Вам также может понравиться
- Mitochondria and Its Pathways: Arvin G. Concha Mst-BioДокумент17 страницMitochondria and Its Pathways: Arvin G. Concha Mst-Biocassidy conchaОценок пока нет
- Think Yourself a Relationships Pro: THINK Yourself®От EverandThink Yourself a Relationships Pro: THINK Yourself®Оценок пока нет
- How Cells Harvest Chemical Energy - Cellular RespirationДокумент25 страницHow Cells Harvest Chemical Energy - Cellular RespirationMohamad Saadatian SipanОценок пока нет
- Cellular RespirationДокумент5 страницCellular RespirationCaptain MarvelОценок пока нет
- 13 Cellular Respiration-KEYДокумент7 страниц13 Cellular Respiration-KEYZOE STONEОценок пока нет
- 2 1 3 A Feedback Student Response Sheet (Revised 10 2 14)Документ7 страниц2 1 3 A Feedback Student Response Sheet (Revised 10 2 14)api-28182463470% (10)
- Ap - Cellular Energetics - Cell RespirationДокумент22 страницыAp - Cellular Energetics - Cell Respirationapi-251015600100% (1)
- Introduction To MetabolismДокумент27 страницIntroduction To Metabolismcream oОценок пока нет
- Krebs Cycle: TCA/ Citric Acid CycleДокумент24 страницыKrebs Cycle: TCA/ Citric Acid Cyclejahanzeb aliОценок пока нет
- 13 Cellular Respiration-KEYДокумент8 страниц13 Cellular Respiration-KEYfkjnf100% (1)
- Cell Respiration WebQuest 2015Документ3 страницыCell Respiration WebQuest 2015Haley HaunОценок пока нет
- Metabolism NotesДокумент208 страницMetabolism Notesbairdman89Оценок пока нет
- AP Biology 018 - Positive and Negative Feedback Loops Video Review SheetДокумент2 страницыAP Biology 018 - Positive and Negative Feedback Loops Video Review SheetKaelyn BrushettОценок пока нет
- Cellular RespirationДокумент3 страницыCellular RespirationErnest Nuñez LoquinarioОценок пока нет
- Chapter Quiz General Biology 2Документ7 страницChapter Quiz General Biology 2Nikki San GabrielОценок пока нет
- ADP, ATP and Cellular RespirationДокумент30 страницADP, ATP and Cellular RespirationJillian Lao100% (2)
- Study Guide 1Документ3 страницыStudy Guide 1msmickey24Оценок пока нет
- Study Questions Biology 141 Cellular Respiration, Kreb CycleДокумент3 страницыStudy Questions Biology 141 Cellular Respiration, Kreb CycleMeï QadriОценок пока нет
- Chapter 5 PDFДокумент13 страницChapter 5 PDFNicholas LeongОценок пока нет
- Writing Lewis StructuresДокумент10 страницWriting Lewis StructuresnОценок пока нет
- AP Bio Chapter 9 Study GuideДокумент4 страницыAP Bio Chapter 9 Study GuidelilyhcyОценок пока нет
- A Quick Look at Biochemistry Carbohydrate Metabolism (ELENA1)Документ15 страницA Quick Look at Biochemistry Carbohydrate Metabolism (ELENA1)Elena FloresОценок пока нет
- Chapter 3. Muscular, Skeletal, and Nervous SystemДокумент53 страницыChapter 3. Muscular, Skeletal, and Nervous SystemFatiha AmiraОценок пока нет
- Cellular Respiration - ReviewerДокумент3 страницыCellular Respiration - ReviewerJoyce Cañal100% (1)
- Sodium Potassium Pump and Action PotentialДокумент4 страницыSodium Potassium Pump and Action PotentialPiyush BhallaОценок пока нет
- Aug 9 Sbi4u Quiz 4Документ3 страницыAug 9 Sbi4u Quiz 4api-216300751Оценок пока нет
- Reduction Potential - WikipediaДокумент28 страницReduction Potential - WikipediaBabar AliОценок пока нет
- Carbohydrate MetabolismДокумент43 страницыCarbohydrate MetabolismMarliza CauntaoОценок пока нет
- Skeletal Muscle and The Molecular Basis of Contraction-TextДокумент23 страницыSkeletal Muscle and The Molecular Basis of Contraction-TextSuraj BisoiОценок пока нет
- Lecture Quiz On Muscular System: QuestionsДокумент2 страницыLecture Quiz On Muscular System: QuestionsCeline MillerОценок пока нет
