Вам также может понравиться
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- Federal Public Service Commission Aga Khan RoadДокумент1 страницаFederal Public Service Commission Aga Khan RoadSHOAIBОценок пока нет
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (895)
- Modern DrmaДокумент7 страницModern DrmaSHOAIBОценок пока нет
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- Macroeconomic Indicators Improve: SBP's Quarterly ReportДокумент2 страницыMacroeconomic Indicators Improve: SBP's Quarterly Reportshoaib625Оценок пока нет
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (399)
- Labour Force P TurkeyДокумент2 страницыLabour Force P TurkeySHOAIBОценок пока нет
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (266)
- Labour Force, Total Rate of TurkeyДокумент2 страницыLabour Force, Total Rate of TurkeySHOAIBОценок пока нет
- Graphs SssДокумент6 страницGraphs SssSHOAIBОценок пока нет
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
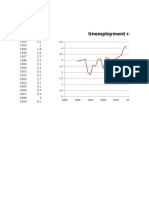
- Unemploy Ment Rate in UAEДокумент2 страницыUnemploy Ment Rate in UAESHOAIBОценок пока нет
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- Challan Form 2014Документ1 страницаChallan Form 2014Tanzeel Ur RahmanОценок пока нет
- US Involvement in YemenДокумент2 страницыUS Involvement in YemenSHOAIBОценок пока нет
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- Internship ReportДокумент15 страницInternship ReportSHOAIB0% (1)
- Procedural DemocracyДокумент1 страницаProcedural DemocracySHOAIBОценок пока нет
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- Unemployment Rate of PeruДокумент2 страницыUnemployment Rate of PeruSHOAIBОценок пока нет
- Pakistan KnowledgeДокумент3 страницыPakistan KnowledgeSHOAIBОценок пока нет
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2259)
- Calculation of Mean, Median, Mode, and Range Given DataДокумент2 страницыCalculation of Mean, Median, Mode, and Range Given DataSHOAIBОценок пока нет
- Mass Communication: Assignment TopicДокумент11 страницMass Communication: Assignment TopicSHOAIBОценок пока нет
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (344)
- Letter To Urdu DigestДокумент1 страницаLetter To Urdu DigestSHOAIBОценок пока нет
- OptimismДокумент2 страницыOptimismSHOAIBОценок пока нет
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- Introduction To Nature of Indian FederalismДокумент14 страницIntroduction To Nature of Indian FederalismSHOAIBОценок пока нет
- Apolitical or Depoliticised? Pakistan's Youth and PoliticsДокумент34 страницыApolitical or Depoliticised? Pakistan's Youth and PoliticsSHOAIBОценок пока нет
- GC, University Lahore: Topic: - Five of The Manners Declined From Our SocietyДокумент3 страницыGC, University Lahore: Topic: - Five of The Manners Declined From Our SocietySHOAIBОценок пока нет
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- AnДокумент5 страницAnSHOAIBОценок пока нет
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- New Microsoft Office Word DocumentДокумент3 страницыNew Microsoft Office Word DocumentSHOAIBОценок пока нет
- Structure of Cities 2Документ35 страницStructure of Cities 2SHOAIBОценок пока нет
- Istan S: Economy & Security IssuesДокумент14 страницIstan S: Economy & Security IssuesSHOAIBОценок пока нет
- Electrical Energy Crises Remedial Measures, Emphasizing The Hydel Power RoleДокумент44 страницыElectrical Energy Crises Remedial Measures, Emphasizing The Hydel Power RoleSHOAIBОценок пока нет
- Quaid-i-Azam and Early Problems of Pakistan: o o o o o o o oДокумент22 страницыQuaid-i-Azam and Early Problems of Pakistan: o o o o o o o oSHOAIBОценок пока нет
- Violence Against Children in Tanzania Report1Документ152 страницыViolence Against Children in Tanzania Report1api-67201372Оценок пока нет
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (121)
- Jack The Ripper-OutlineДокумент3 страницыJack The Ripper-Outlineapi-281742815Оценок пока нет
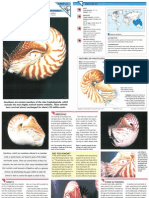
- Wildlife Fact File - Primitive Animals - Pgs. 31-40Документ20 страницWildlife Fact File - Primitive Animals - Pgs. 31-40ClearMind8450% (4)
- PORNOGRAPHY (Page 97) Pornography Addiction and Its Effects-It Used To Be Easy For Parents To Monitor The Kind ofДокумент2 страницыPORNOGRAPHY (Page 97) Pornography Addiction and Its Effects-It Used To Be Easy For Parents To Monitor The Kind ofGazelle CalmaОценок пока нет
