Вам также может понравиться
- Pathology for Toxicologists: Principles and Practices of Laboratory Animal Pathology for Study PersonnelОт EverandPathology for Toxicologists: Principles and Practices of Laboratory Animal Pathology for Study PersonnelElizabeth McInnesОценок пока нет
- Biochimica Et Biophysica ActaДокумент7 страницBiochimica Et Biophysica ActaMax JimenezОценок пока нет
- Cell Biology Assays: ProteinsОт EverandCell Biology Assays: ProteinsFanny JaulinОценок пока нет
- Phagocytosis of Apoptotic Cells by LiverДокумент11 страницPhagocytosis of Apoptotic Cells by Liverthanhthangphan8347Оценок пока нет
- Apoptotic Clearance RegulationДокумент15 страницApoptotic Clearance Regulationare4erОценок пока нет
- 1756 9966 30 87 PDFДокумент14 страниц1756 9966 30 87 PDFkarayevОценок пока нет
- Apoptosis Programmed Cell Death and Its SignalsДокумент11 страницApoptosis Programmed Cell Death and Its Signalsonur ateşОценок пока нет
- The Role of Apoptosis in Disease and DevelopmentДокумент3 страницыThe Role of Apoptosis in Disease and DevelopmentŠišmišova DevojkaОценок пока нет
- Apoptosis LengkapДокумент31 страницаApoptosis Lengkapduhita dyahОценок пока нет
- 1 6apoptosisДокумент8 страниц1 6apoptosisElena PoriazovaОценок пока нет
- ApoptosisДокумент16 страницApoptosisonur ateşОценок пока нет
- International Journal of Pharmaceutical Science Invention (IJPSI)Документ8 страницInternational Journal of Pharmaceutical Science Invention (IJPSI)inventionjournalsОценок пока нет
- Mitochondrial ApoptosisДокумент5 страницMitochondrial ApoptosisRuheen MariamОценок пока нет
- Autophagy Cellular and Molecular MechanismsДокумент10 страницAutophagy Cellular and Molecular MechanismsWilliam LeeОценок пока нет
- Apoptosis Tutorial NotesДокумент8 страницApoptosis Tutorial NotesismealОценок пока нет
- The Biology of Apoptosis: Fouad Boulos, MD August 2010Документ4 страницыThe Biology of Apoptosis: Fouad Boulos, MD August 2010ans11Оценок пока нет
- Mitochondrial Membrane PermeabilizationДокумент66 страницMitochondrial Membrane PermeabilizationGus Tavo BrugesОценок пока нет
- Methods: Guest Editor's IntroductionДокумент3 страницыMethods: Guest Editor's Introductionfsdf6554Оценок пока нет
- J Gerontol A Biol Sci Med Sci-2001-Pollack-B475-82Документ8 страницJ Gerontol A Biol Sci Med Sci-2001-Pollack-B475-82divinebutler13Оценок пока нет
- Cell DДокумент56 страницCell DJhon Jairo Donato ArizaОценок пока нет
- Programmed Cell Death in Plants: Narcin Palavan-Unsal, Elif-Damla Buyuktuncer and Mehmet Ali TufekciДокумент23 страницыProgrammed Cell Death in Plants: Narcin Palavan-Unsal, Elif-Damla Buyuktuncer and Mehmet Ali TufekciSourav GuptaОценок пока нет
- Apoptosis in Acute Kidney InjuryДокумент9 страницApoptosis in Acute Kidney InjuryRiszki PaeОценок пока нет
- Mi 04 005Документ11 страницMi 04 005Jefferson CaesarioОценок пока нет
- Cell Death 2011Документ15 страницCell Death 2011Paula Contreras MezaОценок пока нет
- Apoptosis: Proliferation and Physiological Cell Death, Apoptosis. Both of These ProcessesДокумент3 страницыApoptosis: Proliferation and Physiological Cell Death, Apoptosis. Both of These ProcesseswaheedkabОценок пока нет
- Apoptosis and Cancer: Insights Molecular Mechanisms and TreatmentsДокумент16 страницApoptosis and Cancer: Insights Molecular Mechanisms and TreatmentsOpenaccess Research paperОценок пока нет
- Pyroptosis: A Caspase-1-Dependent Programmed Cell Death and A Barrier To InfectionДокумент21 страницаPyroptosis: A Caspase-1-Dependent Programmed Cell Death and A Barrier To Infectionrovia achmad .Оценок пока нет
