Вам также может понравиться
- Hand Book of Forest ConservationДокумент131 страницаHand Book of Forest Conservationsouravsahoo1100% (1)
- Ch4 Evolution of Vascular PlantsДокумент112 страницCh4 Evolution of Vascular PlantsGabriel Alfonso D. RobeniolОценок пока нет
- Agrobiodiversity AssessmentДокумент9 страницAgrobiodiversity AssessmentParque Natural Montanhas de TeresópolisОценок пока нет
- PETA Investigates Circus Abuse: Naresh Kadyan Demands FIR Against All Circuses and Cancellation of Registration With AWBI - CZA Along With Rescue of Abused AnimalsДокумент43 страницыPETA Investigates Circus Abuse: Naresh Kadyan Demands FIR Against All Circuses and Cancellation of Registration With AWBI - CZA Along With Rescue of Abused AnimalsAbhishek KadyanОценок пока нет
- Plataspidae Related Cantharodes ReviewДокумент33 страницыPlataspidae Related Cantharodes ReviewanilimОценок пока нет
- Unit 7 PDFДокумент22 страницыUnit 7 PDFAdesh RaoОценок пока нет
- The Soil Mites of the World: Vol. 1: Primitive Oribatids of the Palaearctic RegionОт EverandThe Soil Mites of the World: Vol. 1: Primitive Oribatids of the Palaearctic RegionОценок пока нет
- Module III - Unit 3 - THE NATURE AND COMPOSITION OF PLANTSДокумент10 страницModule III - Unit 3 - THE NATURE AND COMPOSITION OF PLANTSRechel SamontinaОценок пока нет
- Plants Adapt to Environmental Factors Through Morphological, Anatomical and Physiological ChangesДокумент25 страницPlants Adapt to Environmental Factors Through Morphological, Anatomical and Physiological Changessrk2005Оценок пока нет
- Mechanistic Modelling in Pig and Poultry ProductionДокумент343 страницыMechanistic Modelling in Pig and Poultry ProductionEverton OliveiraОценок пока нет
- Dominance of InsectsДокумент30 страницDominance of InsectsA R V Kumar0% (1)
- c3 and c4 PlantДокумент13 страницc3 and c4 PlantWaseem HaiderОценок пока нет
- Protista Written ReportДокумент8 страницProtista Written ReportLance RiveraОценок пока нет
- 15 January 2004 1st Lecture Herpetology ECOL 483/583 University of Arizona Spring 2004Документ14 страниц15 January 2004 1st Lecture Herpetology ECOL 483/583 University of Arizona Spring 2004andreaweiler1Оценок пока нет
- Cell Biology and Genetics ManualДокумент29 страницCell Biology and Genetics ManualAyesha FatimaОценок пока нет
- M1 LN Introduction To Animal WelfareДокумент14 страницM1 LN Introduction To Animal Welfarefsuarez113100% (1)
- Effects of Selected Probiotics On The Growth and Survival of Fry - Fingerlings of Clarias GariepinusДокумент5 страницEffects of Selected Probiotics On The Growth and Survival of Fry - Fingerlings of Clarias GariepinusIOSRjournalОценок пока нет
- Plant Growth Promoting RhizobacteriaДокумент11 страницPlant Growth Promoting RhizobacteriaDiral SadriОценок пока нет
- Biodiversity Conservation and Management For BSC Botany StudentsДокумент41 страницаBiodiversity Conservation and Management For BSC Botany StudentsMadan ThapaОценок пока нет
- Cyanobacteria Economic ImportanceДокумент15 страницCyanobacteria Economic ImportanceKumail IslamianОценок пока нет
- Animal Nutrition ANN 212Документ64 страницыAnimal Nutrition ANN 212mfmalikОценок пока нет
- Methods To Study Degradation of Ruminant FeedsДокумент15 страницMethods To Study Degradation of Ruminant FeedsJ Jesus Bustamante GroОценок пока нет
- Papaya in VitroДокумент5 страницPapaya in VitroBiomontec Biomontec BiomontecОценок пока нет
- Peatland HydrologyДокумент87 страницPeatland HydrologyAfita ainiОценок пока нет
- Artropoda (Insecta)Документ51 страницаArtropoda (Insecta)sila rahma100% (1)
- Ethinobotany Assignment 1finalДокумент16 страницEthinobotany Assignment 1finalGedefaw AlemkereОценок пока нет
- Danh Mục Sơ Bộ Các Loài Bướm Phú Quốc, 2008 Initial Photographic Checklist of Butterfly of PQДокумент24 страницыDanh Mục Sơ Bộ Các Loài Bướm Phú Quốc, 2008 Initial Photographic Checklist of Butterfly of PQChoi Song Dong SomОценок пока нет
- RPB V16n1-Mach PDFДокумент146 страницRPB V16n1-Mach PDFLeonardo Romero0% (2)
- Role of Housefly (Musca Domestica, Diptera Muscidae) AsДокумент5 страницRole of Housefly (Musca Domestica, Diptera Muscidae) AsIqbal Ardiansyah100% (1)
