Вам также может понравиться
- Dynamic Correspondance of Rowing To WeighliftingДокумент6 страницDynamic Correspondance of Rowing To WeighliftingSajeel Chaudhry100% (1)
- FontselfДокумент2 страницыFontselfAlexandru VartolomeiОценок пока нет
- The Wisdom of Trauma Booklet - FinalДокумент11 страницThe Wisdom of Trauma Booklet - Finalligiacassola100% (4)
- Darkle Slideshow by SlidesgoДокумент53 страницыDarkle Slideshow by SlidesgoADITI GUPTAОценок пока нет
- Biomechanics of WeightliftingДокумент4 страницыBiomechanics of Weightliftingapi-3589103540% (1)
- Sprinkler Sizing en v1Документ12 страницSprinkler Sizing en v1CristianDumitru0% (1)
- Management Accounting by Cabrera Solution Manual 2011 PDFДокумент3 страницыManagement Accounting by Cabrera Solution Manual 2011 PDFClaudette Clemente100% (1)
- Effects of Different Volume-Equated Resistance Training Loading Strategies On Muscular Adaptations in Well-Trained MenДокумент11 страницEffects of Different Volume-Equated Resistance Training Loading Strategies On Muscular Adaptations in Well-Trained Menjoaoevandro100% (1)
- Protein Before Bedtime Boosts Muscle Mass and Strength GainsДокумент2 страницыProtein Before Bedtime Boosts Muscle Mass and Strength Gainsmehrdad_44Оценок пока нет
- Absolute Experimental PTДокумент5 страницAbsolute Experimental PTIt's Just Me100% (1)
- ACSM DiabetesДокумент12 страницACSM Diabetesjrice003Оценок пока нет
- Jumpstretch FlexibilityДокумент2 страницыJumpstretch Flexibilitymichal900_408346753100% (1)
- Adaptation To Strength TrainingДокумент26 страницAdaptation To Strength TrainingvОценок пока нет
- Kreiger Set Volume BibleДокумент21 страницаKreiger Set Volume Biblechris_blair_1Оценок пока нет
- Author: Phil Campbell: The Key To Longevity - Lengthen Your Telomeres With Sprint 8Документ2 страницыAuthor: Phil Campbell: The Key To Longevity - Lengthen Your Telomeres With Sprint 8Pedro Alberto FontaniveОценок пока нет
- FST 7Документ5 страницFST 7vishnu SomanОценок пока нет
- Building With Concentric Super OverloadДокумент6 страницBuilding With Concentric Super OverloadBill HonakerОценок пока нет
- Dual Factor Hypertrophy Training by Matt ReynoldsДокумент5 страницDual Factor Hypertrophy Training by Matt Reynoldsstormich656Оценок пока нет
- Increasing Lean Mass and Strength - HFT vs. LFTДокумент9 страницIncreasing Lean Mass and Strength - HFT vs. LFTKarn Mamgain100% (1)
- Why Most People Hate Bodybuilding by Ian KelleyДокумент5 страницWhy Most People Hate Bodybuilding by Ian KelleyianbayyОценок пока нет
- Fitness Profit CenterДокумент3 страницыFitness Profit CenterMinard Paul AracanОценок пока нет
- Bill Starr - Hamstring HelpersДокумент5 страницBill Starr - Hamstring HelpersTomSusОценок пока нет
- Build A Savage YokeДокумент6 страницBuild A Savage YokeShawn Parisi100% (1)
- One-Repetition Maximum: Calculating Approximate 1RMДокумент3 страницыOne-Repetition Maximum: Calculating Approximate 1RMkarzinom100% (1)
- Strength and Mass The Ultimate 26 Week Guide To Building Life Changing Strength Muscle and Power 3413587639 1507721668Документ139 страницStrength and Mass The Ultimate 26 Week Guide To Building Life Changing Strength Muscle and Power 3413587639 1507721668koОценок пока нет
- Protien RequirmentsДокумент5 страницProtien RequirmentsSripara KrishnaОценок пока нет
- Hypertrophy Training Volume: Cellular AdaptationsДокумент2 страницыHypertrophy Training Volume: Cellular Adaptationssalva1310Оценок пока нет
- High Intensity Aerobic TrainingДокумент10 страницHigh Intensity Aerobic TrainingdietabiosoficaОценок пока нет