- Apoptosis y CancerДокумент7 страницApoptosis y CancerKatherine RiveraОценок пока нет
- Biochemistry of Apoptotic Cell Death: Acta Pharm. 53 (2003) 151-164Документ14 страницBiochemistry of Apoptotic Cell Death: Acta Pharm. 53 (2003) 151-164Gayathri MaigandanОценок пока нет
- Apoptosis DefinitionДокумент3 страницыApoptosis DefinitionHera ElykaОценок пока нет
- Cellular Injury, Necrosis and ApoptosisДокумент34 страницыCellular Injury, Necrosis and ApoptosisSuraj UsmanОценок пока нет
- Apoptosis FinalДокумент12 страницApoptosis FinalAtul Kumar GoyalОценок пока нет
- Chapter 11-ApoptosisДокумент26 страницChapter 11-ApoptosisKw ChanОценок пока нет
- Project: Debilitating Diseases of TissuesДокумент10 страницProject: Debilitating Diseases of TissuesDaniela GhicaОценок пока нет
- Cell Death and ClearanceДокумент41 страницаCell Death and ClearanceZhongming ChenОценок пока нет
- 1 Apoptosis DesДокумент7 страниц1 Apoptosis DessashiwiiОценок пока нет
- Programmed Cell DeathДокумент5 страницProgrammed Cell DeathAbhivanditaОценок пока нет
- Mitochondrial Ca and Apoptosis: Cell CalciumДокумент8 страницMitochondrial Ca and Apoptosis: Cell CalciumAhmad Ligar SuhermanОценок пока нет
- Izab 277Документ10 страницIzab 277MartinaОценок пока нет
- BEN503 MiniReview OATES 20223512006 2022fall PDFДокумент10 страницBEN503 MiniReview OATES 20223512006 2022fall PDFonur ateşОценок пока нет
- 2021 Kist - Cell Death PathwaysДокумент23 страницы2021 Kist - Cell Death PathwaysClaudio GarganoОценок пока нет
- ApoptosisДокумент3 страницыApoptosisاسماء زياد عبدالجبارОценок пока нет
- Chapter 7 - Mechanisms of Cell DeathДокумент22 страницыChapter 7 - Mechanisms of Cell DeathCynthia Lopes100% (1)
- Acquired Capability: Evading ApoptosisДокумент5 страницAcquired Capability: Evading ApoptosisKaren Joy MariñasОценок пока нет
- The Role of Mitochondria in Apoptosis - PMCДокумент34 страницыThe Role of Mitochondria in Apoptosis - PMCAnumol LoranceОценок пока нет
- Review Article: Apoptosis in YeastsДокумент6 страницReview Article: Apoptosis in YeastsShreya AhujaОценок пока нет
- Atlas of ApoptosisДокумент14 страницAtlas of ApoptosisAnonymous VECoBcRSОценок пока нет
- 12 - Autophagy in Human DiseasesДокумент13 страниц12 - Autophagy in Human DiseasesAndres Felipe Hurtado BautistaОценок пока нет
- Role of Autophagy and Apoptosis in Non-Small-Cell Lung CancerДокумент24 страницыRole of Autophagy and Apoptosis in Non-Small-Cell Lung CancerANGUIE ANETTE MAMANI SUCASAIREОценок пока нет
- New Microsoft Word DocumentДокумент4 страницыNew Microsoft Word DocumentsamiunОценок пока нет
- tmpC00F TMPДокумент7 страницtmpC00F TMPFrontiersОценок пока нет
- ApoptosisДокумент48 страницApoptosisLouella Gamoso Artates50% (2)
- Pathogenesis of Bacterial InfectionДокумент5 страницPathogenesis of Bacterial Infectionshahbaz100% (4)
- MINI: Identifying Enteropathogenic Escherichia Coli Effector Proteins Responsible For Blocking Host Proinflammatory Responses.Документ54 страницыMINI: Identifying Enteropathogenic Escherichia Coli Effector Proteins Responsible For Blocking Host Proinflammatory Responses.Mikeho123Оценок пока нет
- The Regulation of Aging - Does Autophagy Underlie Longevity - PMCДокумент15 страницThe Regulation of Aging - Does Autophagy Underlie Longevity - PMCParth RanaОценок пока нет
- Biotechnology Advances: Antonio L. Grilo, Athanasios Mantalaris TДокумент17 страницBiotechnology Advances: Antonio L. Grilo, Athanasios Mantalaris TAtrocitus RedОценок пока нет