- Recognition of Zoo Rules, 2009 with (Amendment) Rules, 2013Документ24 страницыRecognition of Zoo Rules, 2009 with (Amendment) Rules, 2013salman672003Оценок пока нет
- Restoring Ecology in Alexandria Through Degradation ReductionДокумент31 страницаRestoring Ecology in Alexandria Through Degradation ReductionTmt TarekОценок пока нет
- Lab Algae SimplifiedДокумент21 страницаLab Algae SimplifiedImran AzmanОценок пока нет
- Forest Restoration in LandscapesДокумент440 страницForest Restoration in Landscapesleguim6Оценок пока нет
- Temporal and Spatial Distribution of Common Bacterial Livestock Disease Outbreaks in North Gondar EthiopiaДокумент5 страницTemporal and Spatial Distribution of Common Bacterial Livestock Disease Outbreaks in North Gondar EthiopiaIJARP PublicationsОценок пока нет
- What Determines The Abundance of Lianas and VinesДокумент21 страницаWhat Determines The Abundance of Lianas and VinesccasalvaОценок пока нет
- Fish Cytogenetics, Genotoxicity and Mutagenesis. (5-12-1998)Документ5 страницFish Cytogenetics, Genotoxicity and Mutagenesis. (5-12-1998)Dr.Mrs.Krishnaja A. P.Оценок пока нет
- Identification of Pollen Allelochemical in Hieracium X Dutillyanum Lepage and Its Ecological Impacts On Conyza CanadensisДокумент1 страницаIdentification of Pollen Allelochemical in Hieracium X Dutillyanum Lepage and Its Ecological Impacts On Conyza Canadensisbrkica2011Оценок пока нет
- Ex Situ ConservationДокумент17 страницEx Situ Conservationifa_saimi50% (2)
- Aquatic Animal Nutrition Organic Macro - and Micro-NutrientsДокумент1 082 страницыAquatic Animal Nutrition Organic Macro - and Micro-NutrientsOnur DemirelОценок пока нет
- Plant Seeds and Fruits SamplepagesДокумент40 страницPlant Seeds and Fruits SamplepagesioncacaciosuОценок пока нет
- Home Gardens & in Situ Conservation of Plant Genetic Resources in Farming Systems Gardening GuidebookДокумент192 страницыHome Gardens & in Situ Conservation of Plant Genetic Resources in Farming Systems Gardening GuidebookWinter GardeningОценок пока нет
- Introduction To Plant SystematicsДокумент10 страницIntroduction To Plant SystematicsIzba SanaОценок пока нет
- Mark Kurt Schutze Thesis 2Документ173 страницыMark Kurt Schutze Thesis 2Azhar Ahmad NasriОценок пока нет
- Diversity of Timber Yielding Plants Found in Different Parts of DistrictДокумент7 страницDiversity of Timber Yielding Plants Found in Different Parts of DistrictESSENCE - International Journal for Environmental Rehabilitation and ConservaionОценок пока нет
- PLANT BREEDING - INTRO-2nd Lecture, 2003Документ22 страницыPLANT BREEDING - INTRO-2nd Lecture, 2003lkokodkodОценок пока нет
- David M. JarzenДокумент6 страницDavid M. Jarzen76697669Оценок пока нет
- Micro CH 6 BacteriaДокумент58 страницMicro CH 6 BacteriaBernadette Joyce PascualОценок пока нет
- Finkeldey - Tropical Forest Genetics PDFДокумент318 страницFinkeldey - Tropical Forest Genetics PDFLewisKarl100% (1)
- Ch-1 Nematology IntroductionДокумент6 страницCh-1 Nematology IntroductionHumaNazОценок пока нет
- Applications of Tissue Culture To Plant Improvement: February 2017Документ42 страницыApplications of Tissue Culture To Plant Improvement: February 2017nareman hassanОценок пока нет
- Cultivable Fis, Prawn, Mussels, Crabs and OystersДокумент26 страницCultivable Fis, Prawn, Mussels, Crabs and OystersNarasimha MurthyОценок пока нет
- Chironomus Larvae Culture - A Boon To Aquaculture SectorДокумент13 страницChironomus Larvae Culture - A Boon To Aquaculture SectorInternational Journal of Current Science Research (IJCSR)Оценок пока нет
- Networking To "Blast" Rice Blast From Farmers' FieldsДокумент3 страницыNetworking To "Blast" Rice Blast From Farmers' FieldsIRRI_resourcesОценок пока нет
- The Pea Crop: A Basis for ImprovementОт EverandThe Pea Crop: A Basis for ImprovementP. D. HebblethwaiteОценок пока нет
- Population Ecology Answer KeyДокумент4 страницыPopulation Ecology Answer KeytahamidОценок пока нет
- PollinationДокумент9 страницPollinationMia MadiaОценок пока нет