- Monster Supplements Mass GuideДокумент8 страницMonster Supplements Mass GuideNigel ButcherОценок пока нет
- EatingToWin PDFДокумент102 страницыEatingToWin PDFmaurice86Оценок пока нет
- Anjdeep Lad Extensive MedballДокумент4 страницыAnjdeep Lad Extensive MedballKarlosОценок пока нет
- Chapter Training - Intensity - DistributionДокумент12 страницChapter Training - Intensity - Distributionsilvio da costa guerreiroОценок пока нет
- By Mike Westerdal & Ben Tatar - All Rights ReservedДокумент22 страницыBy Mike Westerdal & Ben Tatar - All Rights ReservedThanosShadow100% (1)
- LO5.1: Analyse The 3 Different Energy Systems & Explain Their Contribution To Different Sports & ActivitiesДокумент6 страницLO5.1: Analyse The 3 Different Energy Systems & Explain Their Contribution To Different Sports & Activitiesnbeer93Оценок пока нет
- Nutrition For The SprinterДокумент12 страницNutrition For The SprinterMarcelo NogueraОценок пока нет
- NextLevel ManualДокумент14 страницNextLevel ManualWilson_SC91Оценок пока нет
- Haff NotesДокумент4 страницыHaff NotesTimmОценок пока нет
- WEEK 6 Muscular SystemДокумент17 страницWEEK 6 Muscular SystemMarjorie Lyka LingatОценок пока нет
- Muscle Recovery 4 ElliottДокумент16 страницMuscle Recovery 4 ElliottAäron WajnbergОценок пока нет
- Russian and Eastern European Approach To Training PDFДокумент8 страницRussian and Eastern European Approach To Training PDFFrancisco García González100% (1)
- Muscular TissueДокумент65 страницMuscular TissueMitzel SapaloОценок пока нет
- Eccentric Exercise Program Design - A Periodization Model For Rehabilitation ApplicationsДокумент37 страницEccentric Exercise Program Design - A Periodization Model For Rehabilitation ApplicationsDeivison Fellipe da Silva CâmaraОценок пока нет
- By Lorenzo Pansini: Home: CorrespondenceДокумент24 страницыBy Lorenzo Pansini: Home: CorrespondenceĐạt NguyễnОценок пока нет
- Muscle Fibers - An in Depth Analysis Part 3 AbcbodybuildingДокумент7 страницMuscle Fibers - An in Depth Analysis Part 3 AbcbodybuildingsupiulilumaОценок пока нет
- Biomechanical Analysis of High JumpДокумент15 страницBiomechanical Analysis of High JumpBen Wat100% (1)
- FF Supplement GuideДокумент17 страницFF Supplement GuideAlina Maria Vlaicu-DunoiuОценок пока нет
- A Biomechanical Analysis of The Heavy SPДокумент37 страницA Biomechanical Analysis of The Heavy SPjeyОценок пока нет
- Strength Training and Endurance Performance - ResearchGateДокумент27 страницStrength Training and Endurance Performance - ResearchGateCristianLopezОценок пока нет
- RPE and WeightliftingДокумент2 страницыRPE and Weightliftingwhatguy309Оценок пока нет
- Warm-Up Exercise Quadriceps:: Lower-Body 1 Set of 15 Repetitions On EachДокумент8 страницWarm-Up Exercise Quadriceps:: Lower-Body 1 Set of 15 Repetitions On EachRaymanОценок пока нет
- The Art of Gaining Fifty Pounds of Muscle in Fifty WeeksДокумент14 страницThe Art of Gaining Fifty Pounds of Muscle in Fifty Weeksqbrooks100% (1)
- Operation Bulk UpДокумент52 страницыOperation Bulk UpPhilip Choo100% (2)
- 1997 - Expression and Development of Maximal Muscle PowerДокумент262 страницы1997 - Expression and Development of Maximal Muscle PowerDonato FormicolaОценок пока нет
- Science of Weight ControlДокумент14 страницScience of Weight ControlWes GearyОценок пока нет
- 327616blood Flow Restriction (BFR) Training - Sports Medicine Center ...Документ2 страницы327616blood Flow Restriction (BFR) Training - Sports Medicine Center ...tyrelawlknОценок пока нет