- Jun Zou, Nathan ShankarДокумент11 страницJun Zou, Nathan ShankarHQ HQОценок пока нет
- Apoptosis PDFДокумент8 страницApoptosis PDFCuarto Paralelo 1Оценок пока нет
- TMP 6605Документ8 страницTMP 6605FrontiersОценок пока нет
- Jékely 2007 - Origin of Phagotrophic Eukaryotes As Social Cheaters in Microbial BiofilmsДокумент15 страницJékely 2007 - Origin of Phagotrophic Eukaryotes As Social Cheaters in Microbial BiofilmsBGEОценок пока нет
- Gyrase Inhibitors Induce An Oxidative Damage Cellular Death Pathway in Escherichia ColiДокумент15 страницGyrase Inhibitors Induce An Oxidative Damage Cellular Death Pathway in Escherichia ColiMariano PerezОценок пока нет
- Disulfide Bond Formation and Cysteine Exclusion inДокумент10 страницDisulfide Bond Formation and Cysteine Exclusion inMariano PerezОценок пока нет
- Peroxiredoxin Systems Structures and FunctionsДокумент409 страницPeroxiredoxin Systems Structures and FunctionsMariano PerezОценок пока нет
- The Ubiquitin-Proteasome Proteolytic System From Classical Biochemistry To Human DiseasesДокумент242 страницыThe Ubiquitin-Proteasome Proteolytic System From Classical Biochemistry To Human DiseasesMariano PerezОценок пока нет
- CHOP Induces Death by Promoting Protein Synthesis and Oxidation in The Stressed Endoplasmic ReticulumДокумент13 страницCHOP Induces Death by Promoting Protein Synthesis and Oxidation in The Stressed Endoplasmic ReticulumMariano PerezОценок пока нет
- Redox Sensing by Escherichia Coli Effects of DithiothreitolДокумент4 страницыRedox Sensing by Escherichia Coli Effects of DithiothreitolMariano PerezОценок пока нет
- Protein Dna RecognitionДокумент31 страницаProtein Dna RecognitionMariano PerezОценок пока нет
- A Pathway For Disulfide Bond Formation in VivoДокумент5 страницA Pathway For Disulfide Bond Formation in VivoMariano PerezОценок пока нет
- Targeting Metabolism To Diagnose and TreatДокумент4 страницыTargeting Metabolism To Diagnose and TreatMariano PerezОценок пока нет
- Contribution of Calcium Influx in Mediating Glucose-Stimulated Oxygen Consumption in Pancreatic IsletsДокумент11 страницContribution of Calcium Influx in Mediating Glucose-Stimulated Oxygen Consumption in Pancreatic IsletsMariano PerezОценок пока нет
- A Molecular Mechanism of Pyruvate Protection Against Cytotoxicity of Reactive Oxygen SpeciesДокумент11 страницA Molecular Mechanism of Pyruvate Protection Against Cytotoxicity of Reactive Oxygen SpeciesMariano PerezОценок пока нет
- Microplate-Based Kinetic Method For Assay of Mitochondrial NADH - and Succinate - Cytochrome C Reductase ActivitiesДокумент3 страницыMicroplate-Based Kinetic Method For Assay of Mitochondrial NADH - and Succinate - Cytochrome C Reductase ActivitiesMariano PerezОценок пока нет
- Selective Removal of Mitochondria Via MitophagyДокумент7 страницSelective Removal of Mitochondria Via MitophagyMariano PerezОценок пока нет
- Generation of Rho0 Cells Utilizing A Mitochondrially Targeted Restriction Endonuclease and Comparative AnalysesДокумент10 страницGeneration of Rho0 Cells Utilizing A Mitochondrially Targeted Restriction Endonuclease and Comparative AnalysesMariano PerezОценок пока нет
- Selective Removal of Mitochondria Via MitophagyДокумент7 страницSelective Removal of Mitochondria Via MitophagyMariano PerezОценок пока нет
- Oxidative Protein Folding Is DrivenДокумент11 страницOxidative Protein Folding Is DrivenMariano PerezОценок пока нет
