Вам также может понравиться
- The Physiology of Cell Energy Production: CPTIPS.COM MonographsОт EverandThe Physiology of Cell Energy Production: CPTIPS.COM MonographsОценок пока нет
- The Science of Nutrition for Athletes: Understanding the Specific Nutritional Needs of Athletes for Optimal Performance and RecoveryОт EverandThe Science of Nutrition for Athletes: Understanding the Specific Nutritional Needs of Athletes for Optimal Performance and RecoveryОценок пока нет
- Lemak Dan OlahragaДокумент4 страницыLemak Dan OlahragaDefita Ayu SagitaОценок пока нет
- Fat Loading For Endurance SportsДокумент9 страницFat Loading For Endurance SportsshaqtimОценок пока нет
- High Carb Vs Fat SportsДокумент7 страницHigh Carb Vs Fat SportsYazen JoudehОценок пока нет
- Biomolecules: The Regulation of Fat Metabolism During Aerobic ExerciseДокумент32 страницыBiomolecules: The Regulation of Fat Metabolism During Aerobic ExerciseMarina LuttazziОценок пока нет
- HMS 4601 NUTRITION TO FUEL SPORTS PERFORMANCE - Case 2 Running On EmptyДокумент10 страницHMS 4601 NUTRITION TO FUEL SPORTS PERFORMANCE - Case 2 Running On Emptystijn.berkmortel1Оценок пока нет
- Chapter 2: Fuel Utilization and Muscle Metabolism During ExerciseДокумент14 страницChapter 2: Fuel Utilization and Muscle Metabolism During ExerciseArizal AbdullahОценок пока нет
- Fat Burning ZoneДокумент7 страницFat Burning ZoneJohnny YuillОценок пока нет
- Lipid Metabolism During Endurance ExerciseДокумент6 страницLipid Metabolism During Endurance ExerciseMTNC. César ZermeñoОценок пока нет
- Lipid MetabolismДокумент6 страницLipid Metabolismbhargavi gopalanОценок пока нет
- Kiens 2011 Fat OxidationДокумент7 страницKiens 2011 Fat Oxidationlinoco1Оценок пока нет
- Sports PhysiologyДокумент32 страницыSports PhysiologyTalpasai InturiОценок пока нет
- Understanding Fat OxidationДокумент10 страницUnderstanding Fat OxidationPaulo CesarОценок пока нет
- Fatty Acid Mobilization From Adipose Tissue During Exercise: Jeffrey F. HorowitzДокумент7 страницFatty Acid Mobilization From Adipose Tissue During Exercise: Jeffrey F. HorowitzDavidson De AraújoОценок пока нет
- Carbohydrates Play A Important Role in The Energetics Sustaining Working MusclesДокумент1 страницаCarbohydrates Play A Important Role in The Energetics Sustaining Working MusclesMina JarehОценок пока нет
- Is Lactic Acid A Four Letter Word?: by Patti and Warren Finke, Team OregonДокумент4 страницыIs Lactic Acid A Four Letter Word?: by Patti and Warren Finke, Team OregonАлександър Антоан НиколовОценок пока нет
- Forum 2 - 11 Nov 2023Документ4 страницыForum 2 - 11 Nov 2023arvinp89Оценок пока нет
- Muscle Glucose and Fatty Acid2Документ22 страницыMuscle Glucose and Fatty Acid2Richard S. RoxasОценок пока нет
- Screenshot 2020-03-20 at 21.27.13Документ3 страницыScreenshot 2020-03-20 at 21.27.13Denise KorkmazОценок пока нет
- Nutrition and Dietetic Recommendarions in TaekwondoДокумент30 страницNutrition and Dietetic Recommendarions in TaekwondoLatifah ZahrohОценок пока нет
- PE17MODULESДокумент14 страницPE17MODULESJasmine GañganОценок пока нет
- Fundamentals of Glycogen.Документ17 страницFundamentals of Glycogen.Kevin VillaОценок пока нет
- Forum 1 - OCT 2023Документ3 страницыForum 1 - OCT 2023arvinp89Оценок пока нет
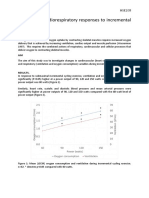