- Eccentric or Concentric Exercises For The Treatment of TendinopathiesДокумент11 страницEccentric or Concentric Exercises For The Treatment of TendinopathiesAnonymous xvlg4m5xLXОценок пока нет
- Categories of Weight Training Part 4Документ2 страницыCategories of Weight Training Part 4Jose Luis Bonilla TОценок пока нет
- Exercise IntensityДокумент5 страницExercise Intensitymr_n_manОценок пока нет
- Https WWW Scribd Com Document 405096854 Ultimate Shoulders Vol 2 Ella New Physique Look PDFДокумент5 страницHttps WWW Scribd Com Document 405096854 Ultimate Shoulders Vol 2 Ella New Physique Look PDFkim gekОценок пока нет
- How Much Can You LiftДокумент6 страницHow Much Can You LiftaboubakereldjazairiОценок пока нет
- Stage 4 - Forced Adaptation: The Quick Guide to Professional Level LiftingОт EverandStage 4 - Forced Adaptation: The Quick Guide to Professional Level LiftingОценок пока нет
- Vitalojak User ManualДокумент2 страницыVitalojak User ManualAlexandru VartolomeiОценок пока нет
- User's Manual: Flash Glucose Monitoring SystemДокумент125 страницUser's Manual: Flash Glucose Monitoring SystemAlexandru VartolomeiОценок пока нет
- AdvisorPro BrochureДокумент4 страницыAdvisorPro BrochureAlexandru VartolomeiОценок пока нет
- Immunity Boost ChecklistДокумент2 страницыImmunity Boost ChecklistAlexandru VartolomeiОценок пока нет
- Rolul Obiceiurilor Alimentare in Sindromul CoronarianДокумент8 страницRolul Obiceiurilor Alimentare in Sindromul CoronarianAlexandru VartolomeiОценок пока нет
- Bomb Defusal ManualДокумент23 страницыBomb Defusal ManualcontactzumorioОценок пока нет
- Low Frequency ElectroStimulationДокумент7 страницLow Frequency ElectroStimulationAlexandru VartolomeiОценок пока нет
- Nutritia in Ziua CompetitieiДокумент2 страницыNutritia in Ziua CompetitieiAlexandru VartolomeiОценок пока нет
- Swimming NutritionДокумент8 страницSwimming NutritionAlexandru Vartolomei100% (1)
- Lyle McDonald Generic Bulking Routine FAQДокумент28 страницLyle McDonald Generic Bulking Routine FAQockhamsrazor2100% (2)
- Consumul de Lapte Degresat Promoveaza Cresterea MuscularaДокумент21 страницаConsumul de Lapte Degresat Promoveaza Cresterea MuscularaAlexandru VartolomeiОценок пока нет
- Aderența La Tratamentul ObezitatiiДокумент10 страницAderența La Tratamentul ObezitatiiAlexandru VartolomeiОценок пока нет
- How Many Portions of Fruit/vegetables/grains in . G Carbohidrates How Many Portions of Meat in . G Proteins How Many Portions Nuts . G FatДокумент1 страницаHow Many Portions of Fruit/vegetables/grains in . G Carbohidrates How Many Portions of Meat in . G Proteins How Many Portions Nuts . G FatAlexandru VartolomeiОценок пока нет
- Obesity and Mental HealthДокумент28 страницObesity and Mental HealthHv EstokОценок пока нет
- FAO-EnACT Workshop Enact-Training July 2012Документ15 страницFAO-EnACT Workshop Enact-Training July 2012Alexandru VartolomeiОценок пока нет
- Endocrinology - EncyclopediaДокумент2 399 страницEndocrinology - Encyclopediaviobanaruu0% (1)
- PRIME Student Course, Cluj 2013Документ4 страницыPRIME Student Course, Cluj 2013Alexandru VartolomeiОценок пока нет
- Amity School of Business:, Semester IV Research Methodology and Report Preparation Dr. Deepa KapoorДокумент23 страницыAmity School of Business:, Semester IV Research Methodology and Report Preparation Dr. Deepa KapoorMayank TayalОценок пока нет
- Week 3 Lab Arado, Patrick James M.Документ2 страницыWeek 3 Lab Arado, Patrick James M.Jeffry AradoОценок пока нет
