Вам также может понравиться
- Plant-Herbivore Networks in The Tropics. In: Dáttilo W., Rico-Gray V. (Eds) Ecological Networks in The Tropics. SpringerДокумент17 страницPlant-Herbivore Networks in The Tropics. In: Dáttilo W., Rico-Gray V. (Eds) Ecological Networks in The Tropics. SpringerAntonio AntonioОценок пока нет
- Desidratador de Alimentos Avantco cfd10 1645570589Документ1 страницаDesidratador de Alimentos Avantco cfd10 1645570589Antonio AntonioОценок пока нет
- Siliwal & Ravichandran 2008Документ7 страницSiliwal & Ravichandran 2008Antonio AntonioОценок пока нет
- Siliwal & Ravichandran 2008Документ1 страницаSiliwal & Ravichandran 2008Antonio AntonioОценок пока нет
- Siliwal & Ravichandran 2008Документ1 страницаSiliwal & Ravichandran 2008Antonio AntonioОценок пока нет
- Alarcon 2008Документ12 страницAlarcon 2008Antonio AntonioОценок пока нет
- A Meta-Analysis of Preference-Performance HerbivoresДокумент11 страницA Meta-Analysis of Preference-Performance HerbivoresAntonio AntonioОценок пока нет
- Alarcon 2008Документ12 страницAlarcon 2008Antonio AntonioОценок пока нет
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (895)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (344)
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (399)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2259)
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (120)
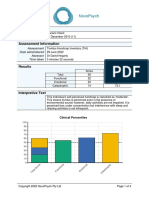
- Tinnitus Assessment Thi ScoringДокумент4 страницыTinnitus Assessment Thi ScoringbssОценок пока нет
- 2018 (Vuorien Et Al)Документ20 страниц2018 (Vuorien Et Al)gowthami mОценок пока нет
- Assignment II & III 208Документ9 страницAssignment II & III 208ramesh pokhrelОценок пока нет
- Completed Component 1 Learner Statement Y2Документ6 страницCompleted Component 1 Learner Statement Y2api-538905113Оценок пока нет
- FINA 4250 Applications of Risk ModelsДокумент67 страницFINA 4250 Applications of Risk ModelsChristopherОценок пока нет
- Raz Correlation ChartДокумент1 страницаRaz Correlation Chartapi-236762108Оценок пока нет
- Experimental Designs in Sentence Processing Research: A Methodological Review and User's GuideДокумент32 страницыExperimental Designs in Sentence Processing Research: A Methodological Review and User's Guidesara1744Оценок пока нет
- TOPAS 5 Technical Reference - UnlockedДокумент247 страницTOPAS 5 Technical Reference - UnlockedEdilberОценок пока нет
- Curriculum Vitae: Maria BeihaghiДокумент19 страницCurriculum Vitae: Maria BeihaghiSamuel Embiza TadesseОценок пока нет
- Daylight Factor - Wikipedia, The Free EncyclopediaДокумент2 страницыDaylight Factor - Wikipedia, The Free EncyclopediadasaОценок пока нет
- Declaration of Interdependence Steele 1976 2pgs GOV POLДокумент2 страницыDeclaration of Interdependence Steele 1976 2pgs GOV POLsonof libertyОценок пока нет
- English 7th Lesson Plan-1Документ7 страницEnglish 7th Lesson Plan-1SanaОценок пока нет
- Irlb8314Pbf: Application V 30 V R Max 2.4 M 3.2 QG 40 NC I 171 A I 130AДокумент8 страницIrlb8314Pbf: Application V 30 V R Max 2.4 M 3.2 QG 40 NC I 171 A I 130AJosè Miguel López RodriguezОценок пока нет
- Physics (Mass, Weight and Density)Документ2 страницыPhysics (Mass, Weight and Density)TVОценок пока нет
- Drum Brakes RT: According To DIN 15435Документ1 страницаDrum Brakes RT: According To DIN 15435LukaОценок пока нет
- ICETEMS-18 Abstract Book PDFДокумент152 страницыICETEMS-18 Abstract Book PDFJAMILОценок пока нет
- 597 - Semiconductor in Equilibrium&Pn Junction1Документ65 страниц597 - Semiconductor in Equilibrium&Pn Junction1dtizikaraОценок пока нет
- Practical MagnetotelluricsДокумент14 страницPractical MagnetotelluricsjesaduОценок пока нет
- TI-EN-UVACURID C81 FarbserieДокумент12 страницTI-EN-UVACURID C81 FarbseriePatxi hernandez suellesОценок пока нет
- Rainbow Water Stacking Influenced by Sugar Density and Principle of Flotation ExperimentДокумент11 страницRainbow Water Stacking Influenced by Sugar Density and Principle of Flotation ExperimentDan Luigi TipactipacОценок пока нет
- Lecture 8 Data Sources in GISДокумент32 страницыLecture 8 Data Sources in GISkhakhamasetiОценок пока нет
- Abaqus Automatic Stabilization When and How To Use ItДокумент8 страницAbaqus Automatic Stabilization When and How To Use ItBingkuai bigОценок пока нет
- Metu Thesis On CappadociaДокумент134 страницыMetu Thesis On CappadociaRamazanОценок пока нет
- SN 74 Ls 245Документ19 страницSN 74 Ls 245Luisdceo LggОценок пока нет
- CocoaДокумент24 страницыCocoaASONG ISMAELОценок пока нет
- Shadows of Esteren Universe PDFДокумент293 страницыShadows of Esteren Universe PDFThibault Mlt EricsonОценок пока нет
- Prof Ed QuestionsДокумент35 страницProf Ed QuestionsKatrina BautistaОценок пока нет
- Bozic Niksa 2018 Chardak Ni Na Zemlji Ni Na Nebu PrikazДокумент1 страницаBozic Niksa 2018 Chardak Ni Na Zemlji Ni Na Nebu PrikazSanja LoncarОценок пока нет
- Magnetic Reduction-To-The-Pole at Low Latitudes: Observations and ConsiderationsДокумент11 страницMagnetic Reduction-To-The-Pole at Low Latitudes: Observations and ConsiderationscarlosОценок пока нет
- 3 Probability DistributionsДокумент78 страниц3 Probability Distributionsha ssanОценок пока нет