- Phase II Trial of Temozolomide Plus O6-Benzylguanine in Adults With Recurrent, Temozolomide-Resistant Malignant GliomaДокумент6 страницPhase II Trial of Temozolomide Plus O6-Benzylguanine in Adults With Recurrent, Temozolomide-Resistant Malignant GliomaMariano PerezОценок пока нет
- Mitochondria/Cytosol Fractionation Kit: Sufficient For Analysis of 50 SamplesДокумент8 страницMitochondria/Cytosol Fractionation Kit: Sufficient For Analysis of 50 SamplesMariano PerezОценок пока нет
- NAD+ and NADH in Cellular Functions and Cell DeathДокумент3 страницыNAD+ and NADH in Cellular Functions and Cell DeathMariano PerezОценок пока нет
- Mitochondria/Cytosol Fractionation Kit: Sufficient For Analysis of 50 SamplesДокумент8 страницMitochondria/Cytosol Fractionation Kit: Sufficient For Analysis of 50 SamplesMariano PerezОценок пока нет
- The Rate of Oxygen Utilization by CellsДокумент32 страницыThe Rate of Oxygen Utilization by CellsMariano PerezОценок пока нет
- Reactive Oxygen Species in Cancer Cells Live by The Sword Die by The Sword 2006 Cancer CellДокумент2 страницыReactive Oxygen Species in Cancer Cells Live by The Sword Die by The Sword 2006 Cancer CellMariano PerezОценок пока нет
- Protein Folding in The Bacterial PeriplasmДокумент7 страницProtein Folding in The Bacterial PeriplasmMariano PerezОценок пока нет
- Fluorescent Dyes Introduction Ratiometric AssaysДокумент2 страницыFluorescent Dyes Introduction Ratiometric AssaysMariano PerezОценок пока нет
- Ratiometric Calcium IndicatorsДокумент1 страницаRatiometric Calcium IndicatorsMariano PerezОценок пока нет
- Oxidative Protein Folding Is DrivenДокумент11 страницOxidative Protein Folding Is DrivenMariano PerezОценок пока нет
- An Assessment of Prediction Algorithms For Nucleosome PositioningДокумент1 страницаAn Assessment of Prediction Algorithms For Nucleosome PositioningMariano PerezОценок пока нет
- Ratiometric Calcium IndicatorsДокумент1 страницаRatiometric Calcium IndicatorsMariano PerezОценок пока нет
- An Assessment of Prediction Algorithms For Nucleosome PositioningДокумент1 страницаAn Assessment of Prediction Algorithms For Nucleosome PositioningMariano PerezОценок пока нет
- Antibiotic Efficacy Is Linked To Bacterial Cellular Respiration.Документ8 страницAntibiotic Efficacy Is Linked To Bacterial Cellular Respiration.Mariano PerezОценок пока нет
- NW NSC GR 10 Life Sciences p1 Eng Nov 2019Документ12 страницNW NSC GR 10 Life Sciences p1 Eng Nov 2019lunabileunakhoОценок пока нет
- What Is Malnutrition?: WastingДокумент6 страницWhat Is Malnutrition?: WastingĐoan VõОценок пока нет
- Cage HomesДокумент8 страницCage HomesstanhopekrisОценок пока нет
- A Comparative Clinical Evaluation of Eranda Taila Matra Basti With Combination of Mefenamic Acid and Dicyclomine in The Management of Udavartini W.S.R To Primary DysmenorrheaДокумент8 страницA Comparative Clinical Evaluation of Eranda Taila Matra Basti With Combination of Mefenamic Acid and Dicyclomine in The Management of Udavartini W.S.R To Primary DysmenorrheaEditor IJTSRDОценок пока нет
- ESC - 2021 - The Growing Role of Genetics in The Understanding of Cardiovascular Diseases - Towards Personalized MedicineДокумент5 страницESC - 2021 - The Growing Role of Genetics in The Understanding of Cardiovascular Diseases - Towards Personalized MedicineDini SuhardiniОценок пока нет
- Neurobiology of Sleep: Madhu Kalia4Документ5 страницNeurobiology of Sleep: Madhu Kalia4Julian ReyesОценок пока нет
- Delayed Homicides and The Proximate Cause.8Документ5 страницDelayed Homicides and The Proximate Cause.8rheanditahafsaОценок пока нет