- Practical 4: Cardiorespiratory Responses To Incremental ExerciseДокумент4 страницыPractical 4: Cardiorespiratory Responses To Incremental ExercisePATRICK OTIATOОценок пока нет
- Burke (2007) Nutritional Strategies For The MarathonДокумент5 страницBurke (2007) Nutritional Strategies For The MarathonMario Pérez100% (1)
- 2x - The Science of Glycogen RepletionДокумент5 страниц2x - The Science of Glycogen RepletionMarco TorreОценок пока нет
- HMS 4601 Case 3Документ4 страницыHMS 4601 Case 3stijn.berkmortel1Оценок пока нет
- Fatmax A New Concept To Optimize Fat Oxidation DurДокумент6 страницFatmax A New Concept To Optimize Fat Oxidation Durjesus.clemente.90Оценок пока нет
- Fuel Choice During Exercise Is Determined by Intensity and Duration ofДокумент4 страницыFuel Choice During Exercise Is Determined by Intensity and Duration ofkrn_robledoОценок пока нет
- ATP Production From Carbs Protein FatsДокумент16 страницATP Production From Carbs Protein Fatshuraira bhattiОценок пока нет
- Exercise Physiology PDFДокумент31 страницаExercise Physiology PDFdupuytrenОценок пока нет
- Exercise Physiology Lectures - Chapters 1 - 5Документ15 страницExercise Physiology Lectures - Chapters 1 - 5TarienОценок пока нет
- Effects of Dietary Fat On Muscle Substrates,.11Документ9 страницEffects of Dietary Fat On Muscle Substrates,.11Saul NavaОценок пока нет
- Conference 2Документ7 страницConference 2naldghaimОценок пока нет
- Cycling Performance TipsДокумент6 страницCycling Performance TipscigarsnobОценок пока нет
- Training For Rowing: by Liz Flavell and Caroline Turnbull Why Train?Документ6 страницTraining For Rowing: by Liz Flavell and Caroline Turnbull Why Train?Nenad KamenovicОценок пока нет
- Carbohydrate Consumption and Glycogen StorageДокумент2 страницыCarbohydrate Consumption and Glycogen StorageОстојић СтрахињаОценок пока нет
- Gizi Kebugaran Pertemuan Iv Nazhif Gifari Ilmu Gizi & FIKESДокумент16 страницGizi Kebugaran Pertemuan Iv Nazhif Gifari Ilmu Gizi & FIKESrhatriОценок пока нет
- Sehs Topic 3 2 Carbohydrate and Fat MetabolismДокумент20 страницSehs Topic 3 2 Carbohydrate and Fat Metabolismapi-343368893Оценок пока нет
- Spriet2014 Article NewInsightsIntoTheInteractionOДокумент10 страницSpriet2014 Article NewInsightsIntoTheInteractionOaguaschma2114Оценок пока нет
- Nutrition NotesДокумент21 страницаNutrition NotesBobОценок пока нет
- Increased Fat Availability and Exercise PDFДокумент7 страницIncreased Fat Availability and Exercise PDFoskr1810Оценок пока нет
- 10 1152@japplphysiol 00907 2010Документ11 страниц10 1152@japplphysiol 00907 2010saulo freitas pereiraОценок пока нет
- Metabolic Adaptations To TrainingДокумент7 страницMetabolic Adaptations To TrainingMozil Fadzil Kamarudin100% (3)
- Physical Activity and Exercise in The Regulation of Human Adipose Tissue PhysiologyДокумент35 страницPhysical Activity and Exercise in The Regulation of Human Adipose Tissue PhysiologyLISTIA NURBAETIОценок пока нет
- Ajcn 138214Документ9 страницAjcn 138214Gustavo AlmeidaОценок пока нет
- Post Exercice Repletion of Muscle Energy Stores With Fructose or Glucose in Mixed MealsДокумент9 страницPost Exercice Repletion of Muscle Energy Stores With Fructose or Glucose in Mixed MealsLeonardo PinheiroОценок пока нет
- Bio Qui MicaДокумент9 страницBio Qui MicaLuciana Santos da SilvaОценок пока нет
- Three Components of Energy ExpenditureДокумент26 страницThree Components of Energy ExpenditureDustin HarrisonОценок пока нет