- Keeping Track of Your Time: Keep Track Challenge Welcome GuideДокумент1 страницаKeeping Track of Your Time: Keep Track Challenge Welcome GuideRizky NurdiansyahОценок пока нет
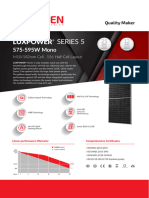
- Ficha Técnica Panel Solar 590W LuxenДокумент2 страницыFicha Técnica Panel Solar 590W LuxenyolmarcfОценок пока нет
- Class 12 Physics Derivations Shobhit NirwanДокумент6 страницClass 12 Physics Derivations Shobhit Nirwanaastha.sawlaniОценок пока нет
- A Project Report ON Strategic Purchasing Procedure, Systems and Policies (Hospital Industry)Документ20 страницA Project Report ON Strategic Purchasing Procedure, Systems and Policies (Hospital Industry)amitwin1983Оценок пока нет
- SSC Gr8 Biotech Q4 Module 1 WK 1 - v.01-CC-released-09May2021Документ22 страницыSSC Gr8 Biotech Q4 Module 1 WK 1 - v.01-CC-released-09May2021Ivy JeanneОценок пока нет
- BECED S4 Motivational Techniques PDFДокумент11 страницBECED S4 Motivational Techniques PDFAmeil OrindayОценок пока нет
- Understanding The Contribution of HRM Bundles For Employee Outcomes Across The Life-SpanДокумент15 страницUnderstanding The Contribution of HRM Bundles For Employee Outcomes Across The Life-SpanPhuong NgoОценок пока нет
- Intelligent Status Monitoring System For Port Machinery: RMGC/RTGCДокумент2 страницыIntelligent Status Monitoring System For Port Machinery: RMGC/RTGCfatsahОценок пока нет
- How Transformers WorkДокумент15 страницHow Transformers Worktim schroderОценок пока нет
- A Short Survey On Memory Based RLДокумент18 страницA Short Survey On Memory Based RLcnt dvsОценок пока нет
- ME-6501Computer Aided Design (CAD) WITH QB - BY Civildatas - Com 1Документ85 страницME-6501Computer Aided Design (CAD) WITH QB - BY Civildatas - Com 1Nathar ShaОценок пока нет
- 7 - Monte-Carlo-Simulation With XL STAT - English GuidelineДокумент8 страниц7 - Monte-Carlo-Simulation With XL STAT - English GuidelineGauravShelkeОценок пока нет
- IPA Smith Osborne21632Документ28 страницIPA Smith Osborne21632johnrobertbilo.bertilloОценок пока нет
- WWW Ranker Com List Best-Isekai-Manga-Recommendations Ranker-AnimeДокумент8 страницWWW Ranker Com List Best-Isekai-Manga-Recommendations Ranker-AnimeDestiny EasonОценок пока нет
- Project Name: Repair of Afam Vi Boiler (HRSG) Evaporator TubesДокумент12 страницProject Name: Repair of Afam Vi Boiler (HRSG) Evaporator TubesLeann WeaverОценок пока нет
- CPD - SampleДокумент3 страницыCPD - SampleLe Anh DungОценок пока нет
- SCD Course List in Sem 2.2020 (FTF or Online) (Updated 02 July 2020)Документ2 страницыSCD Course List in Sem 2.2020 (FTF or Online) (Updated 02 July 2020)Nguyễn Hồng AnhОценок пока нет
- Sony x300 ManualДокумент8 страницSony x300 ManualMarcosCanforaОценок пока нет
- Manhole Head LossesДокумент11 страницManhole Head Lossesjoseph_mscОценок пока нет
- Sensitivity of Rapid Diagnostic Test and Microscopy in Malaria Diagnosis in Iva-Valley Suburb, EnuguДокумент4 страницыSensitivity of Rapid Diagnostic Test and Microscopy in Malaria Diagnosis in Iva-Valley Suburb, EnuguSMA N 1 TOROHОценок пока нет
- Philo Q2 Lesson 5Документ4 страницыPhilo Q2 Lesson 5Julliana Patrice Angeles STEM 11 RUBYОценок пока нет
- Transportation of CementДокумент13 страницTransportation of CementKaustubh Joshi100% (1)
- Point and Figure ChartsДокумент5 страницPoint and Figure ChartsShakti ShivaОценок пока нет
- 12 Step Worksheet With QuestionsДокумент26 страниц12 Step Worksheet With QuestionsKristinDaigleОценок пока нет
- PTW Site Instruction NewДокумент17 страницPTW Site Instruction NewAnonymous JtYvKt5XEОценок пока нет