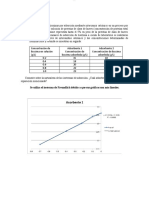
Вам также может понравиться
- Balance de Biorreactor Por Lote AlimentadoДокумент1 страницаBalance de Biorreactor Por Lote AlimentadoLuz RoldanОценок пока нет
- T2 Prob Int M PDFДокумент3 страницыT2 Prob Int M PDFMariana MillanОценок пока нет
- Ejercicio de Trans. Materia en Un BiorreactorДокумент7 страницEjercicio de Trans. Materia en Un BiorreactorKetty Roxana GTerrones0% (1)
- Mètodo Del SulfitoДокумент6 страницMètodo Del SulfitoAllan PgОценок пока нет
- Modelos Cinéticos No Estructurados - No SegregadosДокумент4 страницыModelos Cinéticos No Estructurados - No SegregadosTanilvalОценок пока нет
- Balance Materia en Sistemas de CultivoДокумент19 страницBalance Materia en Sistemas de CultivoWilfredo TullumeОценок пока нет
- Reporte AireaciónДокумент2 страницыReporte AireaciónJenniferОценок пока нет
- 1.3 Estimación de La Difusividad para Sistemas Binarios Gaseosos y LíquidosДокумент5 страниц1.3 Estimación de La Difusividad para Sistemas Binarios Gaseosos y LíquidosRey AgamaОценок пока нет
- Laboratorio 5d - Estequiometría Del Crecimiento y BalancesДокумент44 страницыLaboratorio 5d - Estequiometría Del Crecimiento y Balancesharold olivari liОценок пока нет
- Resumen Balance de Materia y Energía en BiorreactoresДокумент16 страницResumen Balance de Materia y Energía en BiorreactoresLuis AngelОценок пока нет
- Determinación de La VTO (Velocidad de Transferencia de Oxígeno)Документ28 страницDeterminación de La VTO (Velocidad de Transferencia de Oxígeno)Yuki LozanoОценок пока нет
- Semana 2 - Balance de Energia - ProblemasДокумент21 страницаSemana 2 - Balance de Energia - ProblemasGuillermo Saldaña TorresОценок пока нет
- Consumo de Sustrato. Consumo de Sustrato para Crecimiento.Документ5 страницConsumo de Sustrato. Consumo de Sustrato para Crecimiento.Alex TeranОценок пока нет
- Biorreactor de Bucle-Elevacion Por AireДокумент4 страницыBiorreactor de Bucle-Elevacion Por AireGimena Caipo Infantes100% (1)
- Cuadro Comparativo Tipos de Biorreactores - MIGUEL EFRAIN TERRON MEJIA - 1618IBT019 - CONROL DE BIOPROCESOS PDFДокумент4 страницыCuadro Comparativo Tipos de Biorreactores - MIGUEL EFRAIN TERRON MEJIA - 1618IBT019 - CONROL DE BIOPROCESOS PDFSatilab LaboratoriosОценок пока нет
- Capitulo CuatroДокумент82 страницыCapitulo CuatroLéo AyolОценок пока нет
- Ejercicio Extracción Liquido LiquidoДокумент5 страницEjercicio Extracción Liquido LiquidoLina Maria SuarezОценок пока нет
- 1 Estequiometría Del Crecimiento y Balances PDFДокумент59 страниц1 Estequiometría Del Crecimiento y Balances PDFAntonio de la CruzОценок пока нет
- A 3.3 Métodos de Medición Del Kla y Ley de HenryДокумент6 страницA 3.3 Métodos de Medición Del Kla y Ley de HenryKarla TorresОценок пока нет
- Actividad Previa Practica 1 CentrifugaciónДокумент3 страницыActividad Previa Practica 1 CentrifugaciónMattОценок пока нет
- Cuestionario 1-9Документ6 страницCuestionario 1-9Joel Machaca G100% (3)
- Laboratorio Integral 3. - Práctica 9. - Extracción Sólido-LíquidoДокумент9 страницLaboratorio Integral 3. - Práctica 9. - Extracción Sólido-LíquidoDavidHerreraОценок пока нет