- Pericyclic ReactionsДокумент53 страницыPericyclic ReactionsSF ShagorОценок пока нет
- Lipid MetabolismДокумент33 страницыLipid MetabolismDharmveer SharmaОценок пока нет
- Chapter 2 BLMs AnswersДокумент3 страницыChapter 2 BLMs AnswershelloblargОценок пока нет
- Nutrition Support CalculationsДокумент5 страницNutrition Support Calculationsapi-333590706Оценок пока нет
- Kreb CycleДокумент6 страницKreb Cycle211BT014 Jeev Sheen JosephОценок пока нет
- Energy Work Power WebquestДокумент4 страницыEnergy Work Power Webquestapi-3299787630% (2)
- High School Science Virtual Learning: Biology Homeostasi SДокумент20 страницHigh School Science Virtual Learning: Biology Homeostasi SEdchel EspeñaОценок пока нет
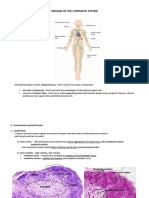
- Lymphatic SystemLabДокумент7 страницLymphatic SystemLabMax DelvalleОценок пока нет
- OAT Exam Secrets Test Prep Team - OAT Practice Questions - OAT Practice Tests & Exam Review For The Optometry Admission Test-Mometrix Media LLC (2013)Документ179 страницOAT Exam Secrets Test Prep Team - OAT Practice Questions - OAT Practice Tests & Exam Review For The Optometry Admission Test-Mometrix Media LLC (2013)SriОценок пока нет
- The Fate of Metabolism Andmabolic PathwaysДокумент25 страницThe Fate of Metabolism Andmabolic PathwaysXuân Vi100% (1)
- Enzyme: Jahangirnagar UniversityДокумент34 страницыEnzyme: Jahangirnagar UniversityShanian AhmedОценок пока нет
- TrishaДокумент3 страницыTrishaapi-343768938100% (3)
- 9 Dna Replication Scramble KeyДокумент2 страницы9 Dna Replication Scramble KeyTy Neal25% (4)
- U4 PhotosynthesisДокумент14 страницU4 PhotosynthesisjanaОценок пока нет
- ATP Photosynthesis and Cell Respiration Web QuestДокумент3 страницыATP Photosynthesis and Cell Respiration Web QuestJakie UbinaОценок пока нет
- Cellular Respiration PDFДокумент10 страницCellular Respiration PDFAnonymous HXLczq3Оценок пока нет
- Cell Respiration Worksheet KEYДокумент3 страницыCell Respiration Worksheet KEYerikabeltranОценок пока нет
- Aerobic Anaerobic RespirationДокумент15 страницAerobic Anaerobic RespirationTing Ming ChanОценок пока нет
- What Are CarbohydratesДокумент36 страницWhat Are CarbohydratesPINNACLE CAFEОценок пока нет
- MYP Exercises. Cell Respiration.: AnswerДокумент4 страницыMYP Exercises. Cell Respiration.: AnswerRishar bokОценок пока нет
- Chapter One, Structure and Bonding in Organic MoleculesДокумент4 страницыChapter One, Structure and Bonding in Organic MoleculesAmin JamjahОценок пока нет
- Atp Synthesis by Cellular Respiration AT P: Caroline SueperДокумент9 страницAtp Synthesis by Cellular Respiration AT P: Caroline SueperCaroline SueperОценок пока нет
- Beta-Oxidation of Fatty AcidsДокумент6 страницBeta-Oxidation of Fatty AcidsBinku Dutta100% (1)
- ProteinsДокумент11 страницProteinsCed HernandezОценок пока нет
- CH 7 Notes Cellular RespirationДокумент3 страницыCH 7 Notes Cellular RespirationCJ100% (1)
- Muscular System ReviewerДокумент16 страницMuscular System ReviewerNicole ZaputОценок пока нет
- Reviewer ANPH111 CandyДокумент21 страницаReviewer ANPH111 CandyRein TopacioОценок пока нет
- HIS 6 - Haemoglobin Recall Oxygen Binding Properties of HB: Iron (II) - Protoporphyrin IXДокумент2 страницыHIS 6 - Haemoglobin Recall Oxygen Binding Properties of HB: Iron (II) - Protoporphyrin IXKristin DouglasОценок пока нет
- Energy TransformationДокумент4 страницыEnergy TransformationAsh Nico BanquilloОценок пока нет
- Treatment of Bronchospasm During Anesthesia .28Документ4 страницыTreatment of Bronchospasm During Anesthesia .28Anish H DaveОценок пока нет
