Вам также может понравиться
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- Stuart Card (Editor) - Thomas P. Moran - Allen Newell - The Psychology of Human-Computer Interaction-Chapman and Hall - CRC (2017) - 40-114Документ75 страницStuart Card (Editor) - Thomas P. Moran - Allen Newell - The Psychology of Human-Computer Interaction-Chapman and Hall - CRC (2017) - 40-114Shabnam SmileОценок пока нет
- ECS401: Cryptography and Network Security: Module 5: Authentication ProtocolsДокумент19 страницECS401: Cryptography and Network Security: Module 5: Authentication ProtocolsShabnam SmileОценок пока нет
- ECS401: Cryptography and Network SecurityДокумент32 страницыECS401: Cryptography and Network SecurityShabnam SmileОценок пока нет
- ECS401: Cryptography and Network Security: Module 5: Authentication ProtocolsДокумент19 страницECS401: Cryptography and Network Security: Module 5: Authentication ProtocolsShabnam SmileОценок пока нет
- ECS401: Cryptography and Network Security: Module 5: Authentication ProtocolsДокумент25 страницECS401: Cryptography and Network Security: Module 5: Authentication ProtocolsShabnam SmileОценок пока нет
- ECS401: Cryptography and Network Security: Module 5: Authentication ProtocolsДокумент15 страницECS401: Cryptography and Network Security: Module 5: Authentication ProtocolsShabnam SmileОценок пока нет
- ECS401: Cryptography and Network Security: Module 5: Authentication ProtocolsДокумент17 страницECS401: Cryptography and Network Security: Module 5: Authentication ProtocolsShabnam SmileОценок пока нет
- ECS401: Cryptography and Network Security: Module 1: IntroductionДокумент24 страницыECS401: Cryptography and Network Security: Module 1: IntroductionShabnam SmileОценок пока нет
- ECS401: Cryptography and Network Security: Module 5: Malicious SoftwareДокумент31 страницаECS401: Cryptography and Network Security: Module 5: Malicious SoftwareShabnam SmileОценок пока нет
- Evoked and Event-Related Potentials (ERP) Event Related Potentials (ERP) : Basics (Part 1)Документ23 страницыEvoked and Event-Related Potentials (ERP) Event Related Potentials (ERP) : Basics (Part 1)Shabnam SmileОценок пока нет
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (400)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (895)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (266)
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (345)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (121)
- Pediatric Acute Lymphoblastic LeukemiaДокумент9 страницPediatric Acute Lymphoblastic LeukemiaAlvin PratamaОценок пока нет
- Osteopathy InfographicДокумент1 страницаOsteopathy Infographicapi-614436724Оценок пока нет
- DOH-HFSRB-QOP01Form1 Rev2 6172022Документ3 страницыDOH-HFSRB-QOP01Form1 Rev2 6172022clarisse salvadorОценок пока нет
- Jsa For Complete Erection of Tank-001Документ52 страницыJsa For Complete Erection of Tank-001Ashutosh80% (10)
- Sciatic AdviceДокумент3 страницыSciatic AdviceJohn Fightakis100% (2)
- StatusДокумент3 страницыStatusjobertОценок пока нет
- Vac Lok Cushions GuideДокумент9 страницVac Lok Cushions GuideYS TangОценок пока нет
- Breech Presentation and DeliveryДокумент7 страницBreech Presentation and DeliverySteven Herbert0% (1)
- Completing The Partogram in Pregnancy 5.0Документ9 страницCompleting The Partogram in Pregnancy 5.0fei cuaОценок пока нет
- CSS Clinical Guidelines Final Oct 14Документ2 страницыCSS Clinical Guidelines Final Oct 14francesco2vivianoОценок пока нет
- On Job Description 1Документ65 страницOn Job Description 1subodh nagarОценок пока нет
- Parorexia and HomoeopathyДокумент10 страницParorexia and HomoeopathyDr. Rajneesh Kumar Sharma MD HomОценок пока нет
- Evaluating The Neurologic Status of Unconscious PatientДокумент3 страницыEvaluating The Neurologic Status of Unconscious Patientsambel_88Оценок пока нет
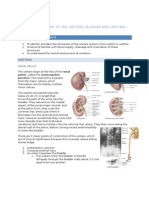
- Lecture 2 - Anatomy of The Ureters, Bladder and UrethraДокумент7 страницLecture 2 - Anatomy of The Ureters, Bladder and UrethraNatalie LiОценок пока нет
- NTS Book For GAT GeneralДокумент4 страницыNTS Book For GAT GeneralAbdul Mudabbir Rehan100% (1)
- 2015 Article 110Документ9 страниц2015 Article 110Boleac HaralambieОценок пока нет
- SYMPO14 1. REV. 1 DR. MUHADI - Diagnosis and Risk Stratification For SCAD Role of ImagingДокумент14 страницSYMPO14 1. REV. 1 DR. MUHADI - Diagnosis and Risk Stratification For SCAD Role of Imagingbagus krisnaОценок пока нет
- The Beauty of DentistryДокумент5 страницThe Beauty of DentistryMorosanu Diana GeorgianaОценок пока нет
- Lacosamide TreatmentДокумент13 страницLacosamide TreatmentawinsyОценок пока нет
- Delayed Replantation of An Avulsed Maxillary Premolar With Open Apex: A 24 Months Follow-Up Case ReportДокумент5 страницDelayed Replantation of An Avulsed Maxillary Premolar With Open Apex: A 24 Months Follow-Up Case ReportDiego Andres Hincapie HerreraОценок пока нет
- Anatomia Orientada para A Clínica - 00022Документ1 страницаAnatomia Orientada para A Clínica - 00022pablotmartinsОценок пока нет
- 380 L MethylfolateДокумент2 страницы380 L MethylfolateBhupender Singh AroraОценок пока нет
- 1 Prismaflexcrrtintro Seg12007 110326040426 Phpapp02Документ38 страниц1 Prismaflexcrrtintro Seg12007 110326040426 Phpapp02Sami Maghrebi100% (1)
- Anal FissureДокумент1 страницаAnal Fissureshailesh284Оценок пока нет
- Justins ResumeДокумент1 страницаJustins Resumeapi-250710016Оценок пока нет
- Final Pediatric Basic Life SupportДокумент26 страницFinal Pediatric Basic Life SupportMohamed Ghanem100% (2)
- Distinctions Dental 2008Документ64 страницыDistinctions Dental 2008Ellery July100% (3)
- Bowel Preparation For Pediatric Colonoscopy AnakДокумент8 страницBowel Preparation For Pediatric Colonoscopy AnakrantaikarbonОценок пока нет
- Anatomy and Physiology of The SkinДокумент40 страницAnatomy and Physiology of The SkinRicko Ciady100% (1)
- IMEI HypertensionДокумент5 страницIMEI HypertensionparkdhingraОценок пока нет