- AdsorcionДокумент4 страницыAdsorcionJanelly TejedaОценок пока нет
- Principios de Operación y Aplicaciones de Un Cultivo Por Lote AlimentadoДокумент5 страницPrincipios de Operación y Aplicaciones de Un Cultivo Por Lote AlimentadoIrma Martinez ValenzuelaОценок пока нет
- ExamenДокумент8 страницExamenSalvador Morales CarreonОценок пока нет
- Práctica VTOДокумент28 страницPráctica VTOHector EstrellaОценок пока нет
- TM I Corte 1Документ18 страницTM I Corte 1Laura Camargo Vega100% (1)
- Taller2 2sem2013Документ9 страницTaller2 2sem2013jose davidОценок пока нет
- CromatografíaДокумент6 страницCromatografíaJose F Moo NaalОценок пока нет
- 7-3 7-4 Ocon TojoДокумент2 страницы7-3 7-4 Ocon TojoMiguel FloresОценок пока нет
- Escalabilidad en Procesos BiotecnologicosДокумент25 страницEscalabilidad en Procesos BiotecnologicosLeonardo Lenin Banegas BarahonaОценок пока нет
- Manual de Laboratorio de BioseparacionesДокумент102 страницыManual de Laboratorio de BioseparacionesIrving Toloache FloresОценок пока нет
- Servicios Auxiliares en Un BiorreactorДокумент41 страницаServicios Auxiliares en Un BiorreactorJasminPonce50% (2)
- Práctica #3 Extracción Liquido LíquidoДокумент34 страницыPráctica #3 Extracción Liquido LíquidoAnonymous nTFar7lb67% (3)
- Casos Especiales de Rectificación Usando El Método de McCabeДокумент4 страницыCasos Especiales de Rectificación Usando El Método de McCabeJorge Luis BarriosОценок пока нет
- PRACTICA 9 Extracción Sólido Líquido en Tanque Agitado (INTRODUCCIÓN)Документ14 страницPRACTICA 9 Extracción Sólido Líquido en Tanque Agitado (INTRODUCCIÓN)Guillermo ReyesОценок пока нет
- Biotecnología Agroindustrial IДокумент14 страницBiotecnología Agroindustrial IMauricio Esteban Mosquera JordánОценок пока нет
- Práctica 6 L L 2Документ7 страницPráctica 6 L L 2Bryan Antonio Maza CordovaОценок пока нет
- Técnicas Empleadas para La Conservación de MicrorganismosДокумент7 страницTécnicas Empleadas para La Conservación de MicrorganismosCarolina Campos PeraltaОценок пока нет
- Unidad 3 Control y Regulación MetabólicaДокумент191 страницаUnidad 3 Control y Regulación MetabólicaElva AvalosОценок пока нет
- ProblemasДокумент2 страницыProblemasjovanaОценок пока нет
- Métodos para Medir El KLa - PДокумент32 страницыMétodos para Medir El KLa - PMariana PizanoОценок пока нет
- Sistemas de Cultivo CelularДокумент38 страницSistemas de Cultivo CelularGeorgia Maria González MezaОценок пока нет
- Practica Reactor EnzimaticoДокумент12 страницPractica Reactor EnzimaticoErik AragonОценок пока нет
- Problemario de Bioseparaciones 1Документ7 страницProblemario de Bioseparaciones 1Lydia EniithОценок пока нет
- 1.6 Destilación Con Rectificación-1Документ17 страниц1.6 Destilación Con Rectificación-1Jitzell Lupita Ruperto GarciaОценок пока нет
- Apuntes de Biorreactores PDFДокумент112 страницApuntes de Biorreactores PDFIngeniería QuímicaОценок пока нет
- Biorreactor AirliftДокумент60 страницBiorreactor AirliftAlexis GonzalezОценок пока нет
- Crecimiento Microbiano y Cálculo de RendimientosДокумент18 страницCrecimiento Microbiano y Cálculo de RendimientosCorazon Contento100% (2)