- Asa Difficult Airway AlgorithmДокумент20 страницAsa Difficult Airway AlgorithmspidymahiОценок пока нет
- Trach Brochure TeachingДокумент2 страницыTrach Brochure Teachingapi-246639896100% (1)
- Introduction To Mechanical VentilationДокумент5 страницIntroduction To Mechanical VentilationHussam GujjarОценок пока нет
- CHest Physio For NeonatesДокумент7 страницCHest Physio For Neonateschri23Оценок пока нет
- Human PerformanceДокумент8 страницHuman PerformanceRamОценок пока нет
- Extubation After Difficult IntubationДокумент3 страницыExtubation After Difficult Intubationramanrajesh83Оценок пока нет
- MT Vitals Signs - RespirationДокумент11 страницMT Vitals Signs - RespirationpriyankaОценок пока нет
- AtelactasisДокумент89 страницAtelactasiswahyuОценок пока нет
- Campbell Biology: Concepts & Connections,: Lecture by Edward J. ZaliskoДокумент90 страницCampbell Biology: Concepts & Connections,: Lecture by Edward J. ZaliskoAnnОценок пока нет
- Bioinstrument 7 (Respiratory System) 16-22Документ7 страницBioinstrument 7 (Respiratory System) 16-22vividmeОценок пока нет
- Investigatory Project Biology Class 11Документ32 страницыInvestigatory Project Biology Class 11ashmitav8m10402Оценок пока нет
- PulmoVista 500 - APPLICATION - Identify Onset of Derecruitment - 032018Документ2 страницыPulmoVista 500 - APPLICATION - Identify Onset of Derecruitment - 032018tomОценок пока нет
- Detailed Lesson Plan Template. 19 SeptemberДокумент3 страницыDetailed Lesson Plan Template. 19 SeptemberrutendoОценок пока нет
- CopdДокумент60 страницCopdRizqy Shofianingrum100% (1)
- Respiratory EmbryologyДокумент7 страницRespiratory EmbryologySam JamОценок пока нет
- CombitubeДокумент38 страницCombitubebrentupdegraffОценок пока нет
- Nursing Care PlanДокумент5 страницNursing Care PlanPaola Marie VenusОценок пока нет
- Ge Carescape R860Документ95 страницGe Carescape R860TRan DuongОценок пока нет
- Altitude Training and Sea Level PerformanceДокумент4 страницыAltitude Training and Sea Level PerformanceALEX SNEHA100% (1)
- Delhi Public School Dwarka Holiday Homework Class: X BiologyДокумент1 страницаDelhi Public School Dwarka Holiday Homework Class: X BiologyAdityaShailajОценок пока нет
- Pink Puffer Blue BloaterДокумент10 страницPink Puffer Blue BloaterMelinda MarianniОценок пока нет
- Chapter 19 PULMONARY FUNCTION TESTINGДокумент13 страницChapter 19 PULMONARY FUNCTION TESTINGZahra Margrette Schuck0% (1)
- Introduction To Respiratory SystemДокумент4 страницыIntroduction To Respiratory SystemExamville.com100% (3)
- IOS - Interpretation and Practical Applications - CHESTДокумент7 страницIOS - Interpretation and Practical Applications - CHESTthiago gibsonОценок пока нет
- Jurnal ARDSДокумент8 страницJurnal ARDSrifki yulinandaОценок пока нет
- Cell Energy WorksheetДокумент3 страницыCell Energy WorksheetMichael BarriosОценок пока нет
- Biots BreathingДокумент2 страницыBiots Breathingramandeep_ahluwaliaОценок пока нет
- Mechanical Ventilation: BasicsДокумент28 страницMechanical Ventilation: BasicsJohn WONG75% (4)
- Class XI Breathing and Exchange of Gases Biology (MCQ'S) (21 - 07 - 2023)Документ5 страницClass XI Breathing and Exchange of Gases Biology (MCQ'S) (21 - 07 - 2023)soda pdfОценок пока нет
- Periodic Tales: A Cultural History of the Elements, from Arsenic to ZincОт EverandPeriodic Tales: A Cultural History of the Elements, from Arsenic to ZincРейтинг: 3.5 из 5 звезд3.5/5 (137)
- Monkeys, Myths, and Molecules: Separating Fact from Fiction, and the Science of Everyday LifeОт EverandMonkeys, Myths, and Molecules: Separating Fact from Fiction, and the Science of Everyday LifeРейтинг: 4 из 5 звезд4/5 (1)