- Rape As Social MurderДокумент4 страницыRape As Social MurdermanishsrjОценок пока нет
- Agrawal Janvi COM 100 TransgenderДокумент2 страницыAgrawal Janvi COM 100 TransgenderJanvi AgrawalОценок пока нет
- UntitledДокумент3 страницыUntitledapi-278804782Оценок пока нет
- Portrayal of Genders in The MediaДокумент4 страницыPortrayal of Genders in The MediaBellaCortezОценок пока нет
- Sexual Health Asylum Seekers and RefugeesДокумент188 страницSexual Health Asylum Seekers and RefugeesRuth WilsonОценок пока нет
- Rayon Payne and Folksalert Answer & Affirmative Defenses LawsuitДокумент11 страницRayon Payne and Folksalert Answer & Affirmative Defenses LawsuitFolksalertОценок пока нет
- Pag-Ibig (Love), Pagnanasa (Desire) :: Filipino Adolescents and Sexual RisksДокумент14 страницPag-Ibig (Love), Pagnanasa (Desire) :: Filipino Adolescents and Sexual RisksHannah dela MercedОценок пока нет
- RA 7610 DoctrinesДокумент2 страницыRA 7610 Doctrinesanna bee100% (2)
- Als-Shs Perdev Activity Module3Документ2 страницыAls-Shs Perdev Activity Module3Joa PingalОценок пока нет
- Importance of CondomДокумент1 страницаImportance of CondomTin AcidreОценок пока нет
- Meiosis Phases (2021 - 12 - 19 23 - 29 - 01 UTC)Документ14 страницMeiosis Phases (2021 - 12 - 19 23 - 29 - 01 UTC)Selenia BonoОценок пока нет
- UK Home Office: LincolnshireДокумент36 страницUK Home Office: LincolnshireUK_HomeOfficeОценок пока нет
- Butler AgainstProperObjects 1994Документ26 страницButler AgainstProperObjects 1994Pedro VieiraОценок пока нет
- Promoting Safer Sex Through PleasureДокумент6 страницPromoting Safer Sex Through PleasureRoel PlmrsОценок пока нет
- Name: Sheena Fe G. Vecina Year and Section: BSED-Social Studies 1Документ6 страницName: Sheena Fe G. Vecina Year and Section: BSED-Social Studies 1Sheena Fe Vecina100% (1)
- UK Home Office: West YorkshireДокумент9 страницUK Home Office: West YorkshireUK_HomeOfficeОценок пока нет
- Stephen Garton - Histories of SexualityДокумент247 страницStephen Garton - Histories of SexualityJanAscot100% (2)
- Times Square Red Times Square Blue PDFДокумент223 страницыTimes Square Red Times Square Blue PDFJujyfruits100% (4)
- Capstone PaperДокумент20 страницCapstone PaperburnettmorgenОценок пока нет
- Task Antropology of Law (Paper Bahasa Inggris Antropologi Hukum)Документ7 страницTask Antropology of Law (Paper Bahasa Inggris Antropologi Hukum)Ratri NugraheniОценок пока нет
- Cell Cycle - Meiosis NotesДокумент63 страницыCell Cycle - Meiosis NotesKathleen Claire MontefalcoОценок пока нет
- Porcelain Club - Letter To HarvardДокумент2 страницыPorcelain Club - Letter To HarvardEvanОценок пока нет
- DMD Full Walkthrough - Chapter 3: at The New Apartment With D (Morning Scene)Документ51 страницаDMD Full Walkthrough - Chapter 3: at The New Apartment With D (Morning Scene)Sherjeel MalikОценок пока нет
- Rat Dissection Student VersionДокумент11 страницRat Dissection Student VersionKong Hung Xi100% (1)
- @anabolic Steroids - Steroid Pharmacology PDFДокумент77 страниц@anabolic Steroids - Steroid Pharmacology PDFMiguelOliveiraОценок пока нет
- (2003) IRVINE, J. The Sociologist As Voyeur - Social Theory and Sexuality Research, 1910-1978Документ29 страниц(2003) IRVINE, J. The Sociologist As Voyeur - Social Theory and Sexuality Research, 1910-1978Lorena Caminhas100% (1)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisОт EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisРейтинг: 3.5 из 5 звезд3.5/5 (2)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceОт EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceРейтинг: 4.5 из 5 звезд4.5/5 (516)
- Tales from Both Sides of the Brain: A Life in NeuroscienceОт EverandTales from Both Sides of the Brain: A Life in NeuroscienceРейтинг: 3 из 5 звезд3/5 (18)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)От EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Рейтинг: 4 из 5 звезд4/5 (378)
- Gut: the new and revised Sunday Times bestsellerОт EverandGut: the new and revised Sunday Times bestsellerРейтинг: 4 из 5 звезд4/5 (392)
- Why We Die: The New Science of Aging and the Quest for ImmortalityОт EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityРейтинг: 4 из 5 звезд4/5 (3)