- Breast Imaging Review - A Quick Guide To Essential Diagnoses (2nd Edition) PDFДокумент264 страницыBreast Imaging Review - A Quick Guide To Essential Diagnoses (2nd Edition) PDFGregorio Parra100% (2)
- Program Implementation With The Health Team: Packages of Essential Services For Primary HealthcareДокумент1 страницаProgram Implementation With The Health Team: Packages of Essential Services For Primary Healthcare2A - Nicole Marrie HonradoОценок пока нет
- Bahasa InggrisДокумент6 страницBahasa InggrisSofi SusantoОценок пока нет
- Punjab Municipal Corporation Act, 1976 PDFДокумент180 страницPunjab Municipal Corporation Act, 1976 PDFSci UpscОценок пока нет
- Hemorrhoid - Pathophysiology and Surgical ManagementA LiteratureДокумент7 страницHemorrhoid - Pathophysiology and Surgical ManagementA LiteratureIndra YaniОценок пока нет
- 8mkebook PDFДокумент300 страниц8mkebook PDFVanja Vujovic100% (2)
- Therapy Enhancing CouplesДокумент22 страницыTherapy Enhancing CouplesluzОценок пока нет
- Amazing Facts About Our BrainДокумент4 страницыAmazing Facts About Our BrainUmarul FarooqueОценок пока нет
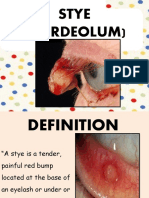
- StyeДокумент21 страницаStyenur syafiqah kamaruzamanОценок пока нет
- Pathophysiology of Ischemic Stroke FinalДокумент3 страницыPathophysiology of Ischemic Stroke FinalAcohCChao67% (3)
- IC Risk Assessment Worksheet - Kangas-V2.1-Aug.2010 1Документ4 страницыIC Risk Assessment Worksheet - Kangas-V2.1-Aug.2010 1Juon Vairzya AnggraeniОценок пока нет
- CATHETERIZATIONДокумент3 страницыCATHETERIZATIONrnrmmanphdОценок пока нет
- ScenarioДокумент1 страницаScenarioAngel Lynn YlayaОценок пока нет
- JC-2 Tutorial-1 Immunology Practice MCQ'sДокумент14 страницJC-2 Tutorial-1 Immunology Practice MCQ'shelamahjoubmounirdmo100% (5)
- Who Ten Step Management of Severe MalnutritionДокумент4 страницыWho Ten Step Management of Severe MalnutritionShely Karma Astuti33% (3)
- Mapeh: Music - Arts - Physical Education - HealthДокумент23 страницыMapeh: Music - Arts - Physical Education - HealthExtremelydarkness100% (1)
- Bentonite MSDSДокумент10 страницBentonite MSDSkhai ruleОценок пока нет
- Lesson 5 Core Elements Evidenced Based Gerontological Nursing PracticeДокумент38 страницLesson 5 Core Elements Evidenced Based Gerontological Nursing PracticeSam GarciaОценок пока нет
- Aac-Augmentative and Alternative CommunicationДокумент35 страницAac-Augmentative and Alternative Communicationrenuka aurangabadkerОценок пока нет
- Test Bank For Pilbeams Mechanical Ventilation 5th Edition CairoДокумент13 страницTest Bank For Pilbeams Mechanical Ventilation 5th Edition Cairochowryurduq0krhОценок пока нет
- Jurnal Bahasa InggrisДокумент9 страницJurnal Bahasa InggrisNur Riza Maulidina100% (1)
- Resveratrol and Its Effects On Human Health and LongevityДокумент367 страницResveratrol and Its Effects On Human Health and LongevityArnulfo Yu LanibaОценок пока нет
- Information On The Use of Domperidone To Increase Milk Production in Lactating WomenДокумент3 страницыInformation On The Use of Domperidone To Increase Milk Production in Lactating WomenKhairul HananОценок пока нет
- Why We Die: The New Science of Aging and the Quest for ImmortalityОт EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityРейтинг: 4 из 5 звезд4/5 (3)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessОт Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessРейтинг: 4 из 5 звезд4/5 (33)
