Вам также может понравиться
- ACLS Full Lesson Plans PDFДокумент60 страницACLS Full Lesson Plans PDFVictoriano Mendez100% (1)
- Bleak House - Book ThreeДокумент36 страницBleak House - Book ThreeNicklaus Adam Rhodes100% (1)
- SDG Business Reporting Analysis 2022Документ559 страницSDG Business Reporting Analysis 2022ComunicarSe-ArchivoОценок пока нет
- Anatomy TerminologyДокумент20 страницAnatomy Terminologykhabbab hussainОценок пока нет
- Concise Guide To Forgetting How Much You SuckДокумент7 страницConcise Guide To Forgetting How Much You SuckihatemakingusernamesОценок пока нет
- Cases and Doctrines On Legal MedicineДокумент44 страницыCases and Doctrines On Legal MedicineEliza MontemayorОценок пока нет
- Leaky Gut TreatmentДокумент5 страницLeaky Gut Treatmentfsilassie8012Оценок пока нет
- Clinical Assessment of The Autonomic Nervous System PDFДокумент312 страницClinical Assessment of The Autonomic Nervous System PDFAndrija100% (1)
- Hydra Facial Machine PDFДокумент11 страницHydra Facial Machine PDFAarifОценок пока нет
- A Handbook of Oral Physiology and Oral BiologyДокумент87 страницA Handbook of Oral Physiology and Oral Biologyamirmaafi100% (1)
- Indrabasti Marma2Документ4 страницыIndrabasti Marma2Rajiv SharmaОценок пока нет
- MSDS ThievesДокумент11 страницMSDS ThievesATOMY KESEHATANОценок пока нет
- Cell Parts and Its FunctionsДокумент25 страницCell Parts and Its FunctionsSen Armario100% (1)
- The Peripheral Vesicles Gather Multivesicular Bodies With Different BehaviorДокумент11 страницThe Peripheral Vesicles Gather Multivesicular Bodies With Different BehaviorUNIG Aluana SantanaОценок пока нет
- Engineering Exosomes For Targeted Drug DeliveryДокумент13 страницEngineering Exosomes For Targeted Drug DeliveryAlexa ReinkeОценок пока нет
- J of Extracellular Bio - 2022 - Ramos - The Functional Role of Soluble Proteins Acquired by Extracellular VesiclesДокумент21 страницаJ of Extracellular Bio - 2022 - Ramos - The Functional Role of Soluble Proteins Acquired by Extracellular VesiclesValen EstevezОценок пока нет
- Extracellular Vesicles Raposo2013Документ11 страницExtracellular Vesicles Raposo2013Karina Pilicita TipánОценок пока нет
- Integrating Cells Into Tissues: OutlineДокумент71 страницаIntegrating Cells Into Tissues: OutlineEvelin Cornejo FundoraОценок пока нет
- All Epithelial Cells in Contact With Subjacent Connective Tissue Have at Their Basal Surfaces A FeltДокумент8 страницAll Epithelial Cells in Contact With Subjacent Connective Tissue Have at Their Basal Surfaces A FeltMegan LewisОценок пока нет
- Cell - The Unit of LifeДокумент26 страницCell - The Unit of LifeKunal MahadikОценок пока нет
- The Role of Extracellular Vesicles in Malaria Biology and PathogenesisДокумент11 страницThe Role of Extracellular Vesicles in Malaria Biology and PathogenesisBruno Moreira SoaresОценок пока нет
- A Gonzalez18originДокумент26 страницA Gonzalez18originRara IndiraОценок пока нет
- Memoranda: Classification of Macrophages, CellsДокумент8 страницMemoranda: Classification of Macrophages, CellsRusu LauraОценок пока нет
- Raport 2Документ9 страницRaport 2shko noshaОценок пока нет
- Chapter 2 Student Copy ZoologyДокумент5 страницChapter 2 Student Copy Zoologyapi-523871804Оценок пока нет
- 0rder 429 Anatomy-Chapter 3 AssignmentДокумент10 страниц0rder 429 Anatomy-Chapter 3 Assignmentjoshua chegeОценок пока нет
- S.L. Kuznetsov, T.V. Boronikhina, V.L. GoryachkinaДокумент106 страницS.L. Kuznetsov, T.V. Boronikhina, V.L. GoryachkinaMustafa MohamedОценок пока нет
- Kalinina Protistology 12-1Документ9 страницKalinina Protistology 12-1jesus javier hoyos acuñaОценок пока нет
- Accepted Manuscript: Genomics, Proteomics & BioinformaticsДокумент25 страницAccepted Manuscript: Genomics, Proteomics & BioinformaticsromanОценок пока нет
- The Journal of Pathology - 2023 - Gregory - Extracellular Vesicles Arising From Apoptosis Forms Functions andДокумент17 страницThe Journal of Pathology - 2023 - Gregory - Extracellular Vesicles Arising From Apoptosis Forms Functions andcoutinhobvictorОценок пока нет
