет
- Fine-Needle Aspiration Biopsy of the Rat Liver: Cytological, Cytochemical and Biochemical MethodsОт EverandFine-Needle Aspiration Biopsy of the Rat Liver: Cytological, Cytochemical and Biochemical MethodsG. ZbindenОценок пока нет
- Proteins: Analysis and DesignОт EverandProteins: Analysis and DesignRuth Hogue AngelettiОценок пока нет
- Communications: The Molecular Structures of Poly (Ethylene Glycol) - Modified Nonviral Gene Delivery PolyplexesДокумент6 страницCommunications: The Molecular Structures of Poly (Ethylene Glycol) - Modified Nonviral Gene Delivery PolyplexesNIMESHKAMDARОценок пока нет
- General PrinciplesОт EverandGeneral PrinciplesRonald AloiaОценок пока нет
- Ions-Cyclic Nucleotides-Cholinergy: Proceedings of the 7th International Congress of Pharmacology, Paris, 1978От EverandIons-Cyclic Nucleotides-Cholinergy: Proceedings of the 7th International Congress of Pharmacology, Paris, 1978J.C. StocletОценок пока нет
- 2013 Multiple Cytotoxic and Genotoxic Effects Induced in Vitro by Differently Shaped Copper Oxide NanomaterialsДокумент13 страниц2013 Multiple Cytotoxic and Genotoxic Effects Induced in Vitro by Differently Shaped Copper Oxide NanomaterialsRamonquimicoОценок пока нет
- GPI Membrane AnchorsОт EverandGPI Membrane AnchorsM AlmeidaОценок пока нет
- Two-Step Synthesis of Aza-And Diazaindoles From Chloroamino - N-Heterocycles Using EthoxyvinylborolaneДокумент5 страницTwo-Step Synthesis of Aza-And Diazaindoles From Chloroamino - N-Heterocycles Using EthoxyvinylborolaneSilvanaMedhatОценок пока нет
- Two-Step Hantzsch Based Macrocyclization Approach For The Synthesis of Thiazole-Containing CyclopeptidesДокумент3 страницыTwo-Step Hantzsch Based Macrocyclization Approach For The Synthesis of Thiazole-Containing CyclopeptidesDiogomussumОценок пока нет
- Natural Products Analysis: Instrumentation, Methods, and ApplicationsОт EverandNatural Products Analysis: Instrumentation, Methods, and ApplicationsVladimir HavlicekОценок пока нет
- Langmuir MultiparametricДокумент10 страницLangmuir MultiparametricShibsekhar RoyОценок пока нет
- Processing and Turnover of Proteins and Organelles in the Cell: FEBS Federation of European Biochemical Societies: 12th Meeting, Dresden, 1978От EverandProcessing and Turnover of Proteins and Organelles in the Cell: FEBS Federation of European Biochemical Societies: 12th Meeting, Dresden, 1978S. RapoportОценок пока нет
- Antimetabolites in Biochemistry, Biology and Medicine: Proceedings of a Symposium on Antimetabolites in Biochemistry, Biology and Medicine, Held in Prague, Czechoslovakia, July 10-12, 1978От EverandAntimetabolites in Biochemistry, Biology and Medicine: Proceedings of a Symposium on Antimetabolites in Biochemistry, Biology and Medicine, Held in Prague, Czechoslovakia, July 10-12, 1978J. SkodaРейтинг: 5 из 5 звезд5/5 (1)
- Spectroscopic Characterization of Spin-Labeled Magnetically Oriented Phospholipid Bilayers by EPR SpectrosДокумент7 страницSpectroscopic Characterization of Spin-Labeled Magnetically Oriented Phospholipid Bilayers by EPR SpectrosSveti JeronimОценок пока нет
- 2PA CdSe-ZnSДокумент4 страницы2PA CdSe-ZnSlabasvisogeroОценок пока нет