- Maximal Lipid Oxidation During Exercise (Lipoxmax)Документ15 страницMaximal Lipid Oxidation During Exercise (Lipoxmax)Juan David Salamanca BecerraОценок пока нет
- Strength EnduranceДокумент27 страницStrength EnduranceAdi HidayatОценок пока нет
- Lec34 F08 HandoutДокумент9 страницLec34 F08 HandoutVanessa VillespinОценок пока нет
- Carbohydrate-Loading and Exercise Performance: An UpdateДокумент9 страницCarbohydrate-Loading and Exercise Performance: An UpdateAdrián Masomenos AlastresОценок пока нет
- Effects of Ketone Bodies On Endurance Exercise: I F S MДокумент10 страницEffects of Ketone Bodies On Endurance Exercise: I F S MBrayam AguilarОценок пока нет
- Energy Metabolism1Документ8 страницEnergy Metabolism1edimulyanaОценок пока нет
- Topic 3 Energy1Документ68 страницTopic 3 Energy1api-343368893Оценок пока нет
- Integration of MetabolismДокумент40 страницIntegration of MetabolismSofie Hanafiah NuruddhuhaОценок пока нет
- Summary of Steve House, Scott Johnston & Kilian Jornet's Training for the Uphill AthleteОт EverandSummary of Steve House, Scott Johnston & Kilian Jornet's Training for the Uphill AthleteОценок пока нет
- Sports Nutrition Handbook: Eat Smart. Be Healthy. Get On Top of Your Game.От EverandSports Nutrition Handbook: Eat Smart. Be Healthy. Get On Top of Your Game.Оценок пока нет
- The Effects of Neuromuscular Training On Knee Joint MOTOR ControlДокумент9 страницThe Effects of Neuromuscular Training On Knee Joint MOTOR ControlalmaformaОценок пока нет
- Futsal Training Program Development For Higher Soccer Performance 1Документ2 страницыFutsal Training Program Development For Higher Soccer Performance 1almaformaОценок пока нет
- Agility Assessmentin Futsal FemaleДокумент6 страницAgility Assessmentin Futsal FemalealmaformaОценок пока нет
- FUTSALStrenght and ConditioningДокумент36 страницFUTSALStrenght and ConditioningalmaformaОценок пока нет
- Agility Assessmentin Futsal FemaleДокумент6 страницAgility Assessmentin Futsal FemalealmaformaОценок пока нет
- Futsal FitnessДокумент20 страницFutsal FitnessalmaformaОценок пока нет
- Speed Training Model in Futsal GameДокумент9 страницSpeed Training Model in Futsal GamealmaformaОценок пока нет
- FA Futsal Benefits Guidance Resource PDFДокумент6 страницFA Futsal Benefits Guidance Resource PDFalmaformaОценок пока нет
- Perth Youth Futsal Session PlannerДокумент4 страницыPerth Youth Futsal Session PlanneralmaformaОценок пока нет
- Reactivating and Strenghtning GluteosДокумент6 страницReactivating and Strenghtning GluteosalmaformaОценок пока нет
- Corrective Exercise The Lumbo Pelvic GirdleДокумент9 страницCorrective Exercise The Lumbo Pelvic GirdlealmaformaОценок пока нет
- Fifa Futsal Coaching ManualДокумент97 страницFifa Futsal Coaching ManualalmaformaОценок пока нет
- NutritionДокумент4 страницыNutritionalmaforma100% (1)
- Functional Anatomy and Muscle Action of The FootДокумент14 страницFunctional Anatomy and Muscle Action of The FootalmaformaОценок пока нет
- Feeding The Young AthleteДокумент20 страницFeeding The Young AthletealmaformaОценок пока нет
- Tennis Balls MassageДокумент7 страницTennis Balls MassagealmaformaОценок пока нет
- Exercise Order and Sequencing For Corrective Exercise ProgramsДокумент10 страницExercise Order and Sequencing For Corrective Exercise ProgramsalmaformaОценок пока нет
- Clinical Hip ExaminationДокумент90 страницClinical Hip ExaminationalmaformaОценок пока нет