- Midterm Exam in Cell BiologyДокумент5 страницMidterm Exam in Cell BiologyRetchel Cadayong NavalesОценок пока нет
- Extracellular VesicleДокумент14 страницExtracellular VesicleLucas Monteiro LopesОценок пока нет
- MorphogenesisДокумент25 страницMorphogenesisDrAbhilasha SharmaОценок пока нет
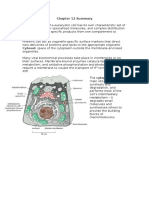
- Chapter 12 SummaryДокумент7 страницChapter 12 SummaryCharlotteОценок пока нет
- Xüsusi Histologiya Dərslik 2015-1Документ14 страницXüsusi Histologiya Dərslik 2015-1aydinmemmedov9009Оценок пока нет
- Sundararajan2018 Article TheVersatileRoleOfExosomesInCaДокумент30 страницSundararajan2018 Article TheVersatileRoleOfExosomesInCaViviana OrellanaОценок пока нет
- Molecular Basis of Morphogenesis During Vertebrate GastrulationДокумент11 страницMolecular Basis of Morphogenesis During Vertebrate GastrulationVero Claros TapiaОценок пока нет
- Histology (Epithelial Tissue)Документ23 страницыHistology (Epithelial Tissue)FazilaОценок пока нет
- Topological Progression in Proliferating Epithelia Is Driven by A Unique Variation in Polygon DistributionДокумент8 страницTopological Progression in Proliferating Epithelia Is Driven by A Unique Variation in Polygon DistributionAuriОценок пока нет
- Cell and Its StructureДокумент70 страницCell and Its StructureEDLYN BRAZILОценок пока нет
- Exosomes: Biogenesis, Biologic Function and Clinical PotentialДокумент18 страницExosomes: Biogenesis, Biologic Function and Clinical PotentialdОценок пока нет
- Perspectives: Extracellular Vesicles: Biology and Emerging Therapeutic OpportunitiesДокумент11 страницPerspectives: Extracellular Vesicles: Biology and Emerging Therapeutic OpportunitiesYanan ZhangОценок пока нет
- The CellДокумент5 страницThe CellAfida Razuna AveОценок пока нет
- Lesson Plan in Cell BiologyДокумент4 страницыLesson Plan in Cell Biologyjahp_13625% (4)
- Histology Text, Problems, TestsДокумент334 страницыHistology Text, Problems, TestsmoahedmaedeОценок пока нет
- Genetics AnswersДокумент8 страницGenetics AnswersLasti DirksawОценок пока нет
- Options For Histological Study of The Structure and Ultrastructure of Human Urinary Bladder EpitheliumДокумент8 страницOptions For Histological Study of The Structure and Ultrastructure of Human Urinary Bladder EpitheliumSergio CerveraОценок пока нет
- Exosome Biogenesis Bioactivities and Functions As New de 2018 BiotechnologyДокумент7 страницExosome Biogenesis Bioactivities and Functions As New de 2018 BiotechnologyMally JamesОценок пока нет
- Mitosis and MeiosisДокумент45 страницMitosis and MeiosisJhon dave SurbanoОценок пока нет
- CELL Study MaterialДокумент12 страницCELL Study MaterialShyamasree SenguptaОценок пока нет
- Tmp4e28 TMPДокумент11 страницTmp4e28 TMPFrontiersОценок пока нет
- Written Assignment 3Документ5 страницWritten Assignment 3Naeem AkhtarОценок пока нет
- 5553 FullДокумент13 страниц5553 FullValen EstevezОценок пока нет
- Desarrollo Del CloroplastoДокумент21 страницаDesarrollo Del CloroplastoAnderson HernándezОценок пока нет
- Chapter 2 Student Copy ZoologyДокумент5 страницChapter 2 Student Copy Zoologyapi-530699294Оценок пока нет
- The CellДокумент20 страницThe CellganeshchandraroutrayОценок пока нет
- JCB 202102001Документ15 страницJCB 202102001Apollo ApolloОценок пока нет
- Advances in Early Kidney Specification, Development and PatterningДокумент12 страницAdvances in Early Kidney Specification, Development and PatterningasdОценок пока нет
- Field JCellBiol 2011Документ10 страницField JCellBiol 2011huouinkyoumaОценок пока нет
- Nucleic Acids in Exosomes: Disease Markers and Intercellular Communication MoleculesДокумент7 страницNucleic Acids in Exosomes: Disease Markers and Intercellular Communication MoleculesGlauce L TrevisanОценок пока нет
- Crozatier Et Al-2007-Cellular MicrobiologyДокумент10 страницCrozatier Et Al-2007-Cellular MicrobiologyVINEET KUREELОценок пока нет