- 225Документ35 страниц225dianaОценок пока нет
- Schering Workshop on Steroid Hormone 'Receptors', Berlin, December 7 to 9, 1970: Advances in the BiosciencesОт EverandSchering Workshop on Steroid Hormone 'Receptors', Berlin, December 7 to 9, 1970: Advances in the BiosciencesGerhard RaspéОценок пока нет
- Introduction AssignmentДокумент2 страницыIntroduction Assignmentfelicia.wОценок пока нет
- AffimerДокумент35 страницAffimerİzem DevecioğluОценок пока нет
- Synthesis of N-And N-Labeled Trityl-Nitroxide Biradicals With Strong Spin - Spin Interaction and Improved Sensitivity To Redox Status and OxygenДокумент7 страницSynthesis of N-And N-Labeled Trityl-Nitroxide Biradicals With Strong Spin - Spin Interaction and Improved Sensitivity To Redox Status and OxygenDiogomussumОценок пока нет
- 108 SynopsisДокумент3 страницы108 SynopsisAbhilipsa MohantyОценок пока нет
- BenzylideneimidazolinoneДокумент7 страницBenzylideneimidazolinoneEdwar CortesОценок пока нет
- Coan & Shoichet, 2008 Stoichiometry and Physical Chemistry of Promiscuous AggregatorsДокумент7 страницCoan & Shoichet, 2008 Stoichiometry and Physical Chemistry of Promiscuous AggregatorsKonkmanОценок пока нет
- Intrinsically Disordered Proteins: Dynamics, Binding, and FunctionОт EverandIntrinsically Disordered Proteins: Dynamics, Binding, and FunctionNicola SalviОценок пока нет
- PDFДокумент3 страницыPDFsreejaОценок пока нет
- Regulatory Proteolytic Enzymes and Their Inhibitors: 11th Meeting Copenhagen 1977От EverandRegulatory Proteolytic Enzymes and Their Inhibitors: 11th Meeting Copenhagen 1977Staffan MagnussonОценок пока нет
- Concepts of Membrane StructureОт EverandConcepts of Membrane StructureRonald AloiaОценок пока нет
- Ultrathin Coatings From Isocyanate-Terminated Star PEGДокумент9 страницUltrathin Coatings From Isocyanate-Terminated Star PEGGanesh KshirsagarОценок пока нет
- Nanoscale: PaperДокумент7 страницNanoscale: Paperirajfarji2481Оценок пока нет
- Low Endotoxin Recovery (LER) in Drug Products - Original - 31428Документ2 страницыLow Endotoxin Recovery (LER) in Drug Products - Original - 31428Putri DozanОценок пока нет
- Kinji Asaka, Hidenori Okuzaki (Eds.) - Soft Actuators - Materials, Modeling, Applications, and Future Perspectives (2014, Springer Japan)Документ492 страницыKinji Asaka, Hidenori Okuzaki (Eds.) - Soft Actuators - Materials, Modeling, Applications, and Future Perspectives (2014, Springer Japan)Deependra Kumar BanОценок пока нет
- Green Chemistry A Greener CleanДокумент21 страницаGreen Chemistry A Greener CleanDeependra Kumar BanОценок пока нет
- Introduction To Optical Spectroscopy: Chemistry 243Документ90 страницIntroduction To Optical Spectroscopy: Chemistry 243Deependra Kumar BanОценок пока нет
- Chapter 3c X Ray DiffractionДокумент51 страницаChapter 3c X Ray DiffractionDeependra Kumar Ban100% (1)
- ImagoVisual A0 1189MMx841MMДокумент1 страницаImagoVisual A0 1189MMx841MMDeependra Kumar BanОценок пока нет
- UV Vis LectureДокумент33 страницыUV Vis LectureJulie AlexanderОценок пока нет
- 2 Introduction To XRPD Data AnalysisДокумент25 страниц2 Introduction To XRPD Data AnalysisMuthu KumarОценок пока нет
- 2 Introduction To XRPD Data AnalysisДокумент25 страниц2 Introduction To XRPD Data AnalysisMuthu KumarОценок пока нет