- Synergy FullerДокумент320 страницSynergy FulleralmaformaОценок пока нет
- Granblue Fantasy - Wikipedia PDFДокумент32 страницыGranblue Fantasy - Wikipedia PDFRichard R.IgnacioОценок пока нет
- Jet ForceДокумент43 страницыJet ForcePanteley Panov ProОценок пока нет
- Suresh Raina - WikipediaДокумент14 страницSuresh Raina - WikipediaSPS Sri Harish . S G10Оценок пока нет
- UntitledДокумент4 страницыUntitledDedy W. HerdiyantoОценок пока нет
- Carcassonne + All Expansions (534335964)Документ1 страницаCarcassonne + All Expansions (534335964)Josue ReisОценок пока нет
- 1985 Centennial CupДокумент3 страницы1985 Centennial CupYob YnnosОценок пока нет
- Grade 11 Physics Take-Home Midterm TestДокумент5 страницGrade 11 Physics Take-Home Midterm TestkkkkkkkkkОценок пока нет
- Avp07aa AV110XCU3-QSB3.3-C99 4102011-XXXXXXX 100712 ENДокумент59 страницAvp07aa AV110XCU3-QSB3.3-C99 4102011-XXXXXXX 100712 ENIbaouene RamdaneОценок пока нет
- klf300 c15 c17 Parts List PDFДокумент97 страницklf300 c15 c17 Parts List PDFOliverОценок пока нет
- McLaren 750S Spider Order A8LCK24 Summary 2023-11-05Документ6 страницMcLaren 750S Spider Order A8LCK24 Summary 2023-11-05Ahmad Al-abdulghaniОценок пока нет
- Route Setting ManualДокумент7 страницRoute Setting Manualapi-268928582Оценок пока нет
- Tim Burroughs CVДокумент1 страницаTim Burroughs CVPiskaDrom228Оценок пока нет
- Equipment Schedule HVACДокумент7 страницEquipment Schedule HVACHussainShabbirОценок пока нет
- Daftar Game PS3 MaestrogameДокумент21 страницаDaftar Game PS3 MaestrogameAsliyanur Anueiy MОценок пока нет
- Louis Vuitton Out Let On Lines LVДокумент24 страницыLouis Vuitton Out Let On Lines LVLiu DehuaОценок пока нет
- Lista de Artículos Disponibles 27-Agosto-18 Devir MéxicoДокумент60 страницLista de Artículos Disponibles 27-Agosto-18 Devir MéxicoSigfridoOrgridОценок пока нет
- Narayana Dental PG Coaching - Week-03 Test (28-09-2014) Biochemistry (Enzymes, Vitamins & Miscellaneous)Документ5 страницNarayana Dental PG Coaching - Week-03 Test (28-09-2014) Biochemistry (Enzymes, Vitamins & Miscellaneous)drpnnreddyОценок пока нет
- Unit 14: at The Sports CentreДокумент2 страницыUnit 14: at The Sports CentrepeanutmilkОценок пока нет
- Team Sports: Volleyball Positions and RolesДокумент3 страницыTeam Sports: Volleyball Positions and RolesPATRICIA VISDA ASTORОценок пока нет
- 0 LK ZV9 MCДокумент190 страниц0 LK ZV9 MCawaleОценок пока нет
- Chevrolet Silverado & GMC Sierra Stock TrucksДокумент1 страницаChevrolet Silverado & GMC Sierra Stock TrucksJoshua EvbotokhaiОценок пока нет
- C20235-13554CN SMV67 Ecb90Документ4 страницыC20235-13554CN SMV67 Ecb90Abas AbasariОценок пока нет
- ConcussionsДокумент3 страницыConcussionsKarlie CapozzoliОценок пока нет
- Ankylosing SpondylitisДокумент24 страницыAnkylosing SpondylitisSiva ShanmugamОценок пока нет
- My 19 Aveo e Brochure enДокумент2 страницыMy 19 Aveo e Brochure enAnonymous 1HFV185Sl4Оценок пока нет
- Garena VsДокумент2 страницыGarena Vsapi-410426030Оценок пока нет
- Ford PPT MainДокумент23 страницыFord PPT MainNishantОценок пока нет
- KTM Canada Weekly Race ReportДокумент3 страницыKTM Canada Weekly Race Reportapi-26009013Оценок пока нет
- Datasheet Kent C 0126 PDFДокумент2 страницыDatasheet Kent C 0126 PDFFWEFWEFWОценок пока нет
- KARATE-Compressed ListingДокумент5 страницKARATE-Compressed Listingburtboyer_moyo928Оценок пока нет