- Biology Form 5Документ373 страницыBiology Form 5shonotano71Оценок пока нет
- Cells: Transport of Materials Across The Cell MembraneДокумент6 страницCells: Transport of Materials Across The Cell MembraneSara SalmanОценок пока нет
- Definition of Biological Science. Biological Science Is A Branch ofДокумент11 страницDefinition of Biological Science. Biological Science Is A Branch ofAlyssa DianneОценок пока нет
- The Cell and Cellular Metabolism ReproductionДокумент4 страницыThe Cell and Cellular Metabolism ReproductionApple AbriamОценок пока нет
- Full Download Human Anatomy 4th Edition Saladin Solutions ManualДокумент35 страницFull Download Human Anatomy 4th Edition Saladin Solutions Manualvocality.duettoqw603100% (36)
- Lesso N2: Cellular StructureДокумент62 страницыLesso N2: Cellular StructureKristine MoresОценок пока нет
- Important Questions For Cell BiologyДокумент16 страницImportant Questions For Cell BiologyTambudzai RazyОценок пока нет
- Exosomes in Viral Disease: ReviewДокумент12 страницExosomes in Viral Disease: ReviewTaniaОценок пока нет
- Embryology GastrulationДокумент9 страницEmbryology GastrulationRivera Gómez América LucíaОценок пока нет
- 2014 Hjer Wangel Smart Sustainable Cities AAMДокумент17 страниц2014 Hjer Wangel Smart Sustainable Cities AAMKayfi Akram MawlanОценок пока нет
- Revisão Sobre Adoção de TecДокумент18 страницRevisão Sobre Adoção de TecGlauce L TrevisanОценок пока нет
- Data Driven Smart Sustainable Cities of The Future An Evidenc - 2021 - SustainaДокумент23 страницыData Driven Smart Sustainable Cities of The Future An Evidenc - 2021 - SustainaLoan ÁnhОценок пока нет
- Brazil Map Infographics by SlidesgoДокумент34 страницыBrazil Map Infographics by SlidesgomrclashpoeОценок пока нет
- Dam AlasДокумент4 страницыDam AlasGlauce L TrevisanОценок пока нет
- Windows of Opportunities and Technological Innovation in The Brazilian Pharmaceutical IndustryДокумент12 страницWindows of Opportunities and Technological Innovation in The Brazilian Pharmaceutical IndustryGlauce L TrevisanОценок пока нет
- Revisão Sobre Adoção de TecДокумент18 страницRevisão Sobre Adoção de TecGlauce L TrevisanОценок пока нет
- TPB Brasil2Документ9 страницTPB Brasil2Glauce L TrevisanОценок пока нет
- Patent Applications On Representative Sectors of Biotechnology in Brazil: An Analysis of The Last DecadeДокумент12 страницPatent Applications On Representative Sectors of Biotechnology in Brazil: An Analysis of The Last DecadeGlauce L TrevisanОценок пока нет
- Brazil Biotechnology Patent Sequences: AC C Y IPL AДокумент2 страницыBrazil Biotechnology Patent Sequences: AC C Y IPL AGlauce L TrevisanОценок пока нет
- CG 14 11 PDFДокумент14 страницCG 14 11 PDFGlauce L TrevisanОценок пока нет
- Brazil Map 2011: BiotechДокумент40 страницBrazil Map 2011: BiotechGlauce L TrevisanОценок пока нет
- Topic ReviewДокумент9 страницTopic ReviewGlauce L TrevisanОценок пока нет
- Advancing Evidence Based Indication Prioritization To Navigate The Segmentation of The Disease LandscapeДокумент7 страницAdvancing Evidence Based Indication Prioritization To Navigate The Segmentation of The Disease LandscapeGlauce L TrevisanОценок пока нет
- An Example of The Utility of Genomic Analysis For Fast and Accurate Clinical Diagnosis of Complex Rare PhenotypesДокумент8 страницAn Example of The Utility of Genomic Analysis For Fast and Accurate Clinical Diagnosis of Complex Rare PhenotypesGlauce L TrevisanОценок пока нет
- Management of Intellectual Property in Brazilian Universities: A Multiple Case StudyДокумент28 страницManagement of Intellectual Property in Brazilian Universities: A Multiple Case StudyGlauce L TrevisanОценок пока нет
- 2016 BIO SubmissionДокумент68 страниц2016 BIO SubmissionGlauce L TrevisanОценок пока нет
- Navigating The Molecular Diagnostic Patent Landscape: Dianne NicolДокумент41 страницаNavigating The Molecular Diagnostic Patent Landscape: Dianne NicolGlauce L TrevisanОценок пока нет