- 005 Andrew Hufton ScientificData BSBDДокумент19 страниц005 Andrew Hufton ScientificData BSBDDeependra Kumar BanОценок пока нет
- 007 Veerle Van Den Eynden Nature ShareSensitiveDate 2014-11-14Документ18 страниц007 Veerle Van Den Eynden Nature ShareSensitiveDate 2014-11-14Deependra Kumar BanОценок пока нет
- Indian Institute Of Technology Kharagpur: विज्ञापन सं: / Advertisement No.: R/02/2015 Dated April 01, 2015Документ4 страницыIndian Institute Of Technology Kharagpur: विज्ञापन सं: / Advertisement No.: R/02/2015 Dated April 01, 2015Deependra Kumar BanОценок пока нет
- 001 Phil Campbell Reproducibility and Data at Nature NPG 13-11-14Документ34 страницы001 Phil Campbell Reproducibility and Data at Nature NPG 13-11-14Deependra Kumar BanОценок пока нет
- 005 Andrew Hufton ScientificData BSBDДокумент19 страниц005 Andrew Hufton ScientificData BSBDDeependra Kumar BanОценок пока нет
- ImagoVisual A0 1189MMx841MMДокумент1 страницаImagoVisual A0 1189MMx841MMDeependra Kumar BanОценок пока нет
- 005 Andrew Hufton ScientificData BSBDДокумент19 страниц005 Andrew Hufton ScientificData BSBDDeependra Kumar BanОценок пока нет
- Coated Nanoparticles Show Alzheimer's Promise - Chemistry WorldДокумент1 страницаCoated Nanoparticles Show Alzheimer's Promise - Chemistry WorldDeependra Kumar BanОценок пока нет
- 001 Phil Campbell Reproducibility and Data at Nature NPG 13-11-14Документ34 страницы001 Phil Campbell Reproducibility and Data at Nature NPG 13-11-14Deependra Kumar BanОценок пока нет
- GPU Accelerated Molecular Dynamics - SHARPENДокумент5 страницGPU Accelerated Molecular Dynamics - SHARPENDeependra Kumar BanОценок пока нет
- 004 Digital Academia The Future of ResearchДокумент40 страниц004 Digital Academia The Future of ResearchDeependra Kumar BanОценок пока нет
- Einstein PhysicsДокумент21 страницаEinstein PhysicsDeependra Kumar BanОценок пока нет
- NanoparticleДокумент18 страницNanoparticleDeependra Kumar BanОценок пока нет
- TransportДокумент39 страницTransportDeependra Kumar BanОценок пока нет
- Nanosciencenanotechnologyspain2011 111213045640 Phpapp02Документ167 страницNanosciencenanotechnologyspain2011 111213045640 Phpapp02Deependra Kumar Ban100% (1)
- ATCC Bacterial Culture GuideДокумент35 страницATCC Bacterial Culture GuidePramoth NarashimmanОценок пока нет
- Mechanism of The Eukaryotic Chaperonin: Protein Folding in The Chamber of SecretsДокумент7 страницMechanism of The Eukaryotic Chaperonin: Protein Folding in The Chamber of SecretsDeependra Kumar BanОценок пока нет
- 3 2. Principles and Applications of Surface Plasmon Resonance. 4Документ50 страниц3 2. Principles and Applications of Surface Plasmon Resonance. 4Deependra Kumar BanОценок пока нет
- EP500 Standard Electropneumatic Positioner: Installation and Maintenance InstructionsДокумент28 страницEP500 Standard Electropneumatic Positioner: Installation and Maintenance InstructionsAlan ValdezОценок пока нет
- 3 Activities For Adults To Practice Modeling SELДокумент10 страниц3 Activities For Adults To Practice Modeling SELDavid Garcia PerezОценок пока нет
- Bacterial Genome Assembly IlluminaДокумент49 страницBacterial Genome Assembly IlluminadksaОценок пока нет