- 4 Procesos Continuos - Criterios de EstabilidadДокумент22 страницы4 Procesos Continuos - Criterios de EstabilidadRuben Dario Marquez FernandezОценок пока нет
- Balances de Materia en Un BiorreactorДокумент10 страницBalances de Materia en Un BiorreactorValeria Nicole Cabrera RomeroОценок пока нет
- Unidad 3 Ingeniería de BiorreactoresДокумент49 страницUnidad 3 Ingeniería de BiorreactoresErick Javier Hernandez Prudencio50% (2)
- Practica 3 Determinación Del Coeficiente de TransferДокумент14 страницPractica 3 Determinación Del Coeficiente de TransferFernando CardonaОценок пока нет
- Cultivos Semicontinuos - P2.3Документ19 страницCultivos Semicontinuos - P2.3Juan Carlos Sasintuña0% (1)
- Balances en BioprocesosДокумент9 страницBalances en BioprocesosMafe BotiaОценок пока нет
- Practica 1 (Completa) - DifusionДокумент22 страницыPractica 1 (Completa) - DifusionLlovll Maywood100% (3)
- Resumen. 8Документ9 страницResumen. 8MayteОценок пока нет
- Cap7 Mi BiorectДокумент13 страницCap7 Mi BiorectErika Lopez ArandaОценок пока нет
- Principios Basicos Del Diseno de BiorreaДокумент32 страницыPrincipios Basicos Del Diseno de BiorreaElvia Alva RojasОценок пока нет
- Para Mejorar El Rendimiento y La Productividad de La Producción de MetabolitosДокумент20 страницPara Mejorar El Rendimiento y La Productividad de La Producción de MetabolitosLuis Enrique Melquiades JimenezОценок пока нет
- Cos Desvs P 01 M 01 Ac 05 F 08Документ4 страницыCos Desvs P 01 M 01 Ac 05 F 08Fender StratОценок пока нет
- Cos Desvs P 01 M 01 Ac 05 F 01Документ8 страницCos Desvs P 01 M 01 Ac 05 F 01Fender StratОценок пока нет
- Cos Desvs P 01 M 01 Ac 05 F 02 PDFДокумент6 страницCos Desvs P 01 M 01 Ac 05 F 02 PDFFender StratОценок пока нет
- Pulsar - NS 150 & As 150 - Users Guide - En.esДокумент35 страницPulsar - NS 150 & As 150 - Users Guide - En.esFender Strat100% (1)
- Cos Desvs P 01 M 01 Ac 05Документ19 страницCos Desvs P 01 M 01 Ac 05Fender StratОценок пока нет
- Transfer de Oxigen oДокумент138 страницTransfer de Oxigen oFender StratОценок пока нет
- Validación de ProcesosДокумент45 страницValidación de ProcesosFender StratОценок пока нет
- DOF - Diario Oficial de La Federación 059 PDFДокумент55 страницDOF - Diario Oficial de La Federación 059 PDFFender StratОценок пока нет
- 164 Ssa 1Документ41 страница164 Ssa 1DrayDergonОценок пока нет
- Capítulo 1 CineticaMicrobianaДокумент22 страницыCapítulo 1 CineticaMicrobianaMau TeissierОценок пока нет
- EmulsionesДокумент8 страницEmulsionesFender StratОценок пока нет
- Examen Muestra Area 1 2014Документ32 страницыExamen Muestra Area 1 2014Dantes InfiernoОценок пока нет
- Guia ValidacionДокумент55 страницGuia ValidacionFender Strat100% (1)
- Control de Calidad en La Industria FarmaceuticaДокумент15 страницControl de Calidad en La Industria FarmaceuticaNicole KnightОценок пока нет
- Curso de CantoДокумент13 страницCurso de Cantoinserasteljerez97% (200)
- Preguntas de BiologíaДокумент4 страницыPreguntas de BiologíaHector Eduardo Coutiño HernandezОценок пока нет
- Anatomía VegetalДокумент3 страницыAnatomía Vegetalsteven chicaОценок пока нет
- Cambios en Tu Hijo Adolescente - Roberto FontanarrosaДокумент5 страницCambios en Tu Hijo Adolescente - Roberto FontanarrosaSebastian CordobaОценок пока нет
