Вам также может понравиться
- Theurgia Pratica CabalisticaДокумент179 страницTheurgia Pratica CabalisticaHeltic BaruchОценок пока нет
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- Ritual of The Invisible GuardiansДокумент7 страницRitual of The Invisible Guardiansenzomontresol100% (1)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- Drug Delivery Systems - An Update ReviewДокумент10 страницDrug Delivery Systems - An Update ReviewenzomontresolОценок пока нет
- Basic Coronary Anatomy - Paul Fefer, MD.Документ57 страницBasic Coronary Anatomy - Paul Fefer, MD.enzomontresolОценок пока нет
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (400)
- Cardiac Cath Lab Basis - Indications, Complications and Radiation ManagementДокумент62 страницыCardiac Cath Lab Basis - Indications, Complications and Radiation ManagementenzomontresolОценок пока нет
- Pediatric Ocular EmergenciesДокумент71 страницаPediatric Ocular EmergenciesenzomontresolОценок пока нет
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- Para-Transes Pre-Final Exam - Unit 4Документ11 страницPara-Transes Pre-Final Exam - Unit 4Aysha AishaОценок пока нет
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- Hollinshead's Functional Anatomy of The Limbs and Back PDFДокумент353 страницыHollinshead's Functional Anatomy of The Limbs and Back PDFCarla S Morales Garcia100% (1)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (895)
- Anemia Nursing Care PlanДокумент4 страницыAnemia Nursing Care PlanMaverick Lim80% (5)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
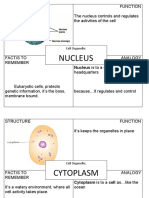
- Nucleus: Structure Function The Nucleus Controls and Regulates The Activities of The CellДокумент13 страницNucleus: Structure Function The Nucleus Controls and Regulates The Activities of The CellAndres ValleОценок пока нет
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- Appendix G Exam Content Outlines and Sample Questions. Certification-Handbook Effective Jan 2020Документ9 страницAppendix G Exam Content Outlines and Sample Questions. Certification-Handbook Effective Jan 2020DannyОценок пока нет
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- Embryogenesis & Fetal DevelopementДокумент38 страницEmbryogenesis & Fetal DevelopementDasha Vee100% (1)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (266)
- 1 - Anatomical Terminology KEYДокумент6 страниц1 - Anatomical Terminology KEY愛.NiniОценок пока нет
- Integration of Metabolism WebДокумент72 страницыIntegration of Metabolism WebLini Anisfatus SholihahОценок пока нет
- Endocrine System and AdolescenceДокумент22 страницыEndocrine System and Adolescencenia tombriОценок пока нет
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- 11th SA-I Bio Merged Paper23-24Документ11 страниц11th SA-I Bio Merged Paper23-24PriyanshuОценок пока нет
- Kinesio Taping 2Документ137 страницKinesio Taping 2abdul haseebОценок пока нет
- Enzymes Mode of Action of EnzymesДокумент6 страницEnzymes Mode of Action of EnzymesNamra MazherОценок пока нет
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (345)
- Andries, 2001, Endocrine and Environmental Control of Reproduction in Polychaeta PDFДокумент17 страницAndries, 2001, Endocrine and Environmental Control of Reproduction in Polychaeta PDFLeonardo ZambranoОценок пока нет
- 653138b8bb9c9f00188ac1f4 - ## - Practice Sheet - 02 - Cell Cycle and Cell DivisionДокумент13 страниц653138b8bb9c9f00188ac1f4 - ## - Practice Sheet - 02 - Cell Cycle and Cell Divisionskrohul7760Оценок пока нет
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2259)
- Neuro Estudio de Casos Del Dolor Neurologico 2021Документ151 страницаNeuro Estudio de Casos Del Dolor Neurologico 2021rogelioОценок пока нет
- Cell AssignmentДокумент3 страницыCell AssignmentDreamy AspirationsОценок пока нет
- Animal KingdomДокумент22 страницыAnimal KingdomAnonymous B2B6lAzeGwОценок пока нет
- 1992 - Rayle, Cleland - The Acid Growth Theory of Auxin-Induced Cell Elongation Is Alive and WellДокумент4 страницы1992 - Rayle, Cleland - The Acid Growth Theory of Auxin-Induced Cell Elongation Is Alive and Wellpond_1993Оценок пока нет
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- Sexual Reproduction in Flowering Plants DPP-01 Discussion Class NotesДокумент4 страницыSexual Reproduction in Flowering Plants DPP-01 Discussion Class NotesTrendy TrendsОценок пока нет
- Biology Mitosis Notes (O Level)Документ9 страницBiology Mitosis Notes (O Level)ojilongОценок пока нет
- Exercise 5. Measuring Carbon Dioxide Production in Animals PDFДокумент3 страницыExercise 5. Measuring Carbon Dioxide Production in Animals PDFJM MelecioОценок пока нет
- HSC Biology Module 5 NotesДокумент15 страницHSC Biology Module 5 Notesarabellatav23Оценок пока нет
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (121)
- 04 30 2023 Scenar Therapy For Tense Muscles Caused by Stress - oДокумент3 страницы04 30 2023 Scenar Therapy For Tense Muscles Caused by Stress - oHora Castro100% (1)
- Temporal Expression of Genes Involved in Folate Metabolism and Transport During Placental Development, Preeclampsia and Neural Tube DefectsДокумент9 страницTemporal Expression of Genes Involved in Folate Metabolism and Transport During Placental Development, Preeclampsia and Neural Tube DefectsMauro Porcel de PeraltaОценок пока нет
- Biochemistry and MetabolismДокумент8 страницBiochemistry and MetabolismPrincess Angie GonzalesОценок пока нет
- Saida Hal 8-18 "Analisis Faktor Risiko Kejadian Hipertensi ... " Jurnal KeperawatanДокумент11 страницSaida Hal 8-18 "Analisis Faktor Risiko Kejadian Hipertensi ... " Jurnal KeperawatanYanti HarjonoОценок пока нет
- Robbins 7th Ed - Chapter 3 - Tissue Renewal & Repair (2) Flashcards by Heli Sinnpson - BrainscapeДокумент12 страницRobbins 7th Ed - Chapter 3 - Tissue Renewal & Repair (2) Flashcards by Heli Sinnpson - BrainscapeCarlos MaingeОценок пока нет
- Legal Medicine: Chapter IV - Medicolegal Aspects of DeathДокумент27 страницLegal Medicine: Chapter IV - Medicolegal Aspects of DeathIchimaru Tokugawa100% (1)
- GoutДокумент32 страницыGoutEren YooОценок пока нет
- CH 20 Manter Gatz S Essentials of Clinical Neuroanatomy and NeurophysiologyДокумент27 страницCH 20 Manter Gatz S Essentials of Clinical Neuroanatomy and Neurophysiologyasarg214100% (1)