- Aphid, (Family Aphididae), Also Called Plant Louse, Greenfly, or Ant Cow, Any of A Group of SapДокумент11 страницAphid, (Family Aphididae), Also Called Plant Louse, Greenfly, or Ant Cow, Any of A Group of SapAmjadRashidОценок пока нет
- Animal Form and Function: C E L PДокумент38 страницAnimal Form and Function: C E L PJoshua Miguel Buno0% (1)
- Entomology: Naming of TaxaДокумент19 страницEntomology: Naming of TaxaAliah Alleyne Canals - BSA - 1AОценок пока нет
- Human Body System WorksheetДокумент2 страницыHuman Body System WorksheetRPh Krishna Chandra JagritОценок пока нет
- Global Burden of Bronchial Asthma and Community-Based Management /TITLEДокумент43 страницыGlobal Burden of Bronchial Asthma and Community-Based Management /TITLEMarius-Sorin CionteaОценок пока нет
- Clin Anat 08 DermatomesДокумент11 страницClin Anat 08 DermatomesOchiMataОценок пока нет
- Exam 3 Study Guide - 2017Документ2 страницыExam 3 Study Guide - 2017kerredaiОценок пока нет
- CSCS Practice ExamДокумент15 страницCSCS Practice ExamKhaled Salhi100% (1)
- 5.nephritic SyndromeДокумент64 страницы5.nephritic Syndromeyeni100% (1)
- Lecture 3 - Monod KineticsДокумент19 страницLecture 3 - Monod KineticsZeny Naranjo0% (1)
- Company Most Recent Adposition (S) First Name Last Name Contact TypeДокумент8 страницCompany Most Recent Adposition (S) First Name Last Name Contact TypepaytencasОценок пока нет
- CarbohydratesДокумент41 страницаCarbohydratesStrange Me100% (3)
- LdfısjlısДокумент52 страницыLdfısjlısChristine HudginsОценок пока нет
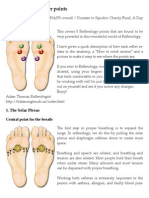
- Reflexology - 5 Power PointsДокумент6 страницReflexology - 5 Power Pointsrickiti9100% (1)
- Potato Poisoning: Understanding Solanine ToxicityДокумент13 страницPotato Poisoning: Understanding Solanine ToxicityDaz Jones100% (2)
- (BOTAFUN) Plant Cell Cycle ControlДокумент3 страницы(BOTAFUN) Plant Cell Cycle ControlBen Joshua MancileОценок пока нет
- Cooperative Activity 3-1 Through 3-3Документ3 страницыCooperative Activity 3-1 Through 3-3Thalia Sanders100% (2)
- Nitrifying Moving Bed Biofilm Reactor (MBBR) Biofilm and Biomass Response To Long Term Exposure To 1 CДокумент10 страницNitrifying Moving Bed Biofilm Reactor (MBBR) Biofilm and Biomass Response To Long Term Exposure To 1 CSyamsul Bahry HarahapОценок пока нет
- Pathophysiology of PainДокумент31 страницаPathophysiology of PainJonniwal Sanusi100% (4)
- OAT Reading Comprehension Practice TestДокумент8 страницOAT Reading Comprehension Practice TestioatprepОценок пока нет
- The Development of Tibiofemoral Angle in Children - Salenius e VankkaДокумент3 страницыThe Development of Tibiofemoral Angle in Children - Salenius e VankkaVictor SechinОценок пока нет
- A Guide To Identifying Snakes by Scalation and Other DetailsДокумент4 страницыA Guide To Identifying Snakes by Scalation and Other DetailsReticulatusОценок пока нет
- Glosarium B.ing BiologiДокумент5 страницGlosarium B.ing BiologiAnita HandayaniОценок пока нет
- CLS Aipmt-17-18 XI Zoo Study-Package-1 SET-1 Chapter-1A PDFДокумент14 страницCLS Aipmt-17-18 XI Zoo Study-Package-1 SET-1 Chapter-1A PDFChunnu BishtОценок пока нет
- 100 Tips To Get Rank-YandamooriДокумент10 страниц100 Tips To Get Rank-Yandamoorijdprasad63% (16)
- CNS Stimulants 2023Документ10 страницCNS Stimulants 2023ManWol JangОценок пока нет
- Mizuho Vascular (Transcranial) DopplerДокумент4 страницыMizuho Vascular (Transcranial) DopplersigmakarsaОценок пока нет
- Free Printable at Home Brain Breaks and Active CalmingДокумент12 страницFree Printable at Home Brain Breaks and Active Calmingapi-505036826Оценок пока нет
- Mod 7 PoisoningДокумент12 страницMod 7 Poisoningrez198767% (3)
- Psychology Prelim Exam 1st Sem 2016-17Документ2 страницыPsychology Prelim Exam 1st Sem 2016-17Monroe Ortizano100% (1)
- Macmillan Science Level 4 Pupil S Book Unit 1 PP 11 16Документ6 страницMacmillan Science Level 4 Pupil S Book Unit 1 PP 11 16reema2050reemaОценок пока нет
- Leaf StructureДокумент10 страницLeaf StructureKristina PonceОценок пока нет
- Botany II PDFДокумент7 страницBotany II PDFMuhammad AmirОценок пока нет