- Gut: the new and revised Sunday Times bestsellerОт EverandGut: the new and revised Sunday Times bestsellerРейтинг: 4 из 5 звезд4/5 (392)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisОт EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisРейтинг: 3.5 из 5 звезд3.5/5 (2)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)От EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Рейтинг: 4 из 5 звезд4/5 (378)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceОт EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceРейтинг: 4.5 из 5 звезд4.5/5 (516)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsОт EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsРейтинг: 4.5 из 5 звезд4.5/5 (6)
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouОт EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouРейтинг: 4.5 из 5 звезд4.5/5 (62)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperОт EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperРейтинг: 4.5 из 5 звезд4.5/5 (15)
- Tales from Both Sides of the Brain: A Life in NeuroscienceОт EverandTales from Both Sides of the Brain: A Life in NeuroscienceРейтинг: 3 из 5 звезд3/5 (18)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesОт EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesРейтинг: 4.5 из 5 звезд4.5/5 (397)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldОт EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldРейтинг: 4.5 из 5 звезд4.5/5 (18)
- Moral Tribes: Emotion, Reason, and the Gap Between Us and ThemОт EverandMoral Tribes: Emotion, Reason, and the Gap Between Us and ThemРейтинг: 4.5 из 5 звезд4.5/5 (115)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionОт EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionРейтинг: 4 из 5 звезд4/5 (812)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedОт EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedРейтинг: 4 из 5 звезд4/5 (11)
- Who's in Charge?: Free Will and the Science of the BrainОт EverandWho's in Charge?: Free Will and the Science of the BrainРейтинг: 4 из 5 звезд4/5 (65)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorОт EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorОценок пока нет
- Good Without God: What a Billion Nonreligious People Do BelieveОт EverandGood Without God: What a Billion Nonreligious People Do BelieveРейтинг: 4 из 5 звезд4/5 (66)
- Seven and a Half Lessons About the BrainОт EverandSeven and a Half Lessons About the BrainРейтинг: 4 из 5 звезд4/5 (109)
- Lymph & Longevity: The Untapped Secret to HealthОт EverandLymph & Longevity: The Untapped Secret to HealthРейтинг: 4.5 из 5 звезд4.5/5 (13)
- The Rise and Fall of the Dinosaurs: A New History of a Lost WorldОт EverandThe Rise and Fall of the Dinosaurs: A New History of a Lost WorldРейтинг: 4 из 5 звезд4/5 (595)
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindОт EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindРейтинг: 4.5 из 5 звезд4.5/5 (93)
- Human: The Science Behind What Makes Your Brain UniqueОт EverandHuman: The Science Behind What Makes Your Brain UniqueРейтинг: 3.5 из 5 звезд3.5/5 (38)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildОт EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildРейтинг: 4.5 из 5 звезд4.5/5 (44)
- Change Your Brain, Change Your Life (Before 25): Change Your Developing Mind for Real-World SuccessОт EverandChange Your Brain, Change Your Life (Before 25): Change Your Developing Mind for Real-World SuccessРейтинг: 4 из 5 звезд4/5 (18)