- 080116001Документ49 страниц080116001Glauce L TrevisanОценок пока нет
- Review Article: Evolution of Genetic Techniques: Past, Present, and BeyondДокумент8 страницReview Article: Evolution of Genetic Techniques: Past, Present, and BeyondGlauce L TrevisanОценок пока нет
- Synthetic and Systems Biotechnology: Rosalind A. Le Feuvre, Nigel S. ScruttonДокумент8 страницSynthetic and Systems Biotechnology: Rosalind A. Le Feuvre, Nigel S. ScruttonGlauce L TrevisanОценок пока нет
- 158 Full PDFДокумент10 страниц158 Full PDFGlauce L TrevisanОценок пока нет
- 2014 PAYD BrochureДокумент9 страниц2014 PAYD BrochureGlauce L TrevisanОценок пока нет
- Patent Incentives, Technology Markets, and Public-Private Bio-Medical Innovation Networks in BrazilДокумент12 страницPatent Incentives, Technology Markets, and Public-Private Bio-Medical Innovation Networks in BrazilGlauce L TrevisanОценок пока нет
- Gaia Bernstein AccommodatinДокумент77 страницGaia Bernstein AccommodatinGlauce L TrevisanОценок пока нет
- CG 14 11Документ10 страницCG 14 11Glauce L TrevisanОценок пока нет
- Development and Characterization of Genetic Sensors and RegulatorДокумент197 страницDevelopment and Characterization of Genetic Sensors and RegulatorGlauce L TrevisanОценок пока нет
- Ctga2016 Lec02 PDFДокумент50 страницCtga2016 Lec02 PDFGlauce L TrevisanОценок пока нет
- Gaia Bernstein AccommodatinДокумент77 страницGaia Bernstein AccommodatinGlauce L TrevisanОценок пока нет
- Development and Characterization of Genetic Sensors and RegulatorДокумент197 страницDevelopment and Characterization of Genetic Sensors and RegulatorGlauce L TrevisanОценок пока нет
- Antifertility DrugsДокумент12 страницAntifertility DrugsforplancessОценок пока нет
- CDC Org ChartДокумент1 страницаCDC Org Chartwelcome martinОценок пока нет
- TAPSE AgainДокумент8 страницTAPSE Againomotola ajayiОценок пока нет
- An Overview of Mechanical Ventilation in The Intensive Care UnitДокумент9 страницAn Overview of Mechanical Ventilation in The Intensive Care UnitJaime Javier VegaОценок пока нет
- Radiographic InterpretationДокумент46 страницRadiographic InterpretationSanaFatimaОценок пока нет
- 4 - Chapter - Respiratory SystemДокумент25 страниц4 - Chapter - Respiratory SystemleonОценок пока нет
- Children 07 00162Документ5 страницChildren 07 00162Camille MalilayОценок пока нет
- Nguyen 2019Документ6 страницNguyen 2019ClintonОценок пока нет
- Ratio: Number Rate of Events, Items, Persons, Etc One Group Number Rate of Events, Items, Persons, Etc Another GroupДокумент11 страницRatio: Number Rate of Events, Items, Persons, Etc One Group Number Rate of Events, Items, Persons, Etc Another GroupdayafterОценок пока нет
- Family Health Survey ToolДокумент7 страницFamily Health Survey ToolReniella HidalgoОценок пока нет
- Clinical Biochemical Investigations For Inborn Errors of MetabolismДокумент19 страницClinical Biochemical Investigations For Inborn Errors of Metabolismzain and mariamОценок пока нет
- REKAPITULASI PASIEN CA PENIS NewДокумент51 страницаREKAPITULASI PASIEN CA PENIS Newagus sukarnaОценок пока нет
- Orofacial CleftsДокумент19 страницOrofacial CleftsTj AngelaОценок пока нет
- Report 2Документ13 страницReport 2api-3711225Оценок пока нет
- Rubber Dam Isolation For Endodontic Treatment in Difficult Clinical SituationsДокумент8 страницRubber Dam Isolation For Endodontic Treatment in Difficult Clinical SituationsDavid ColonОценок пока нет
- Pre Assessment Mapeh - 9 - 2014 2015Документ3 страницыPre Assessment Mapeh - 9 - 2014 2015Arianne B. Cabañez100% (1)
- Group 19Документ21 страницаGroup 19Mr. ShaikhОценок пока нет
- Prevalence of Cusp of Carabelli in Permanent Teeth in A Group of Dental Student of School of Dentistry at University of SulaimaniДокумент2 страницыPrevalence of Cusp of Carabelli in Permanent Teeth in A Group of Dental Student of School of Dentistry at University of SulaimaniIOSRjournalОценок пока нет
- BCMJ 53 Vol6 Febrile SeizuresДокумент6 страницBCMJ 53 Vol6 Febrile SeizuresdyyyaОценок пока нет