- GEC - ReviewerДокумент23 страницыGEC - ReviewerGlycel BagabagonОценок пока нет
- 17-003 MK Media Kit 17Документ36 страниц17-003 MK Media Kit 17Jean SandiОценок пока нет
- Meat Plant FeasabilityДокумент115 страницMeat Plant FeasabilityCh WaqasОценок пока нет
- Notes Marriage and Family in Canon LawДокумент5 страницNotes Marriage and Family in Canon LawmacОценок пока нет
- Soalan 9 LainДокумент15 страницSoalan 9 LainMelor DihatiОценок пока нет
- Pe 3 Syllabus - GymnasticsДокумент7 страницPe 3 Syllabus - GymnasticsLOUISE DOROTHY PARAISO100% (1)
- AtelectasisДокумент37 страницAtelectasisSandara ParkОценок пока нет
- ISO - TR - 15608 - 2017 (En) - Pipe Grouping SystemsДокумент12 страницISO - TR - 15608 - 2017 (En) - Pipe Grouping SystemsTeodor ProdanОценок пока нет
- Unit 8 Packet KeyДокумент21 страницаUnit 8 Packet KeyHiddenОценок пока нет
- A Comprehensive Review of Cementitious Grouts - Composition, Properties, Requirements and Advanced PerformanceДокумент16 страницA Comprehensive Review of Cementitious Grouts - Composition, Properties, Requirements and Advanced PerformanceiporrasОценок пока нет
- Sample SWMSДокумент4 страницыSample SWMSJuma KavesuОценок пока нет
- Scope: Procter and GambleДокумент30 страницScope: Procter and GambleIrshad AhamedОценок пока нет
- 204-04B - Tire Pressure Monitoring System (TPMS)Документ23 страницы204-04B - Tire Pressure Monitoring System (TPMS)Sofia AltuzarraОценок пока нет
- Eric Koech CV - Docx..bakДокумент7 страницEric Koech CV - Docx..bakPeter Osundwa KitekiОценок пока нет
- 10 2005 Dec QДокумент6 страниц10 2005 Dec Qspinster40% (1)
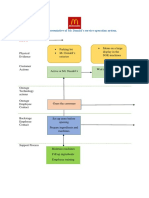
- Blueprint Huynh My Ky Duyen 2022 McDonald'sДокумент2 страницыBlueprint Huynh My Ky Duyen 2022 McDonald'sHuỳnh Mỹ Kỳ DuyênОценок пока нет
- G1 Series User Manual Ver. 1.2Документ101 страницаG1 Series User Manual Ver. 1.2unedo parhusip100% (1)
- ASD Diagnosis Tools - UpToDateДокумент3 страницыASD Diagnosis Tools - UpToDateEvy Alvionita Yurna100% (1)
- BV Lesson Plan 4Документ3 страницыBV Lesson Plan 4api-252119803Оценок пока нет
- Soil SSCДокумент11 страницSoil SSCvkjha623477Оценок пока нет
- ACF5950 - Assignment # 7 Semester 2 2015: The Business Has The Following Opening Balances: Additional InformationДокумент2 страницыACF5950 - Assignment # 7 Semester 2 2015: The Business Has The Following Opening Balances: Additional InformationkietОценок пока нет
- Libro Resumenes 2012Документ735 страницLibro Resumenes 2012fdobonat613100% (2)
- Engineering Project ListДокумент25 страницEngineering Project ListSyed ShaОценок пока нет
- Education in America: The Dumbing Down of The U.S. Education SystemДокумент4 страницыEducation in America: The Dumbing Down of The U.S. Education SystemmiichaanОценок пока нет
- 1A Wound Care AdviceДокумент2 страницы1A Wound Care AdviceGrace ValenciaОценок пока нет
- Join Our Telegram Channel: @AJITLULLA: To Get Daily Question Papers & SolutionsДокумент24 страницыJoin Our Telegram Channel: @AJITLULLA: To Get Daily Question Papers & SolutionsNaveen KumarОценок пока нет
- AMS 48 - 2000-n - D0114354 - 055 - 00Документ116 страницAMS 48 - 2000-n - D0114354 - 055 - 00wanhall100% (1)