- Is That a Fact?: Frauds, Quacks, and the Real Science of Everyday LifeОт EverandIs That a Fact?: Frauds, Quacks, and the Real Science of Everyday LifeРейтинг: 5 из 5 звезд5/5 (4)
- The Disappearing Spoon: And Other True Tales of Madness, Love, and the History of the World from the Periodic Table of the ElementsОт EverandThe Disappearing Spoon: And Other True Tales of Madness, Love, and the History of the World from the Periodic Table of the ElementsРейтинг: 4 из 5 звезд4/5 (146)
- The Regenerative Grower's Guide to Garden Amendments: Using Locally Sourced Materials to Make Mineral and Biological Extracts and FermentsОт EverandThe Regenerative Grower's Guide to Garden Amendments: Using Locally Sourced Materials to Make Mineral and Biological Extracts and FermentsРейтинг: 5 из 5 звезд5/5 (3)
- Chemistry for Breakfast: The Amazing Science of Everyday LifeОт EverandChemistry for Breakfast: The Amazing Science of Everyday LifeРейтинг: 4.5 из 5 звезд4.5/5 (90)
- Organic Chemistry for Schools: Advanced Level and Senior High SchoolОт EverandOrganic Chemistry for Schools: Advanced Level and Senior High SchoolОценок пока нет
- The Elements We Live By: How Iron Helps Us Breathe, Potassium Lets Us See, and Other Surprising Superpowers of the Periodic TableОт EverandThe Elements We Live By: How Iron Helps Us Breathe, Potassium Lets Us See, and Other Surprising Superpowers of the Periodic TableРейтинг: 3.5 из 5 звезд3.5/5 (22)
- Meltdown: Nuclear disaster and the human cost of going criticalОт EverandMeltdown: Nuclear disaster and the human cost of going criticalРейтинг: 5 из 5 звезд5/5 (5)
- Essential Oil Chemistry Formulating Essential Oil Blends that Heal - Aldehyde - Ketone - Lactone: Healing with Essential OilОт EverandEssential Oil Chemistry Formulating Essential Oil Blends that Heal - Aldehyde - Ketone - Lactone: Healing with Essential OilРейтинг: 5 из 5 звезд5/5 (1)
- Chemistry: a QuickStudy Laminated Reference GuideОт EverandChemistry: a QuickStudy Laminated Reference GuideРейтинг: 5 из 5 звезд5/5 (1)
- The Periodic Table of Elements - Post-Transition Metals, Metalloids and Nonmetals | Children's Chemistry BookОт EverandThe Periodic Table of Elements - Post-Transition Metals, Metalloids and Nonmetals | Children's Chemistry BookОценок пока нет
- Handbook of Formulating Dermal Applications: A Definitive Practical GuideОт EverandHandbook of Formulating Dermal Applications: A Definitive Practical GuideОценок пока нет
- Taste: Surprising Stories and Science About Why Food Tastes GoodОт EverandTaste: Surprising Stories and Science About Why Food Tastes GoodРейтинг: 3 из 5 звезд3/5 (20)
- A Perfect Red: Empire, Espionage, and the Quest for the Color of DesireОт EverandA Perfect Red: Empire, Espionage, and the Quest for the Color of DesireРейтинг: 4 из 5 звезд4/5 (129)
- Formulating, Packaging, and Marketing of Natural Cosmetic ProductsОт EverandFormulating, Packaging, and Marketing of Natural Cosmetic ProductsОценок пока нет
- Tribology: Friction and Wear of Engineering MaterialsОт EverandTribology: Friction and Wear of Engineering MaterialsРейтинг: 5 из 5 звезд5/5 (1)
- Science Goes Viral: Captivating Accounts of Science in Everyday LifeОт EverandScience Goes Viral: Captivating Accounts of Science in Everyday LifeРейтинг: 5 из 5 звезд5/5 (1)
- The Periodic Table of Elements - Alkali Metals, Alkaline Earth Metals and Transition Metals | Children's Chemistry BookОт EverandThe Periodic Table of Elements - Alkali Metals, Alkaline Earth Metals and Transition Metals | Children's Chemistry BookОценок пока нет