- Ingreso Eutm 2020Документ3 страницыIngreso Eutm 2020Mauricio VianaОценок пока нет
- Cuestionario Semana 1 Gabriel ObandoДокумент3 страницыCuestionario Semana 1 Gabriel Obandogabriel obandoОценок пока нет
- Fisiologia Del Aparato Reproductor FemeninoДокумент10 страницFisiologia Del Aparato Reproductor Femeninonatalia201812Оценок пока нет
- Movimientos ArticularesДокумент29 страницMovimientos ArticularesCerrado CerradОценок пока нет
- Mensaj Quim - Hormonas 2020bДокумент41 страницаMensaj Quim - Hormonas 2020balejandroescobar264Оценок пока нет
- Histología de La Placenta (Autoaprendizaje)Документ8 страницHistología de La Placenta (Autoaprendizaje)Xochilth MairenaОценок пока нет
- Sistema UrinarioДокумент10 страницSistema UrinarioAuroraDiaz100% (1)
- Alicorp Apuesta Por La Innovación para Incrementar Su Participación de MercadoДокумент1 страницаAlicorp Apuesta Por La Innovación para Incrementar Su Participación de MercadoMirilly Maile Centurion TorresОценок пока нет
- LimerenciaДокумент4 страницыLimerenciaManny Del Septimo ZoodyackoОценок пока нет
- Expo de Bioquimica ClasificacionДокумент36 страницExpo de Bioquimica ClasificacionGraciela Anabel Castro ChumbeОценок пока нет
- Exposicion de Los Lipidos y VitaminasДокумент47 страницExposicion de Los Lipidos y VitaminasMarily Chanta MontalvoОценок пока нет
- Ejemplo. 1Документ10 страницEjemplo. 1Kelvin GasparОценок пока нет
- Tema 01. Tejidos Vegetales Grado 6° - Diapositivas 1Документ12 страницTema 01. Tejidos Vegetales Grado 6° - Diapositivas 1julianaОценок пока нет
- PUBERTAD PRECOZ y RETARDADAДокумент28 страницPUBERTAD PRECOZ y RETARDADAGinecologa Dra MariaElena Martinez Silva50% (2)
- Esclerodermia y EMTCДокумент15 страницEsclerodermia y EMTCdoctor_carmonaОценок пока нет
- Denise A. Agnew - Corazones OcultosДокумент26 страницDenise A. Agnew - Corazones OcultosMew Rincone100% (1)
- Actividad 7 - ENTREGA SISTEMA NERVIOSO E INMUNITARIOДокумент66 страницActividad 7 - ENTREGA SISTEMA NERVIOSO E INMUNITARIOnathalia mora100% (1)
- Principios de La Lógica Molecular y Jerarquía de La OrganizacionДокумент28 страницPrincipios de La Lógica Molecular y Jerarquía de La OrganizacionAlexanderSKateОценок пока нет
- Analisis de Puesto de Trabajo Al MalacateroДокумент12 страницAnalisis de Puesto de Trabajo Al MalacaterocamiloОценок пока нет
- Trabajo Final de Anatomia-Resumen de SNДокумент211 страницTrabajo Final de Anatomia-Resumen de SNJoseph Ibáñez0% (1)
- 2006 Líquido Pulmonar FetalДокумент5 страниц2006 Líquido Pulmonar FetalWilDe MendozaОценок пока нет
- Manual de Laboratorio de Anatomia y Fisiología Humana 150917Документ36 страницManual de Laboratorio de Anatomia y Fisiología Humana 150917Kaary Lizaama100% (2)
- Medicina Interna PediatricaДокумент137 страницMedicina Interna PediatricaMaryuris Parra100% (1)
- PK Clase 4Документ20 страницPK Clase 4Herflab Cia LtdaОценок пока нет
- Rce49 Desarrollo Motriz de Los 0 Meses A Los 2 AñosДокумент7 страницRce49 Desarrollo Motriz de Los 0 Meses A Los 2 AñosCamila LlanosОценок пока нет
- Glosario Morfofisiologia.Документ2 страницыGlosario Morfofisiologia.Alejandro AmayaОценок пока нет